
Abstract
Data from 11 pop-up archival transmitting tags attached to opah (Lampris guttatus, F. Lampridae) in the central North Pacific between November 2003 and March 2005 were used to describe their vertical movement and habitat. In the subtropical gyre northwest of the Hawaiian Islands, opah generally inhabited a 50–400 m depth range and 8–22°C temperatures. They were frequently found in depths of 50–150 m at night and in greater depths (100–400 m) during the day, but were constantly moving vertically within this broad range. At night, excursions below 200 m were not uncommon and during the day they were very likely to spend some time at depths <175 m. Their vertical speeds were generally <25 cm s−1 but on one occasion an opah descended at a burst speed of 4 m s−1. Vertical habitat use by individual opah apparently varied with local oceanographic conditions, but over a 24-h period the average temperature experienced was always in the narrow range of 14.7 to 16.5°C.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Opah (Lampris guttatus) (Lampridae), also called moonfish, are mid-water pelagic fish with worldwide distribution. Opah are typically found well offshore in temperate and tropical waters of all the world’s oceans, including the Mediterranean and Caribbean Seas (Heemstra 1986). Opah reportedly inhabit waters from the surface to the lower epipelagic–mesopelagic in excess of 500 m (Nakano et al. 1997). They can reach 144 kg in weight (Jordan 1905).
In the Hawaii longline fishery, opah are generally caught on deep sets targeting bigeye tuna; and, while not a target species, they represent an important commercial component of the bycatch. In 2005, for example, 13,332 opah were caught.Footnote 1
In this paper, pop-up archival transmitting (PAT) tags are used to describe the temperature and depth habitat and vertical movement of this species in the central North Pacific.
Methods
Study area
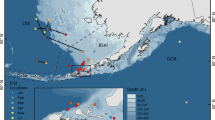
We deployed 16 Wildlife Computer (Redmond, WA, USA) PAT tags (versions 3 and 4) and 1 Microwave Telemetry (Columbia, MD, USA) high frequency PAT-100 tag on opah caught and released from US commercial longline vessels in the central North Pacific. These vessels typically set about 2,000 hooks on a monofilament longline that spans a distance of about 40 km. The tags were deployed in four trips covering spring, summer, and fall periods, over the period 10/11/2002 to 29/3/2005, and the spatial range from about 21 to 31°N latitude and 147 to 154°W longitude (Fig. 1; Table 1).

Lampris guttatus. Map with tag numbers and deployment and pop-up locations indicated as the base and head of each arrow respectively. Dotted, solid, and dashed arrows represent tags deployed on Trips 2, 3, and 4, while large solid arrow denotes the Microwave Telemetry Tag. Islands shown are the main Hawaiian Islands
Tagging
All tags were attached by co-author Don Hawn. The hardware tethered to each tag consisted of a single strand of 1.8-mm diameter fluorocarbon line, two stainless-steel sleeves, a single mechanical guillotine for the Wildlife Computer tags (Wildlife Computers RD 1500/1800) (to prevent tag implosion if the tag traveled deeper than 1,500 m), and a 3.5 × 1.7 cm modified titanium dart head. The length of fluorocarbon line used as a tether was 12.7 cm. Opah selected for tagging were usually caught during the first 2 h of the gear-hauling phase. Opah were selected at the beginning of the gear-hauling phase to reduce the physical exertion and physiological stress (i.e., hooking trauma and anthropogenic noises) loads that accrued during the soak period. Individual opah selected for this study were based on size (>30.0 kg). As they were brought along side of the vessel the general state of their body condition was assessed. Visual assessment for body damage included liveliness, body color, line damage to head and body, hooking location, eye and fin movement, and the presence of blood. Once an opah was selected for tagging, its branch line was released from the longline and immediately transferred to a 40-m braided tarred line. The fish was then brought to the side of the vessel and harpooned with a stainless steel applicator head with the tag attached. The tag was anchored at a depth of 10–12 cm anterior to the base of the dorsal fin. The opah was subsequently released by severing the leader closest to the hooking location. All opah tagged had estimated weights between 30 and 50 kg.
The Wildlife Computer tags were programmed to collect time-at-temperature and time-at-depth in 12 temperature and depth bins. The temperature bins were in increments of 2°C between the top and bottom bins, >26°C and <6°C, respectively. The depth bins were in 50-m increments from surface to 300 m and 100 m increments to 600 m, followed by bins of 600–750 m, 750–1,000 m, and >1,000 m. Two tags (# 30568 and 30571 from Trip 2) were programmed to compile the depth and temperature data in two 12-h time intervals representing day and night periods (in local time: 6:00–18:00 and 18:00–6:00). For all the other tags, the depth and temperature binned data were compiled in six 4-h time intervals (in local time: 02:00–06:00, 06:00–10:00, 10:00–14:00, 14:00–18:00, 18:00–22:00, and 22:00–02:00). Minimum and maximum depth and temperature plus six pairs of temperature and depth readings distributed between the extremes were also recorded for each 4 or 12-h time interval.
The Wildlife Computer tags were programmed to pop-up after a fixed time period, set at 6 months for most of the tags, and to transmit these binned data after pop-up via the Argos satellite.
The single Microwave Telemetry tag used was a high temporal resolution tag programmed to obtain temperature and depth data every 40 s over 28 days and transmit as much of these data as possible via the Argos satellite. The format of the data returned from the Wildlife Computer and Microwave Telemetry tags are so different that they will be analyzed and reported separately.
Satellite data
Satellite altimetry data were used to identify an eddy encountered by an opah. These data came from the Jason-1 altimeter, provided by AVISO, and had weekly temporal and 0.3° × 0.3° spatial resolution. Geostrophic currents were calculated from the gradient in sea surface height (Polovina et al. 1999).
Data analysis
Correspondence analysis was used to group the tags into clusters with similar time-at-temperature distributions. Correspondence analysis is a form of factor analysis used with contingency table data to describe the multi-dimensional data with a reduced number of dimensions (axes) (Grower 1987). Time-at-temperature frequency distributions for the clusters were computed as means of the individual tag frequency distributions. Mean daily depth and mean daily temperature were estimated from the 24-h time-at-depth and time-at-temperature distributions as the sum of the product of the bin frequency multiplied by the bin interval mid-point. For several tags the vertical temperature structure encountered by the fish was estimated from eight discrete temperature and depth values collected during each time bin. The tag reports minimum and maximum temperature at each discrete depth and we used the mean of these two temperatures in the vertical temperature structure calculation.
For the Microwave Telemetry tag the temperature and depth data collected over the tag deployment period were pooled into 24 hourly bins. Mean and standard deviation temperature and depth were computed for each of the 24 hourly bins.
Results
Tag returns
We received some data from 15 of the 16 deployed Wildlife Computer tags. Two tags transmitted pop-up locations but no subsequent data. Another three tags detached from the fish within 2 days of deployment when the fish reached depths in excess of 1,500 m. These rapid descents shortly after release suggested the opah suffered mortality. The very limited data obtained from these tags were not used in subsequent analyses.
For another three tags (#s 30568, 40610, 41863), the tag and possibly the opah were eaten by another animal some time after tagging. Evidence was based on an abrupt change in the tag data. During the first portion of the data time series, the depth data recorded minimum depths that were at least 50 m, but then abruptly that changed to indicate regular excursions to the surface during the remainder of the time series. At the same time the depth pattern changed, the temperature data stopped varying with depth and temperature remained constant a few degrees above surface temperature. Finally, concurrent with the changes in depth and temperature patterns, the light sensor stopped showing variation and remained constant at a low (dark) level. Three tags exhibited these conditions for 5, 13, and 24 days respectively, and we believe that during these periods they were inside an endothermic shark (Kerstetter et al. 2004). Subsequently, the tags were expelled from the predator, floated to the surface, and transmitted their data. The data from the predation portion of the time series were not included in our analyses; however, after removing those data, the three tags still contained 6–46 days of usable opah data.
Thus, of the 16 deployed Wildlife Computer tags, 10 tags provided at least 6 days of data that will be used in the subsequent analyses (Table 1).
Time-at-depth/time-at-temperature
We begin by presenting a composite or population picture of the opah habitat over this broad spatial and temporal range by pooling data from all 10 tags and constructing time-at-depth and time-at-temperature distributions.
Specifically, we examined the time-at-depth and time-at-temperature distributions from the eight Wildlife tags binned in 4-h time intervals for: (i) the day, represented by the 4-h period (10:00–14:00); (ii) the night, represented by the 4-h night period (22:00–02:00); and (iii) the daily, 24-h period constructed by pooling the data from all 10 tags over all time periods (Fig. 2). During the day, opah were generally distributed broadly between 100 and 400 m, while during the night they were shallower primarily, in the range 50–150 m (Fig. 2). During the day, opah often inhabited a broad range of 8–20°C water, while their night time temperature habitat was warmer, primarily 16–22°C, with a peak frequency in the 18–20°C bin (Fig. 2). A chi-square test found day and night frequency distributions both for depth and temperature were statistically different (P < 0.001). When the depth and temperature distributions are pooled over all time periods, it showed the opah inhabit a broad depth distribution. They spent about 40% of the time between 50 and 150 m and the remaining 60% of the time distributed between 150 and 400 m (Fig. 2). They very rarely spent time <50 m or >400 m (Fig. 2). The pooled time-at-temperature distribution showed a broad temperature distribution, fairly uniformly distributed from 8 to 22°C, with a slight peak between 18 and 20°C (Fig. 2). They rarely spent time in water <8°C or >22°C (Fig. 2).

Lampris guttatus. a Time-at-depth for day (10:00–14:00) and night (22:00–02:00). b Time-at-temperature for day (10:00–14:00) and night (22:00–02:00). c Time-at-depth and time-at-temperature pooled over all time periods. Bars indicate standard errors
Correspondence analyses
Next we examine the differences in the distributions between the tags. Mean daily time-at-depth and time-at-temperature distributions were constructed for each tag as the mean frequency over all the daily frequency distributions within each depth and temperature bin. A Chi-squared test was used to test whether the underlying time-at-depth and time-at-temperature distributions were the same for all tags. This hypothesis was rejected (P < 0.001). To explore this between tag variation we used time-at-temperature, rather than time-at-depth, since the temperature bins provide slightly finer resolution over the opah habitat range than the depth bins. Correspondence analysis was applied to the daily (24-h) time-at-temperature distributions and the first two factors explained 71% of the time-at-temperature variation between the 10 tags. With two exceptions, the 10 tags plotted on these first two factors showed a clustering by trip. All tags from Trip 2 clustered together (Fig. 3). All tags from Trip 3 clustered together along with Tag # 41863 from Trip 4 (Fig. 3). Tag # 41867 from Trip 4 formed its own cluster (Fig. 3). The remaining two tags from Trip 4 formed a cluster (Fig. 3). The time at-temperature distribution for the four clusters showed considerable variation (Fig. 4). Trips 2 and 4 both have bimodal distributions, especially Trip 4, where most the time was spent either in 18–22 or 8–12°C water (Fig. 4). There were four tags in trip 2 and two tags from trip 4 in that showed bimodal distributions. Trip 3 and Tag # 41867 have distributions that are more unimodal, especially Tag # 41867 which showed a very narrow temperature distribution with about 60% of the time spent at 12–16°C (Fig. 4). There were similar variations in the time-at-depth distributions (not shown) between the clusters.

Lampris guttatus. Plot of the time-at-temperature distributions for all 10 tags on the first two axes obtained from correspondence analysis

Time-at-temperature distributions for tag clusters determined from correspondence analyses
From the daily time-at-depth and time-at-temperature distributions for each cluster we can compute cluster mean daily (24-h) depth and temperature (Table 2). The mean daily depths ranged from 171 to 225 m while the mean daily temperatures ranged from 14.7 to 16.5°C with the means from three clusters between 16.0 and 16.5°C (Table 2). While the time-at-temperature distributions showed considerable variation between clusters, the mean daily temperatures were remarkably similar.
The clustering of the time-at-temperature by trip is not unexpected since the tags on each trip were deployed at about the same time and location while the trips were separated from each other in both location and date (Table 1). However, it is interesting that Tag # 41867, from Trip 4, did not cluster with the other tags from Trip 4. An examination of the sea surface height from satellite altimetry for August 17–23, 2003, showed evidence of a cold-core or cyclonic eddy located between the Tag # 41867 deploy and pop-up locations (Fig. 5). The geolocation estimation based on light levels recorded by the tag provided only very approximate daily position estimates with large confidence intervals so based on geolocation positions we cannot determine if the opah traveled through the center of the eddy or around the edge. However, the temperature–depth structure constructed from the opah vertical movement showed that during the August 18–26 period, the opah occupied a region with a broad temperature–depth relationship (Fig. 6). This temperature–depth relationship suggests the opah was in the convergent or downwelling edge of the eddy rather than the compact temperature–depth relationship that would be expected if the opah were in the divergent or upwelling center of the eddy (Fig. 6). The narrow time-at-temperature distribution (Fig. 4) is also consistent with the opah’s use of the edge of the cyclonic eddy since that is where the isotherms are more vertical and the opah would experience a narrower temperature range.

Sea surface height plot with geostrophic currents estimated from satellite altimetry 17/8/2003–23/8/2003, with a possible track line for opah (Lampris guttatus ) Tag # 41867 from deployment (solid box) to pop-off (arrow)

Contoured temperature–depth structure along the track of opah (Lampris guttatus) Tag # 41867 based on eight temperature and depth points collected during each 4-h time bin (black dots)
The other tag that did not cluster with tags in the same trip was Tag # 41863 from Trip 4 that clustered with Trip 3 tags. It is not clear why this occurred except that Tag # 41863 had only 6 days of data and was the southernmost tag of Trip 4, close in location to Trip 3 tags (Table 1; Fig. 1).
Mean daily depth and temperature
The similarity of mean daily temperature between clusters with different time-at-temperature distributions suggests the opah may be adjusting their vertical behavior so that the temperature encountered when averaged over each 24-h period falls within a narrow range. We can further investigate the temporal dynamics of mean daily depth and temperature in two opahs that are moving through an environment with a changing temperature–depth structure. This allows us to observe how these opah change mean depth in response to a changing temperature–depth distribution. The temperature–depth structure the opah encounters is estimated from a temporal interpolation using the eight pairs of temperature and depth values from each 4-h period over the duration of the tag deployment (Fig. 7) Overlaid on this vertical structure is the mean daily opah depth (Fig. 7). Tag # 41869 recorded an opah’s vertical movement and habitat from July 31 to December 10, 2003. The mean daily depth of this opah was about 200 m until about October 21 when it suddenly shifted to about 150 m for most of the remaining record (Fig. 7). The shift in mean depth occurred when there was a shoaling of the temperature–depth structure and it appeared the opah moved shallower in response to a shoaling of the temperature–depth structure (Fig. 7). The second Tag # 41867, is the opah that we believe used the edge of the cyclonic eddy. After about 5 days with a mean depth of about 125 m the opah appeared to have moved into a region with a deeper temperature–depth structure (the edge of the eddy) and its mean daily depth increased to 225 m (Fig. 7). About a week later, the temperature–depth structure shoaled somewhat and the opah moved shallower (Fig. 7). Thus, in both opah, their mean daily depths moved deeper or shallower coherent with changes in the temperature–depth structure (Fig. 7). However, when the mean daily depth and mean daily temperature for each of the two opahs were plotted together, the mean temperature exhibited a more constant long-term trend and did not alter in response to changes in the vertical structure as did mean depth (Fig. 8).

Lampris guttatus. Mean daily depth overlaid with the track contoured temperature–depth structure for opah # 41869 (top) and opah # 41867 (bottom)

Lampris guttatus. Mean daily depth (solid line) and mean daily temperature (dashed line) for opah Tag # 41869 (top) and opah Tag # 41867 (bottom)
Minimum and maximum temperature and depth
So far we have examined the opah time-at-depth and time-at-temperature distributions. We now examine the distribution of the minimum and maximum habitat values. The distribution of the shallowest and deepest depths and warmest and coolest temperatures occupied by the opah during each 24-h period is plotted in Fig. 9. The shallowest depth occupied each day was primarily 50–100 m while the deepest depth reached each day was often 350–450 m (Fig. 9). The very shallowest and deepest depths recorded were 28 and 736 m, respectively. The warmest daily temperature was generally 19–24°C while the coolest daily temperature was primarily 7–10°C (Fig. 9). The warmest and coolest temperatures recorded were 25.6 and 5.2°C, respectively. It is interesting to note that the shallowest depths fall largely in a narrow 50-m band while the deepest depths span a broad 150 m range (Fig. 9). Yet the shallowest temperatures covered a broader 6°C range while about 70% of the time the deepest temperatures were in a 2°C range (8–9°C) (Fig. 9). These results suggest that opah may define its habitat as water deeper than 50 m and warmer than 7°C (Fig. 9).

Lampris guttatus. Over a 24-h period, the minimum and maximum a depth and b temperature
The distribution of the deepest and shallowest depths for the mid-day, 4–h time bin (10:00–14:00) and the mid-night, 4-h time bin (22:00–02:00) provides further insight into the opah’s vertical dynamics. For the 10:00–14:00 day bin the deepest depths recorded were below 200 m as expected from the time-at-depth distribution (Fig. 10). However for this same period, the shallowest depths were primarily shallower than 175 m, with a peak between 100 and 125 m (Fig. 10). Thus, even during a daylight period, the opah generally made at least one excursion in this 4-h period into depths shallower than 175 m (Fig. 10). During the night bin, the shallowest depths were all shallower than 125 m as expected but a portion of the deepest depths extended from 125 m down to over 600 m (Fig. 10) Thus, at night while the opah were generally shallow they sometimes also made at least one excursion to deep depths.

Lampris guttatus. Minimum and maximum depth for a day (10:00–14:00) and b night (22:00–02:00)
The data from the Microwave Telemetry high frequency tag provides a further picture of the opah’s vertical dynamics. The tag recorded temperature and depth every 40 s; unfortunately, limitations in battery power allowed only a fraction of the data to be transmitted to the Argos satellite. Analysis of the vertical depth change in successive 40-s intervals showed that about 60% of the time the opah moved vertically less than 10 m in 40 s or 0.25 m/s, while about 40% of the time it moved vertically between 0.25 and 0.5 m/s. However, in one instance, it exhibited much faster movement. At 07:15 a.m. the opah descended from 43 to 204.4 m in 40 s then in the next 40-s interval to 258 m where it remained. The initial rapid descent of 161.4 m in 40 s or about 4 m/s may have been a predator-avoidance response.
Lastly, mean hourly depth, temperature, and their standard deviations are computed from the Microwave Telemetry high-frequency tag for all data pooled into 24, 1-h bins (Fig. 11). The mean depth and temperature showed a familiar diurnal pattern of shallower and warmer habitat during the night and deeper and cooler habitat during the day (Fig. 11). The large standard deviation of depth and temperature at 3:00 and 4:00 hours with means intermediate between day and night periods suggests that sometimes the opah stayed at shallow depths during these hours while at other times it remained at greater depths (Fig. 11).

Lampris guttatus. Mean and standard deviation (bars) of hourly temperature (top) and depth (bottom) from the Microwave Telemetry high-frequency Tag # 52494 from all data pooled over a 24-h period
Discussion and conclusions
In the central North Pacific, opah generally inhabited a 50–400 m depth range and a 8–22°C temperature range. They often exhibited vertical behavior like many other large pelagic visual predators, including swordfish and bigeye tuna, with deeper day and shallower night depth distributions (Musyl et al. 2003; Carey and Robinson 1981). At night, opah often occupied the 50–150 m depth range, while during the day they inhabited a broader range of 100–400 m. Further, opah frequently deviated from this diurnal pattern. During the day opah made at least one excursion into shallow depths and at night they often descended deep, especially during the hours of 3:00 and 4:00 hours. Unlike many of the tunas and billfishes that generally reside at or near the surface at night, opah rarely occupied depths <50 m. The deep habitat of opah from the time-at-depth and maximum daily depth show they frequented 275–475 m daily but generally did not spend much time at depths greater than 400 m. By comparison, bigeye tuna have a broader habitat range than opah. They reside at or very near the surface during the night and frequently descend and reside at depths between 400 and 500 m during the day to forage (Musyl et al. 2003). However, yellowfin tuna around Hawaii generally occupy depths <100 m during both day and night (Brill and Lutcavage 2001). Thus at least when foraging during the day opah appear to occupy a vertical niche deeper than yellowfin tuna and shallower than bigeye tuna.
Diets of yellowfin, opah, and bigeye also suggest there may be some partitioning of the forage base. While there are no published diet studies available for opah in the North Pacific, observations of stomachs of opah caught on Hawaii-based commercial longliners suggested the dominance of squids in their diet (D. Hawn personal communication). A study on the diet of southern opah (Lampris immaculatus) along the Patagonia Shelf showed they had a narrow range of prey items with the most common being the deepwater onychoteuthid squid (Moroteuthis ingens) (Jackson et al. 2000). By contrast squids are not a large component of bigeye and yellowfin tunas diets (Bertand et al. 2002). Bigeye tuna diets in the subtropical gyre include an abundance of deepwater myctophids and crustaceans while yellowfin tuna diets are dominated by shallow water fishes including juvenile reef fishes and epipelagic fishes (Bertand et al. 2002).
Considerable variation occurred in the opah’s use of its vertical habitat. Opah tagged close together in location and time generally had time-at-temperature distributions that were similar compared to those tagged farther apart in location and time. Further, when opah were tagged close in location and time but one used the edge of an eddy with a temperature–depth distribution different from that occupied by the other opah, their time-at-depth distributions also differed. This suggested the opah’s use of its vertical habitat can vary in response to changes in local oceanography.
It has been suggested that the absolute temperature does not define tuna and billfish habitat as well as the temperature range (Brill and Lutcavage 2001). The opah’s upper limit of their temperature habitat varied somewhat, 22–26°C in Trip 2 to 20–22°C in Trip 3. However, the lower limit of their temperature habitat was narrower at 8–9°C for all clusters. Also, the range between warmest and coolest temperatures they inhabit varied from about 18°C in Trip 2 to 10°C for Tag # 41867. Based on the minimum and maximum depth and temperature distributions, it appeared that the opah’s habitat consisted of water deeper than 50 m and warmer than 7°C.
However, within these bounds the opah’s vertical movement and habitat were likely determined by finding prey and avoiding predators, while occupying water temperature such that the mean daily temperature it occupied is limited to a narrow range from about 14.7 to 16.5°C. They appeared to vary their time-at-depth and time-at-temperature so that over the course of 24 h the average temperature they experienced remainded in a narrow range. This is true for both the same opah over several months moving through different habitats and different opah in habitats separated by 1,000 km and 6 months. The narrow range of mean daily temperature is in contrast to tuna that apparently have more ability to thermoregulate and hence can tolerate a wide variation in mean daily temperature. For example, giant bluefin tuna mean daily temperatures that were also estimated with PAT tags varied by as much as 10°C both for the same fish over time and among fishes (Lutcavage et al. 1999).
Regarding predation, the opah’s night depth limit of about 50 m may represent a strategy to reduce encounters with predators using the surface layer at night. The occurrence of potential predators in proximity to opah was suggested by both the high loss of PAT tags (3 out of 15) on opah that were likely ingested by a predator, possibly mako or great white shark, and the example of the opah’s rapid descent as recorded by the high-frequency tag.
Large pelagic animals are thought to forage in association with ocean features. Because of the difficulty in accurately estimating the daily positions of opah with only light-based geolocation, we were not able to examine possible associations between the movements of our tagged opah and ocean features in most cases. However, in one instance, the opah was tagged near an obvious eddy. The suggested use of the edge of a cyclonic eddy by the opah was consistent with the hypothesis that new production generated from water vertical upwelled at the center would be concentrated at the eddy’s edge to support a food web (Seki et al. 2001). Residence at the edge of a cold core eddy has been documented for another pelagic animal, the loggerhead turtle (Polovina et al. 2006).
Notes
Hawaii longline fishery logbook summary reports http://www.pifsc.noaa.gov/fmsd/reports.php (accessed February 1, 2007).
References
Bertand A, Bard FX, Jose E (2002) Tuna food habits related to the microneckton distribution in French Polynesia. Mar Bio 140:1023–1037
Brill RW, Lutcavage ME (2001) Understanding environmental influences on movements and depth distributions of tunas and billfishes can significantly improve population assessments. AFS Sym 25:179–198
Carey FG, Robinson BH (1981) Daily patterns in the activities of swordfish, Xiphias gladius, observed by acoustic telemetry. Fish Bull 79:277–292
Grower JC (1987) Introduction to ordination techniques. In: Legendre P, Legendre L (eds) Developments in numerical ecology. NATO ASI Ser G14:3–64. Kluwer, Dordrecht
Heemstra PC (1986) Family No. 117. Lampridae. In: Smith MM, Heemstra PC (eds) Smiths’ sea fishes. Springer, Berlin
Jackson GD, Buxton NG, George MJA (2000) Diet of the southern opah Lampris immaculatus on the Patagonian Shelf: the significance of the squid Moroteuthis ingens and anthropogenic plastic. Mar Ecol Prog Ser 206:261–271
Jordan DS (1905) A guide to the study of fishes, Vol 1. Holt and Co, New York
Kerstetter DW, Polovina JJ, Graves JE (2004) Evidence of shark predation and scavenging of fishes equipped with pop-up satellite archival tags. Fish Bull 102(4):750–756
Lutcavage ME, Brill RW, Somai GB, Chase BC, Howey PW (1999) Results of pop-up satellite tagging of spawning size class fish in the Gulf of Maine: do North Atlantic bluefin tuna spawn in the mid-Atlantic? Can J Fish Aquat Sci 56:173–177
Musyl MK, Brill RW, Boggs CH, Curran DS, Kazama TK, Seki MP (2003) Vertical movement of bigeye tuna (Thunnus obesus) associated with islands, buoys, and seamounts near the main Hawaiian Islands from archival tagging data. Fish Oceanogr 12(3): 152–169
Nakano H, Okazaki M, Okamoto H (1997) Analysis of catch depth by species for tuna longline fishery based on catch by branch lines. Bull Nat Res Inst Far Seas Fish 34:43–62
Polovina JJ, Kleiber P, Kobayashi D (1999) Application of TOPEX/POSEIDON satellite altimetry to simulate transport dynamics of larvae of the spiny lobster (Panulirus marginatus), in the Northwestern Hawaiian Islands, 1993–96. Fish Bull 97:132–143
Polovina JJ, Uchida I, Balazs G, Howell E, Parker D, Dutton P (2006) The Kuroshio extension current bifurcation region: a pelagic hotspot for juvenile loggerhead sea turtles. Deepsea Res II(53):326–339
Seki MP, Polovina JJ, Brainard RE, Bidigare RR, Leonard CL, Foley DG (2001) Biological enhancement at cyclonic eddies tracked with GOES thermal imagery in Hawaiian waters. GRL 28(8):1583–1586
Acknowledgments
We thank the captains and crews of the F/V Sea Pearl and F/V Kelly Ann used to tag the opah. The Matlab scripts used in the correspondence analyses were written and kindly provided by Prof. Jean-Philippe Labat and David Pellicer, Laboratoire d’Océanographie de Villefranche, University Pierre et Marie Curie, Paris, France. This work was partially funded by a grant from the University of Hawaii Pelagic Fisheries Research Program under Cooperative Agreement NA17RJ12301 from NOAA and a grant from the Ocean Exploration Program, NOAA.
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by J.P. Grassle.
Rights and permissions
About this article
Cite this article
Polovina, J.J., Hawn, D. & Abecassis, M. Vertical movement and habitat of opah (Lampris guttatus) in the central North Pacific recorded with pop-up archival tags. Mar Biol 153, 257–267 (2008). https://doi.org/10.1007/s00227-007-0801-2
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00227-007-0801-2