
Abstract
Feeding, egg production, hatching success and early naupliar development of Calanus finmarchicus were measured in three north Norwegian fjords during a spring bloom dominated by diatoms and the haptophyte Phaeocystis pouchetii. Majority of the copepod diet consisted of diatoms, mainly Thalassiosira spp. and Chaetoceros spp., with clearance rates up to 10 ml ind−1 h−1 for individual algae species/groups. Egg production rates were high, ranging from ca 40 up to 90 eggs f−1 d−1, with a hatching success of 70–85%, and fast naupliar development through the first non-feeding stages. There was no correlation between the egg or nauplii production and diatom abundance, but the hatching success was slightly negatively correlated with diatom biomass. However, the overall high reproductive rates suggested that the main food items were not harmful for C. finmarchicus reproduction in the area, although direct chemical measurements were not conducted. The high population egg production (>1,20,000 eggs m−2 d−1) indicated that a large part of the annual reproduction took place during the investigation, which stresses the importance of diatom-dominated spring phytoplankton bloom for population recruitment of C. finmarchicus in these northern ecosystems.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
At high latitudes, Calanus finmarchicus generally has one main spawning period per year (Tande 1982; Falkenhaug et al. 1997a, b), although a second less pronounced period of reproductive activity may occur during late summer (Arashkevich et al. 2004; Pasternak et al. 2004). In northern Norway, late copepodites of C. finmarchicus rise to surface waters in January–February, mature and start the egg production in March–early April and reach the peak in egg production during the spring bloom in April (Hopkins et al. 1984; Diel and Tande 1992). The peak abundance of nauplii stages is observed from April onwards, by June–July transforming to a peak abundance of lipid collecting late copepodite stages, which form the bulk of the overwintering generation (Hopkins et al. 1984). Although a few eggs may be produced by C. finmarchicus before the spring bloom (Niehoff et al. 1999), it is generally believed that food is essential for gonad maturation and egg production, and that the reproductive biology of C. finmarchicus is closely related to phytoplankton development (Diel and Tande 1992; Hirche et al. 1997; Niehoff et al. 1999). In areas with only one annual generation, successful spring reproduction and juvenile development may directly affect the breeding stock of the successive year (Diel and Tande 1992), although also advection of zooplankton can at times be important (Aksnes et al. 1989; Falkenhaug et al. 1997a, b).
The spring bloom in northern Norway mainly consists of diatoms and the prymnesiophyte Phaeocystis pouchetii (Wassmann et al. 2005): the food quality of both of these species/groups is somewhat controversial issue. Although some studies show a positive correlation between reproduction of Calanus spp. and diatom biomass (Niehoff et al. 1999; Irigoien et al. 2000a, b; Jónasdóttir et al. 2002), others have presented evidence on harmful effects of diatoms on egg production and hatching (Ban et al. 1997; Miralto et al. 1999) and juvenile development (Poulet et al. 1995, 2003; Ianora et al. 2004) of a number of copepod species, including C. finmarchicus (Ban et al. 1997; Starr et al. 1999). Whereas there is strong evidence of a reduced hatching success of copepod eggs produced on pure diatom diets from laboratory experiments (Ban et al. 1997), the field studies on copepod–diatom interactions are relatively scarce (Miralto et al. 2003; Halsband-Lenk et al. 2005; Pierson et al. 2005; Poulet et al. 2006), and the general relevance of harmful effects of diatoms in the field has been questioned (Irigoien et al. 2002). Similarly, the effect of Phaeocystis spp. on secondary production and feeding of copepods is still not fully understood: whereas some studies show high feeding rates and positive or no effect on egg production (Tande and Båmstedt 1987; Irigoien et al. 2000a, b), others reveal generally low feeding and egg production rates (Verity and Smayda 1989; Tang et al. 2001).
Most of the calanoid copepods are generally assumed to be omnivorous, and able to feed selectively (e.g. Cowles et al. 1988). Calanus finmarchicus has been shown to select diatoms (Meyer-Harms et al. 1999; Teegarden et al. 2001), dinoflagellates (Meyer-Harms et al. 1999; Levinsen et al. 2000) and ciliates (Ohman and Runge 1994; Nejstgaard et al. 1997; Irigoien et al. 1998; Levinsen et al. 2000), or to feed relatively non-selectively (Huntley 1981; Harris et al. 2000; Meyer et al. 2002; Koski and Wexels Riser 2006): e.g. Phaeocystis pouchetii seems to be ingested according to its availability (Meyer-Harms et al. 1999). However, only few field studies have combined measurements of selective feeding of Calanus spp. with reproduction and/or growth, so that the information on the effect of food selection on the actual production rates of this species remains controversial. Until now, the only studies combining selective feeding and egg production measurements of C. finmarchicus suggest high egg production during a period of selective feeding on diatoms (peak of the spring bloom), but low egg production during high post-bloom ingestion of P. pouchetii (Meyer-Harms et al. 1999; Niehoff et al. 1999), or controversially, lower egg production during high ingestion of diatoms, compared to higher egg production during high ingestion of the haptophyte Emiliania huxleyi (Nejstgaard et al. 1997). Similarly, diatoms and haptophytes (mainly P. pouchetii) seemed to have a positive effect for egg production of Calanus helgolandicus in English Channel off Plymouth (Irigoien et al. 2000b), but a diatom dominated diet induced poor egg production and/or hatching success of the same species in west coast of Norway (Nejstgaard et al. 2001b) as well as in English Channel off Roscoff (Poulet et al. 2006). For Calanus pacificus, ingestion of Thalassiosira spp. induced reduced naupliar survival, although egg production and mostly also hatching success remained unaffected (Leising et al. 2005; Pierson et al. 2005).
In the present study, I measured selective feeding, reproduction (egg production and hatching success) and early nauplii development of C. finmarchicus during the spring bloom in three north Norwegian fjords, in order to observe the magnitude and relevance of the possible negative effects of the dominant phytoplankton groups (diatoms and prymnesiophytes) on spring reproduction. The study was conducted during the peak reproductive season, when most of the females were supposedly spawning (Hopkins et al. 1984; Diel and Tande 1992), and no effect of gonad maturation on the egg production rates was therefore expected (see Campbell and Head 2000). Since most of the potential food during the study consisted of diatoms and Phaeocystis pouchetii (Wassmann et al. 2005), it was possible to investigate the diet selection during the dominance of these controversial species, and its consequence for reproduction and early nauplii development. Since C. finmarchicus is among the dominant copepod species in the area (Tande 1991; Falkenhaug et al. 1997a, b; Pasternak et al. 2000), with peak reproduction occurring during the spring bloom (Diel and Tande 1992; Arashkevich et al. 2004; Pasternak et al. 2004), any negative (or positive) effect of diatoms and/or P. pouchetii for its reproduction should have large consequences for these northern ecosystems.
Materials and methods
Study area
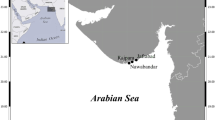
Grazing, egg production, hatching success and early naupliar development of C. finmarchicus were measured during a 10-day cruise in three north Norwegian fjords, Balsfjord, Ullsfjord and Malangen, in April 2001 (Fig. 1). Malangen (max. depth 450 m) and Ullsfjord (max. depth 300 m) are fjords which are relatively open towards the Norwegian coast, with deep sills and substantial water exchange, while Balsfjord (max. depth 190 m) is separated from the more open fjords by a shallow sill of 35 m (Wassmann et al. 1996). In April the water column in the area is, however, almost homogeneous, with temperatures around 2–4°C and salinities of 32.5–33.5 psu. The spring bloom generally starts in late March–early April, and reaches its peak towards the last part of April (Reigstadt et al. 2000; Wassmann et al. 2005). The spring bloom prevails for up to ca 1.5 months, and is typically dominated by diatoms and the haptophyte Phaeocystis pouchetii (Lutter et al. 1989). For details of the sampling area, see Wassmann et al. (1996, 2005) and Reigstadt et al. (2000).
In April 2001, the phytoplankton composition was dominated by diatoms and single cells of Phaeocystis pouchetii. A bloom of Chaetoceros sociales dominated the phytoplankton during the first sampling time in Balsfjord, whereas P. pouchetii was generally numerically dominant at other times and fjords sampled. The integrated suspended phytoplankton carbon (0–40 m) was 4,840, 6,790 and 6,420 mg C m−2 at Balsfjord, Malangen and Ullsfjord, respectively, with diatoms forming 44, 39 and 27%, respectively and P. pouchetii 28, 30 and 50% of the phytoplankton biomass (Wassmann et al. 2005). The maximum chlorophyll-a concentration varied between 0.5 and 3.2 μg l−1, and was, especially at Malangen and Ullsfjord, relatively equally distributed in the first 40 m of the water column (Fig. 1). At Balsfjord, Chl-a showed a more pronounced vertical peak in distribution, which during the cruise fluctuated between 8 and 20 m (P. Wassmann, University of Tromsø, unpublished data).

Map of the study area, showing the three investigated fjords, Malangen, Balsfjord and Ullsfjord, and the city of Tromsø in north Norway, and the vertical distribution of chlorophyll-a (μg l−1) in the 0–40 m water layer during the first sampling at each station (mean ± SE of triplicate samples). The arrow indicates the sampling depth (8 m)
Experiments
Each of the three fjords was visited three times with 3 days intervals. Egg production of C. finmarchicus was measured each time the fjord was visited, whereas hatching was measured during the first two visits, and grazing was measured during the first visit to each fjord. Development experiments were started from the eggs produced during the first visit to each fjord, and the water was exchanged every time the fjord was revisited (thus at 3 day intervals). In addition, seston was sampled from 8 m depth for phytoplankton and microzooplankton composition at the start of each egg production experiment.
Calanus finmarchicus for experiments were collected by vertical net hauls from the depth of ca 40 m to the surface, using a WP-2 plankton net (mesh size 180 μm), whereas incubation water was collected with Niskin bottles at 8 m depth and filtered <112 μm, to remove mesozooplankton grazers. Since the majority of C. finmarchicus females in March–April in the study area are dwelling in the uppermost 20 m (Falkenhaug et al. 1997a, b), where also the peak Chl-a concentration was detected (Fig. 1), sampling at 8 m depth was assumed to relatively well represent the food availability for this species in the area.
At each station, 3–7 females per each of 4 (grazing) or 5 (egg production) replicates were carefully sorted and placed either into bottles of 0.6 l (grazing) or 0.5 l egg production beakers (with a 500 μm mesh separating females from the produced eggs), containing <112 μm filtered in situ water. Three replicate bottles with GF/C filtered seawater were added as a control for background reproduction and three replicate bottles with <112 μm filtered in situ water without animals were included as a control for grazing experiments. The bottles were incubated close to in situ temperatures (ca 3°C) for ca 24 h; the grazing bottles were incubated on a plankton wheel (∼1 round per minute). A 100 ml sample from the start food suspension was preserved in 4% Lugol’s solution for later microscopic counts.
At the end of the incubations, eggs were counted, 100 ml of the grazing and control bottles were preserved in 4% Lugol, and the condition of females was checked (actively swimming vs. dead). The females in egg production controls (GF/C filtered seawater) were transferred to fresh filtered seawater; these control experiments were continued for three subsequent days. During the first visit to each fjord, the produced eggs were divided into four replicate bottles of 330 ml, containing <112 μm filtered in situ water, for development experiments. The hatching success of these eggs was estimated from the egg: nauplii ratio in the first development sample, taken after 3 days. During the second visit at the fjord, the eggs were left to hatch in GF/C filtered seawater in Petri-dishes for 3 days, after which the number of eggs and nauplii were counted (see Corkett et al. 1986 for hatching time of C. finmarchicus eggs). The hatching success was based on >120 eggs sample−1. Development bottles were sampled each time the fjord was visited: in Malangen and Balsfjord 100, 100 and 330 ml of the bottles were sampled respectively in days 3, 6 and 7/8, whereas in Ullsfjord 100 and 330 ml of the bottles were sampled at days 3 and 6, respectively. The sample was preserved in 4% Lugol for later counting of nauplii and determination of development stage (≥30 nauplii sample−1).
Analysis
The abundance of phytoplankton and microzooplankton at the start of each grazing experiment was counted from duplicate settled 2 ml or 25–50 ml samples, respectively. The whole samples were counted (≥50 cells per species) and ca 30 cells of each species at each fjord were sized. The cell volumes were calculated assuming simple geometric shapes and converted to carbon using volume-carbon conversions according to Montagnes et al. (1994). Clearance and ingestion rates were calculated according to Frost (1972), whereas selection index α of Chesson (1983) was used to evaluate the food selection. α was calculated as the ratio of the clearance rate on one food type to the sum of clearance rates on all food types. α varies between 0 and 1, and α > m−1, where m is the number of food types, indicates positive selection (Chesson 1978).
Due to the high concentration of females in incubation bottles, the concentration of the preferred food items, the diatoms Thalassiosira spp. and Chaetoceros spp., were strongly reduced during incubations (on average 54 and 58%, respectively), whereas the reduction of other food species was always <20%. This likely resulted in underestimation of clearance, ingestion and selection of these diatoms, but should not have affected the feeding rates on other species, or changed the general conclusions based on the food selection and diet composition (see Results and Discussion). However, the feeding rates and selection of the two preferred diatom species should be considered as minimum estimates.
Weight-specific ingestion and egg production were estimated based on the average female length of 2.8 mm (C. Wexels Riser, University of Tromsø, unpubl. data), length—carbon regression of Davis (1984), and egg carbon content of 0.29 μg C egg−1 (E. Arashkevich, Russian Academy of Science, unpublished data). Nauplii development was expressed as the increase of the mean stage (calculated from the stage frequency distribution in the samples) over the course of time. Nauplii mortality was calculated from the linear model of mortality (ln no of individuals as a function of time), after correcting for sampling mortality according to Klein Breteler et al. (2004). The production of nauplii (NI) was calculated by multiplying the egg production with the proportion of hatched eggs, separately for each fjord and sampling time. The production of nauplii in the first feeding stage (NIII) was calculated from the production of NI and specific daily mortality during 6 days; on day 6 ≥50% of the population had reached the third nauplius stage (see Results). The population egg production was calculated by multiplying the egg production rate with the number of females (P. Wassmann, University of Tromsø, unpublished data), separately for each fjord and sampling time.
Egg production, hatching success and nauplii production were tested for differences between the fjords and days (time the fjord was visited) with a two-way analysis of variance (ANOVA), whereas total ingestion and final development stage reached during the incubations were tested for differences between the fjords with a one-way ANOVA. All data were tested for normal distribution and homogenety of variances. Tukey HSD a posteriori test was used for pairwise comparisons. Clearance and ingestion rates as well as selection index α for or against each food species were formally tested against zero (significant clearance and ingestion rates) or the value of α indicating no selection (significant positive or negative selection), with a one-sample t-test. To test whether reproduction related to the composition or quantity of food, a Spearman rank correlation analysis was run for egg production, hatching, production of NI and NIII and quantity and proportional abundance of different algae and microzooplankton groups (diatoms, dinoflagellates, flagellates dominated by Phaeocystis pouchetii and ciliates) at the start of the incubations.
Results
Food concentration and composition
Food concentration varied from 27 to 223 μg C l−1, but was above or close to 100 μg C l−1 during most of the experiments, with diatoms dominating the biomass. The proportional abundance of diatoms was 61–84% in Malangen, 52–63% in Balsfjord and 42–49% in Ullsfjord, with flagellates (mainly Phaeocystis pouchetii) contributing 13–30, 6–18 and 34–40% and ciliates 2–6, 3–10 and 5–10% of the biomass in Malangen, Balsfjord and Ullsfjord, respectively. Dinoflagellates were least abundant in Malangen (1–5%), but increased in importance in Balsfjord (15–27%) and Ullsfjord (7–13%; Table 1). The maximum chl-a (>3 μg l−1) and diatom carbon (>200 μg C l−1) concentrations were representative for the average spring bloom concentrations in the area (Wassmann et al. 2005), and in the range of spring bloom concentrations observed in other temperate, boreal and arctic areas e.g. Dabob Bay (Washington State; Horner et al. 2005), central Gulf of Maine (Durbin et al. 2003), southern North Sea (Rousseau et al. 2002), English Channel (Pond et al. 1996) and Greenland Sea (Levinsen et al. 2000).
Ingestion and food selection
Calanus finmarchicus females were weakly selecting for diatoms, with clearance rates up to 10 ml ind−1 h−1 for individual algae species (Figs. 2, 3, 4). Copepods were significantly filtering different diatoms and ciliates, although in Balsfjord also flagellates, mostly consisting of Phaeocystis pouchetii, were filtered (Figs. 2, 3, 4a). The only species which were sometimes significantly selected for were the diatoms Thalassiosira spp. and Chaetoceros spp., whereas flagellates (P. pouchetii), dinoflagellates and the diatom Nitzchia sp. were occasionally selected against (Figs. 2, 3, 4c). The copepod diet was dominated by diatoms; especially in Malangen and Balsfjord where diatoms formed 96 and 82%, respectively, of the diet (Figs. 2, 3b). In Ullsfjord the diet was more diverse, with substantial contributions from diatoms (59%), flagellates (22%), dinoflagellates (9%) and ciliates (9%; Fig. 4b). The total ingestion of 22 μg C ind−1 d−1 in Malangen was significantly higher than the total ingestion of 7–8 μg C ind−1 d−1 in Balsfjord and Ullsfjord (one-way ANOVA; F2 = 13.5, Tukey HSD; P < 0.05), probably due to the significantly higher food concentration of ca 220 μg C l−1 in Malangen, compared to 94–96 μg C l−1 in Balsfjord/Ullsfjord (one-way ANOVA F2 = 38, Tukey HSD; P < 0.001; Table 1). The weight-specific ingestion was, however, relatively low at all fjords, 0.17, 0.06 and 0.06 μg C (μg C)−1 d−1 at Malangen, Balsfjord and Ullsfjord, respectively.

a Clearance rate (ml ind−1 h−1), b ingestion rate (μg C ind−1 d−1) and c selection (selection index α) of Calanus finmarchicus females in Malangen in northern Norway (mean ± SE). Fl Flagellate spp. (mainly Phaeocystis pouchetii), Dif diverse dinoflagellates, Th the diatoms Thalassiosira spp., Ch Chaetoceros spp. and Ni Nitzchia spp., Dia other diatoms, Ci ciliates (mainly Strombidium-type with an equivalent spherical diameter of ca 30 μm). The star indicates clearance and ingestion rates which are significantly different from zero (one sample t-test, P < 0.05), and selection which is significantly different from α of 0.16 (no selection; see Methods). The total ingestion rate and percentage contribution of different food groups to the diet are indicated in the figure, as is the line showing where food species are eaten according to their availability (no selection; α = 0.16)

a Clearance rate (ml ind−1 h−1), b ingestion rate (μg C ind−1 d−1) and c selection (selection index α) of Calanus finmarchicus females in Balsfjord in northern Norway (mean ± SE). At α value of 0.17 the food species is eaten according to its availability (no selection). Abbreviations as in Fig. 2

a Clearance rate (ml ind−1 h−1), b ingestion rate (μg C ind−1 d−1) and c selection (selection index α) of Calanus finmarchicus females in Ullsfjord in northern Norway (mean ± SE). At α value of 0.16 the food species is eaten according to its availability (no selection). Abbreviations as in Fig. 2
Reproduction and nauplii development
There were no significant differences in egg production between the three fjords or the three sampling times (two-way ANOVA; P > 0.05). The egg production rates in the three fjords were generally high, 60–90 eggs f−1 d−1, which corresponded to a weight-specific egg production of 0.13–0.20 μg C (μg C)−1 d−1, with a notable exception of the first visit in Malangen when egg production of ca 40 eggs f−1 d−1 [0.09 μg C (μg C)−1 d−1] was measured. In the absence of food, C. finmarchicus females never produced more than 8 eggs f−1 d−1, corresponding to a weight-specific egg production of 0.02 μg C (μg C)−1 d−1 (Fig. 5a).

a Egg production rate (eggs f−1 d−1), b hatching success (%), c early nauplii development (increase in mean stage as a function of time) and d nauplii mortality (ln no. of individuals as a function of time) of Calanus finmarchicus in Malangen, Balsfjord and Ullsfjord in northern Norway and in GF/C filtered seawater (mean ± SE). Closed circles Malangen, closed squares Balsfjord, stars Ullsfjord and open diamonds GF/C filtered seawater. The parameters for linear model of mortality, including the slope showing specific mortality of 10 ± 3% d−1, are shown in the figure. The egg production and hatching success in GF/C filtered seawater represent rates measured during a three-day incubation, whereas the rates measured in different fjords represent in situ rates during three subsequent visits to each fjord (see Material and Methods )
Hatching success was relatively high, ≥70%, and did not differ significantly between the three fjords (two-way ANOVA; P > 0.05). Hatching success of eggs produced in filtered seawater was always lower than hatching success of eggs produced with in situ water (41 ± 10%), and decreased with the time spent without food (Fig. 5b). The production of nauplii (NI) was significantly lower in Malangen than in Balsfjord or Ullsfjord (two-way ANOVA, F2 = 5.5; Tukey HSD; P < 0.05), due to the low nauplii production during the first visit to this fjord (Table 2). The production of NI in filtered seawater was low, between 0.5 and 4 NI f−1 d−1.
There was no significant difference in the final development stage of C. finmarchicus between the different fjords (one-way ANOVA; P > 0.05), and >60% of the nauplii reached the first feeding stages (third nauplius stage) on the day 6 of the incubations (Fig. 5c), appearing healthy and actively swimming. The mean development stage on day 6 was 2.6 ± 0.3, 2.7 ± 0.1 and 2.5 ± 0.4 in Malangen, Balsfjord and Ullsfjord, respectively, indicating an average stage duration of 2.3 ± 0.3 days (at ca 3°C). The specific daily mortality rate of the nauplii was 10 ± 3% d−1, and did not differ between the three fjords (Fig. 5d).
The egg production rate did not correlate with any of the food parameters measured or with hatching success (Spearman rank correlation; P > 0.05; Fig. 6a), although egg production was lowest during the first visit to Malangen, when both seston diatom concentration and proportional ingestion of diatoms were extremely high. Hatching success was negatively correlated with diatom concentration (Fig. 6b), and positively correlated with proportion of dinoflagellates in seston (Spearman; P ≤ 0.05, data not shown), although there was no significant correlation between hatching success and proportion of diatoms or hatching success and concentration of dinoflagellates. Further, the production of nauplii (either NI or NIII) did not correlate with diatom abundance or any other food parameter (Fig. 6c).

a Egg production rate (eggs f−1 d−1), b hatching success (%) and c production of nauplii in the first feeding stage (NIII f−1 d−1) of C. finmarchicus as a function of total seston concentration (μg C l−1), diatom concentration (μg C l−1) and proportional abundance (%) of diatoms (mean ± SE). Correlation coefficient and P value for the significant Spearman rank correlation are indicated
Discussion
Little selection—high ingestion of diatoms
Calanus finmarchicus was significantly selecting for diatoms, such as Thalassiosira spp. and Chaetoceros spp., whereas ciliates and dinoflagellates were generally eaten according to their availability, and flagellates (mainly Phaeocystis pouchetii) were selected against. This is in agreement with some spring bloom studies, showing selection for diatoms (Meyer-Harms et al. 1999; Teegarden et al. 2001), but in contrast with others, showing selection for ciliates (Nejstgaard et al. 1997; Irigoien et al. 1998; Levinsen et al. 2000). Possibly the selection for or against diatoms simply depends on the size of the diatom species in question, since the selected diatoms generally seem to be relatively large or chain forming (Chaetoceros spp. and Thalassiosira spp.: Teegarden et al. 2001; Koski and Wexels Riser 2006; this study; Rhizosolenia delicatula: Meyer-Harms et al. 1999). This would support the general idea of C. finmarchicus as mainly a size selective grazer (Harris et al. 2000; Meyer et al. 2002; Koski and Wexels Riser 2006) although selection based on food quality (Huntley et al. 1983), toxicity (Huntley et al. 1986) and even diatom aldehydes (Leising et al. 2005) have been suggested for Calanus spp.
Although colonial forms of Phaeocystis pouchetii seem to be readily ingested by Calanus spp. (Huntley et al. 1987; Tande and Båmstedt 1987), field studies show non-selective feeding on haptophytes in general (Meyer-Harms et al. 1999; Irigoien et al. 2000a). In the present study, however, flagellates (mainly P. pouchetii) were significantly avoided at two out of three fjords, although flagellates still contributed up to 22% of the diet. Negative selection could either be an artefact resulting from possible underestimation of feeding rates on small phytoplankton cells (see below), or due to size selective grazing on larger cells (see Hansen et al. 1994), such as diatoms. Since P. pouchetii colonies were rare or absent during the present study (Wassmann et al. 2005), the contribution of this species to the diet was probably smaller than it would be during episodes of high colony formation.
Relatively small changes in selection, irrespective of larger changes in seston composition, seem to be a general feature of Calanus finmarchicus feeding. In the present study, selection pattern in the different fjords remained more or less the same, as observed in an earlier study with C. finmarchicus copepodites (Koski and Wexels Riser 2006). The inflexibility in selection in both of these studies led to ca 3 times higher ingestion rates at the fjord/time with higher seston concentration. Similarly, no large changes in selective feeding seem to take place between the different phases of the spring phytoplankton bloom, even though an order of magnitude difference in ingestion rates sometimes follow the changes in seston composition and concentration (Nejstgaard et al. 1997; Irigoien et al. 1998; Meyer-Harms et al. 1999; Levinsen et al. 2000). This kind of inflexible selection behaviour and/or non-selective feeding should result in large dependence of seston composition, and suggest that C. finmarchicus reproduction would indeed closely follow the phytoplankton/microzooplankton development (see Diel and Tande 1992). In this respect selective feeding of C. finmarchicus seems to substantially differ from Calanus pacificus, which may change its food selection on a week-to-week basis (Leising et al. 2005).
The ingestion rates measured during the present study were similar to most earlier spring bloom studies, with ingestion ranging from 7 to 23 μg C ind−1 d−1 (Nejstgaard et al. 1997; Irigoien et al. 1998; Meyer-Harms et al. 1999; Levinsen et al. 2000; Teegarden et al. 2001). However, ingestion rate of 7–22 μg C ind−1 d−1 was far too low to support the observed high egg production (9–20 μg C ind−1 d−1), leading to unrealistic gross growth efficiencies close to or above 100%. The disparity between ingestion and egg production could be due to underestimated grazing rates, feeding on non-identified food sources, previous feeding or use of lipid reserves. The high concentration of females incubated (3–7 per 0.6 l bottle) likely led to underestimated ingestion rates, since sometimes ≥50% of the preferred food items (diatoms Chaetoceros spp. and Thalassiosira spp.) were consumed during the incubation. Then, part of the eggs should have been produced either with reserve lipids or based on previous ingestion. Exactly similar incubations in filtered seawater, however, produced only very few eggs, demonstrating a very fast (<24 h) response to starvation. Since C. finmarchicus is not sensitive to short-term starvation (Dagg 1977), it seems unlikely that the egg production would stop immediately due to depleted reserves. Possibly some kind of physiological mechanism slows down the egg production in the absence of food, irrespective of the previous good feeding conditions or reserve lipids, although no evidence exists to support this assumption. In any case, recent feeding seemed to be necessary to maintain a high egg production rate in the incubations.
Another possibility for underestimation of ingestion rates are the unidentified or underestimated food sources, such as detritus, picoplankton or small flagellates. Since I did not account for the possible effect of ciliate feeding in incubation bottles (Nejstgaard et al. 2001a), ingestion rates on small flagellates could have been underestimated. However, since the ciliate concentration especially at Malangen and Balsfjord was very low (1–2 cells ml−1), and since picoplankton and detritus are not generally considered to be high quality food sources for copepod egg production, it seems unlikely that underestimation of these items would account for the missing ingestion.
The most likely explanation for the discrepancy between ingestion and egg production rates seems therefore to be that part of the eggs were produced based on previous feeding, with a possible small contribution from lipid reserves (see Niehoff et al. 1999; Richardson et al. 1999). Since the faecal pellet production of C. finmarchicus females remained relatively stable between the sampling times (1.6 ± 0.6 pellets f−1 h−1) indicating rather stable ingestion rates (C. Wexels Riser, University of Tromsø; unpublished data), since C. finmarchicus did not seem to adjust its selective feeding behaviour to changes in seston composition (see above), and since the co-dominance of diatoms and Phaeocystis seems to prevail during the 1–1.5 months spring bloom period (Lutter et al. 1989), the proportional ingestion of different food groups in incubations should rather well have represented the ingestion on which the egg production was based on (with likely underestimation of diatom ingestion). However, the ingestion rates presented here should be considered as minimum estimates. This probably accounts for most of the feeding studies with adult C. finmarchicus during the reproductive season: if C. finmarchicus produces ca 40 eggs f−1 d−1 (Mauchline 1998), the corresponding ingestion should exceed 38 μg C ind−1 d−1 (with a gross growth efficiency of 30%; Straile 1997). Such high ingestion rates, which are sufficient to account for a moderate to high egg production, have only been presented by Ohman and Runge (1994) for the post-bloom season of the productive St. Lawrence estuary.
Diatoms and reproduction
Despite of the high proportional feeding on diatoms, egg production and hatching success of C. finmarchicus were high, and nauplii developed fast past the first non-feeding stages. The egg production rates observed here (40–90 eggs f−1 d−1) were similar to maximum rates reported for C. finmarchicus in literature (Hirche 1990; Niehoff et al. 1999; Richardson et al. 1999), and higher than what would be expected at the corresponding temperatures with superabundant food (≤40 eggs f−1 d−1; Hirche et al. 1997). Similarly, the hatching rates corresponded to the upper range of previous observations (Jónasdóttir et al. 2002, 2005), and nauplii development was faster than in laboratory experiments with superabundant food (Campbell et al. 2001). The production of nauplii (either NI or NIII) was high, and not connected to the abundance or proportion of diatoms, although the diatom concentration was sufficiently high to induce adverse effects (see Chaudron et al. 1996; Leising et al. 2005). My results are therefore different from those of Nejstgaard et al. (1997, 2001b), Miralto et al. (2003) and Poulet et al. (2006), showing a strong reduction of egg production and/or hatching success of Calanus spp. during diatom dominated blooms, but more similar to observations by Irigoien et al. (2000a, b) and Jónasdóttir et al. (2002) where high egg production and/or hatching success was associated with high diatom abundance. The diatom-copepod interactions seem therefore to be both species (C. finmarchicus/C. helgolandicus) and population (C. finmarchicus in Tromsø/C. finmarchicus in Bergen) specific.
I did, however, observe a significant negative correlation between hatching success and diatom abundance, and lower (although still high) egg production, hatching success and nauplii production at the station where proportion of diatoms both in seston and in the diet were highest. This could be either due to harmful aldehydes observed in some diatom species (Ianora et al. 2003; Pohnert 2005; Wichard et al. 2005), one-sided diet leading to nutritional deficiency (Jónasdóttir et al. 1998; Jones and Flynn 2005), or both. Since I measured neither aldehydes, nor biochemical or mineral components of the copepod diet, the reason for the decreased hatching in the present study remains unclear. The only diatoms which could potentially have produced aldehydes were Thalassiosira spp., which were sometimes ingested in large quantities. The Thalassiosira species which contributed mostly to the total Thalassiosira biomass during the cruise were T. nordenskioeldii, T. antarctica, T. angulata and T. anguste-lineata (P. Wassmann, University of Tromsø, unpublished data); of these species T. nordenskioeldii and T. anguste-lineata are listed as potential aldehyde producers (Wichard et al. 2005). By assuming similar aldehyde concentrations as found by Wichard et al. (2005), we can estimate the range of diatom aldehyde ingestion to be 0.0003–0.02 μg ind−1 d−1, assuming 100% of the ingested Thalassiosira spp. to be T. nordenskioeldii or T. anguste-lineata, respectively. Similarly, the range of aldehyde concentration in the media would be 0.002–0.13 μg ml−1, assuming a 100% concentration of T. nordenskioeldii and T. anguste-lineata, respectively. Ianora et al. (2004) report strong adverse effects of maternal ingestion of 0.0001 μg aldehydes d−1, suggesting that sufficient amount of aldehydes could have been ingested by C. finmachicus for adverse effects to occur, although the seston concentrations appear to be in the lower range of concentrations reported to have harmful effects. However, since aldehyde production among diatoms are species and even strain specific (Pohnert et al. 2002), the effect of aldehydes on hatching success here remains speculative.
Similarly, there was no clear evidence of effects of biochemical composition (Jónasdóttir et al. 1998), mineral content (Jones and Flynn 2005) or algae senescence (Jónasdóttir et al. 2002) on hatching success. I did not observe a superiority of any taxonomic group for C. finmarchicus reproduction (assuming that different algae groups would have different biochemical content; see Brown et al. 1997), and, since the inorganic nutrients were not depleted during the cruise (P. Wassmann, University of Tromsø, unpublished data), there was no reason to assume that the algae would have been senescent. However, a diverse diet seemed to be profitable: a total ingestion which was more equally divided between different taxonomic groups did result in a higher reproductive success (Ullsfjord > Balsfjord > Malangen). This agrees with the general idea of a superiority of mixed diets and high diet diversity for copepod production (Kleppel 1993).
Population egg production
Irrespective of the reason for the slightly decreased hatching success, if compared to density-dependent predation (or cannibalism) mortality of freely spawned eggs (up to 99%; Peterson and Kimmerer 1994; Ohman and Hirche 2001), a maximum increase of 15% in egg mortality due to the diet does not seem to be a high number. If combined with egg production, however, the 15% decrease in hatching at the first visit to Malangen resulted in nearly three times lower production of nauplii than in other fjords and sampling times and coincided with the lowest population egg production observed during the study (Table 2). It remains to be speculated if (1) this one-day dip in population egg and nauplii production is of great consequence for C. finmachicus production and (2) if it is primarily caused by the harmful effects of diatoms. Since (1) population egg production was primarily determined by female abundance, in turn affected by e.g., advection of zooplankton to the area (Aksnes et al. 1989; Falkenhaug et al. 1997a, b), (2) no correlation was observed between nauplii production and diatom abundance/proportion, and (3) the population egg production was high at stations/times with only slightly lower abundance/proportion of diatoms (see Table 1), the negative effect of diatoms does not seem to be convincingly demonstrated in the present study. Further, the overall high reproduction, irrespective of high biomass and ingestion of diatoms, seem to suggest a generally successful recruitment, although a lower reproductive rate can sometimes be observed.
The average population egg production amounted to >1,20,000 eggs m−2 d−1 in Malangen and Balsfjord (1,27,000 ± 76,000 eggs m−2 d−1) and to >60,000 eggs m−2 d−1 in Ullsfjord (60,700 ± 20,030 eggs m−2 d−1; Table 2), close to the maximum rates reported in Norwegian Sea (Harris et al. 2000). Assuming the peak in reproductive season to last for 3 weeks (Tande 1991), up to 2.7 million eggs m−2 would be produced. In contrast to high pre-bloom population egg production observed in the Norwegian Sea (Niehoff et al. 1999), it appears that in north Norwegian fjords most of the pre-bloom females are immature (Tande 1982), and that only the onset of the spring phytoplankton bloom in the beginning of April triggers the final ovary maturation (Tande 1991; Diel and Tande 1992). Although the total C. finmarchicus biomass in, e.g. Balsfjord, in February may be three times higher than in April (Barthel 1995), it is unlikely to compensate for the low pre-bloom egg production and low percentage of mature females in the population. Assuming a pre-bloom egg production of <10 eggs f−1 d−1 (Niehoff et al. 1999), ca 25% of the population to be females (Tande 1982) and a 1.5 month pre-bloom season (see Tande 1982), the maximum pre-bloom population egg production would be 21,000 eggs m−2 d−1 and total pre-bloom egg production 9,00,000 eggs m−2, ca 5 times lower than the population egg production during the peak season. Further, since the stage IV gonads actually only start to appear in early April (Tande 1982), these numbers are likely to be overestimated. Therefore, a substantial part of the annual reproduction seems to be produced during the spring phytoplankton bloom, which stresses the importance of spring diatom bloom for population dynamics of C. finmarchicus in northern Norwegian fjords. Seasonal studies on the population dynamics of C. finmarchicus confirm this: high abundance of nauplii and subsequently copepodites and adults follows the peak reproductive season (Hopkins et al. 1984; Tande and Slagstad 1991). Diatoms therefore do not seem to have substantial negative effects on the population dynamics of C. finmachicus in these northern ecosystems.
References
Aksnes DL, Aure J, Kaartvedt S, Magnesen T, Richard J (1989) Significance of advection for the carrying capacities of fjord populations. Mar Ecol Prog Ser 50:263–274
Arashkevich EG, Tande KS, Pasternak AE, Ellertsen B (2004) Seasonal moulting patterns and the generation cycle of Calanus finmarchicus in the NE Norwegian Sea, as inferred from gnathobase structures, and the size of gonads and oil sacs. Mar Biol 146:119–132
Ban S, Burns C, Castel J, Chaudron Y, Christou E, Escribano R, Umani SF, Gasparini S, Ruiz FG, Hoffmeyer M, Ianora A, Kang H-K, Laabir M, Lacoste A, Miralto A, Ning X, Poulet S, Rodriguez V, Runge J, Shi J, Starr M, Uye S, Wang Y (1997) The paradox of diatom–copepod interactions. Mar Ecol Prog Ser 157:287–293
Barthel K-G (1995) Zooplankton dymanics in Balsfjorden, northern Norway. In: Skjoldal HR, Hopkins C, Erikstad KE, Leinaas HP (eds) Ecology of fjords and coastal waters. Elsevier, Amsterdam, pp 113–126
Brown MR, Jeffrey SW, Volkman JK, Dunstan GA (1997) Nutritional properties of microalgae for mariculture. Aquaculture 151:315–331
Campbell RW, Head EJH (2000) Egg production rates of Calanus finmarchicus in the western North Atlantic: effect of gonad maturity, female size, chlorophyll concentration and temperature. Can J Fish Aquat Sci 57:518–529
Campbell RG, Wagner MM, Teegarden GJ, Boudreau CA, Durbin EG (2001) Growth and development rates of the copepod Calanus finmarchicus reared in the laboratory. Mar Ecol Prog Ser 221:161–183
Chaudron Y, Poulet SA, Laabir M, Ianora A, Miralto A (1996) Is hatching success of copepod eggs diatom density-dependent? Mar Ecol Prog Ser 144:185–193
Chesson J (1978) Measuring preference in selective predation. Ecology 59:211–215
Chesson J (1983) The estimation and analysis of preference and its relationship to foraging models. Ecology 64:1297–1304
Corkett CJ, McLaren IA, Sevigny J-M (1986) The rearing of the marine calanoid copepods Calanus finmarchicus (Gunnerus), C. glacialis Jaschnov and C. hyperboreus Kroyer with comment on the equiproportional rule. Syllogeus 58:539–546
Cowles TJ, Olson RJ, Chisholm SW (1988) Food selection by copepods: discrimination on the basis of food quality. Mar Biol 100:41–49
Dagg M (1977) Some effects of patchy food environments on copepods. Limnol Oceanogr 22:99–107
Davis CS (1984) Predatory control of copepod seasonal cycles on Georges Bank. Mar Biol 82:31–40
Diel S, Tande K (1992) Does the spawning of Calanus finmarchicus in high latitudes follow a reproducible pattern. Mar Biol 113:21–31
Durbin EG, Campbell RG, Casas MC, Ohman MD, Niehoff B, Runge J, Wagner M (2003) Interannual variation in phytoplankton blooms and zooplankton productivity and abundance in the Gulf of Maine during winter. Mar Ecol Prog Ser 254:81–100
Falkenhaug T, Tande K, Semenova T (1997a) Diel, seasonal and ontogenetic variations in the vertical distribution of four marine copepods. Mar Ecol Prog Ser 149:105–119
Falkenhaug T, Tande K, Timonin A (1997b) Spatio-temporal patterns in the copepod community in Malangen, northern Norway. J Plankton Res 19:449–468
Frost BW (1972) Effects of size and concentration of food particles on the feeding behaviour of the marine planktonic copepod Calanus pacificus. Limnol Oceanogr 17:805–815
Halsband-Lenk C, Pierson JJ, Leising AW (2005) Reproduction of Pseudocalanus newmani (Copepoda: Calanoida) is deleteriously affected by diatom blooms—a field study. Prog Oceanogr 67:332–348
Hansen B, Verity P, Falkenhaug T, Tande KS, Norrbin F (1994) On the trophic fate of Phaeocystis pouchetii (Harriot). 5. Trophic relationships between Phaeocystis and zooplankton: An assessment of methods and size dependence. J Plankton Res 16:487–511
Harris RP, Irigoien X, Head RN, Rey C, Hygum BH, Hansen BW, Niehoff B, Meyer-Harms B, Carlotti F (2000) Feeding, growth, and reproduction in the genus Calanus. ICES J Mar Sci 57:1708–1726
Hirche H-J (1990) Egg production of Calanus finmarchicus at low temperature. Mar Biol 106:53–58
Hirche H-J, Meyer U, Niehoff B (1997) Egg production of Calanus finmarchicus: effect of temperature, food and season. Mar Biol 127:609–620
Hopkins CCE, Tande KS, Grønvik S (1984) Ecological investigations of the zooplankton community of Balsfjorden, northern Norway: an analysis of growth and overwintering tactics in relation to niche and environment in Metridia longa (Lubbock), Calanus finmarchicus (Gunnerus), Thysanoessa inermis (Krøyer) and T. raschi (M. Sars). J Exp Mar Biol Ecol 82:77–99
Horner R, Postel JR, Halsband-Lenk C, Pierson JJ, Pohnert G, Wichard T (2005) Winter-soring phytoplankton blooms in Dabob Bay, Washington. Prog Oceangr 67:286–313
Huntley M (1981) Nonselective, nonsaturated feeding by three calanid copepod species in the Labraror Sea. Limnol Oceanogr 26:831–842
Huntley ME, Barthel K-G, Star JL (1983) Particle rejection by Calanus pacificus: discrimination between similarly sized particles. Mar Biol 74:151–160
Huntley ME, Sykes P, Rohan S, Marin V (1986) Chemically-mediated rejection of dinoflagellate prey by the copepod Calanus pacificus and Paracalanus parvus: mechanism, occurrence and significance. Mar Ecol Prog Ser 28:105–120
Huntley M, Tande K, Eilertsen HC (1987) On the trophic fate of Phaeocystis pouchetii (Hariot). 2. Grazing rates of Calanus hyperboreus (Kroeyer) on diatoms and different size categories of Phaeocystis pouchetii. J Exp Mar Biol Ecol 110:197–212
Ianora A, Poulet SA, Miralto A (2003) The effects of diatoms on copepod reproduction: a review. Phycologia 42:351–363
Ianora A, Miralto A, Poulet SA, Carotenuto Y, Buttino I, Romano G, Casotti R, Pohnert G, Wichard T, Colucci-D’Amato L, Terrazzano G, Smetacek V (2004) Aldehyde suppression of copepod recruitment in blooms of a ubiquitous planktonic diatom. Nature 429:403–407
Irigoien X, Head R, Klenke U, Meyer-Harms B, Harbour D, Niehoff B, Hirche H-J, Harris R (1998) A high frequency time series at weathership M, Norwegian Sea, during the 1997 spring bloom: feeding of adult female Calanus finmarchicus. Mar Ecol Prog Ser 172:127–137
Irigoien X, Head RN, Harris RP, Cummings D, Harbour D, Meyer-Harms B (2000a) Feeding selectivity and egg production of Calanus helgolandicus in the English Channel. Limnol Oceanogr 45:44–54
Irigoien X, Harris RP, Head RN, Harbour D (2000b) The influence of diatom abundance on the egg production rate of Calanus helgolandicus in the English Channel. Limnol Oceanogr 45:1433–1439
Irigoien X, Harris RP, Verheye HM, Joly P, Runge J, Starr M, Pond D, Cambell R, Shreeve R, Ward P, Smith AN, Dam HG, Peterson W, Tirelli V, Koski M, Smith T, Harbour D, Davidson R (2002) Copepod hatching success in marine ecosystems with high diatom concentrations. Nature 419:387–389
Jónasdóttir SH, Kiørboe T, Tang KW, St. John M, Visser AW, Saiz E, Dam HG (1998) Role of diatoms in copepod production: Good, harmless or toxic? Mar Ecol Prog Ser 172:305–308
Jónasdóttir SH, Gudfinnsson HG, Gislason A, Astthorsson OS (2002) Diet composition and quality for Calanus finmarchicus egg production and hatching success off south-west Iceland. Mar Biol 140:1195–1206
Jónasdóttir SH, Trung NH, Hansen F, Gärtner S (2005) Egg production and hatching success in the calanoid copepods Calanus helgolandicus and Calanus finmarchicus in the North Sea from March to September 2001. J Plankton Res 27:1–21
Jones RH, Flynn KJ (2005) Nutritional status and diet composition affect the value of diatoms as copepod prey. Science 307:1457–1459
Klein Breteler W, Koski M, Rampen S (2004) Role of essential lipids in copepod nutrition: no evidence for trophic upgrading of food quality by a marine ciliate. Mar Ecol Prog Ser 274:199–208
Kleppel GS (1993) On the diets of calanoid copepods. Mar Ecol Prog Ser 99:183–195
Koski M, Wexels Riser C (2006) Post-bloom feeding of Calanus finmarchicus copepodites: selection for autotrophic vs. heterotrophic prey. Mar Biol Res 2:109–119
Leising AW, Pierson JJ, Halsband-Lenk C, Horner R, Postel J (2005) Copepod grazing during spring blooms: Does Calanus pacificus avoid harmful diatoms? Prog Oceanogr 67:384–405
Levinsen H, Turner JT, Nielsen TG, Hansen BW (2000) On the trophic coupling between protists and copepods in arctic marine ecosystems. Mar Ecol Prog Ser 204:65–77
Lutter S, Taasen JP, Hopkins CCE, Smetacek V (1989) Phytoplankton dynamics and sedimentation processes during spring and summer in Balsfjord, northern Norway. Polar Biol 10:113–124
Mauchline J (1998) The biology of calanoid copepods. Academic, San Diego
Meyer-Harms B, Irigoien X, Head R, Harris R (1999) Selective feeding on natural phytoplankton by Calanus finmarchicus before, during, and after the 1997 spring bloom in the Norwegian Sea. Limnol Oceanogr 44:154–165
Meyer B, Irigoien X, Graeve RN, Harris RP (2002) Feeding rates and selectivity among nauplii, copepodites and adult females of Calanus finmarchicus and Calanus helgolandicus. Helgol Mar Res 56:169–176
Miralto A, Barone G, Romano G, Poulet SA, Ianora A, Russo GL, Buttino I, Mazzarellas G, Laabir M, Cabrini M, Giacobbe MG (1999) The insidious effect of diatoms on copepod reproduction. Nature 402:173–176
Miralto A, Guglielmo L, Zagami G, Buttino I, Granata A, Ianora A (2003) Inhibition of population growth in the copepods Acartia clausi and Calanus helgolandicus during diatom blooms. Mar Ecol Prog Ser 254:253–268
Montagnes DJS, Berges JA, Harrison PJ, Taylor FJR (1994) Estimating carbon, nitrogen, protein, and chlorophyll a from volume in marine phytoplankton. Limnol Oceanogr 39:1044–1060
Nejstgaard JC, Gismervik I, Solberg PT (1997) Feeding and reproduction by Calanus finmarchicus, and microzooplankton grazing during mesocosm blooms of diatoms and the coccolithophore Emiliania huxley. Mar Ecol Prog Ser 147:197–217
Nejstgaard JC, Naustvoll L-J, Sazhin A (2001a) Correcting for underestimation of microzooplankton grazing in bottle incubation experiments with mesozooplankton. Mar Ecol Prog Ser 221:59–75
Nejstgaard JC, Hygum BH, Naustvoll L-J, Båmstedt U (2001b) Zooplankton growth, diet and reproductive success compared in simultaneous diatom- and flagellate-microzooplankton-dominated plankton blooms. Mar Ecol Prog Ser 221:77–91
Niehoff B, Klenke U, Hirche H-J, Irigoien X, Head R, Harris R (1999) A high frequency time series at Weathership M, Norwegian Sea, during the 1997 spring bloom: the reproductive biology of Calanus finmarchicus. Mar Ecol Prog Ser 176:81–92
Ohman MD, Runge JA (1994) Sustained fecundity when phytoplankton resources are in short supply: Omnivory by Calanus finmarchicus in the Gulf of St. Lawrence. Limnol Oceanogr 39:21–36
Ohman MD, Hirche H-J (2001) Density-dependent mortality in an oceanic copepod population. Nature 412:638–641
Pasternak A, Arashkevich E, Wexels Riser C, Ratkova T, Wassmann P (2000) Seasonal variation in zooplankton and suspended faecal pellets in the subarctic Norwegian Balsfjorden, in 1996. Sarsia 85:439–452
Pasternak A, Tande KS, Arashkevich E, Melle W (2004) Reproductive patterns of Calanus finmarchicus at the Norwegian midshelf in 1997. J Plankton Res 26:839–849
Peterson WT, Kimmerer WJ (1994) Processes controlling recruitment of the marine calanoid copepod Temora longicornis in Long Island Sound: Egg production, egg mortality, and cohort survival rates. Limnol Oceanogr 39:1594–1605
Pierson JJ, Halsband-Lenk C, Leising AW (2005) Reproductive success of Calanus pacificus during diatom blooms in Dabob Bay, Washington. Prog Oceanogr 67:314–331
Poulet SA, Laabir M, Ianora A, Miralto A (1995) Reproductive response of Calanus helgolandicus. I. Abnormal embryonic and naupliar development. Mar Ecol Prog Ser 129:85–95
Poulet SA, Richer de Forges M, Cueff A, Lennon J-F (2003) Double-labelling methods used to diagnose apoptic and necrotic cell degradations in copepod nauplii. Mar Biol 143:889–895
Poulet SA, Wichard T, Ledoux JB, Lebreton B, Marchetti J, Dancie C, Bonnet D, CuefF A, Morin P, Pohnert G (2006) Influence of diatoms on copepod reproduction. I. Field and laboratory observations related to Calanus helgolandicus egg production. Mar Ecol Prog Ser 308:129–142
Pohnert G (2005) Diatom/copepod interactions in plankton: the indirect chemical defence of unicellular algae. Chembiochem 6:946–959
Pohnert GB, Lumineau O, Cueff A, Adolph S, Cordevant C, Lange M, Poulet S (2002) Are volatile unsaturated aldehydes from diatoms the main line of chemical defence against copepods? Mar Ecol Prog Ser 245:33–45
Pond D, Harris R, Head R, Harbour D (1996) Environmental and nutritional factors determining seasonal variability in the fecundity and egg viability of Calanus helgolandicus in coastal waters off Plymouth, UK. Mar Ecol Prog Ser 143:45–63
Reigstad M, Wassmann P, Ratkova T, Arashkevich E, Pasternak A, Øygarden S (2000) Comparison of springtime vertical export of biogenic matter in three northern Norwegian fjords. Mar Ecol Prog Ser 201:73–89
Richardson K, Jónasdóttir SH, Hay SJ, Christoffersen A (1999) Calanus finmarchicus egg production and food availability in the Faroe-Shetland Channel and northern North Sea: October-March. Fish Oceanogr 8:153–162
Rousseau V, Leynaert A, Daoud N, Lancelot C (2002) Diatom succession, silification and silicic acid availability in Belgian coastal waters (Southern North Sea). Mar Ecol Prog Ser 236:61–73
Starr M, Runge JA, Therriault J-C (1999) Effects of diatom diets on the reproduction of the planktonic copepod Calanus finmarchicus. Sarsia 84:379–389
Straile D (1997) Gross growth efficiencies of protozoan and metazoan zooplankton and their dependence on food concentration, predator-prey weight ratio, and taxonomic group. Limnol Oceanogr 42:1375–1385
Tande KS (1982) Ecological investigations of the zooplankton community of Balsfjorden, northern Norway: generation cycles, and variations in body weight and body content of carbon and nitrogen related to overwintering and reproduction in the copepod Calanus finmarchicus (Gunnerus). J Exp Mar Biol Ecol 62:129–142
Tande KS (1991) Calanus in North Norwegian fjords and in the Barents Sea. Polar Res 10:389–407
Tande KS, Båmstedt U (1987) On the trophic fate of Phaeocystis pouchetii. 1. Copepod feeding rates on solitary cells and colonies of P. pouchetii. Sarsia 72:313–320
Tande KS, Slagstadt D (1991) Regional and interannual variation in biomass and productivity of Calanus finmarchicus in subarctic environments. Oceanologica Acta 15:309–312
Tang KW, Jakobsen HH, Visser AW (2001) Phaeocystis globosa (Prymnesiophyceae) and the planktonic food web: feeding, growth, and trophic interactions among grazers. Limnol Oceanogr 46:1860–1870
Teegarden GJ, Campbell RG, Durbin EG (2001) Zooplankton feeding behavior and particle selection in natural plankton assemblages containing toxic Alexandrium spp. Mar Ecol Prog Ser 218:213–226
Verity PG, Smayda TJ (1989) Nutritional value of Phaeocystis pouchetii (Prymnesiophyceae) and other phytoplankton for Acartia spp. (Copepoda). Ingestion, egg production, and growth of nauplii. Mar Biol 100:161–171
Wassmann P, Svendsen H, Keck A, Reigstad M (1996) Selected aspects of the physical oceanography and particle fluxes in fjords of northern Norway. J Mar Syst 8:53–71
Wassmann P, Ratkova T, Reigstad M (2005) The contribution of single and colonial cells of Phaeocystis pouchetii to spring and summer blooms in the north-eastern North Atlantic. Harmful Algae 4:823–840
Wichard T, Poulet SA, Halsband-Lenk C, Albaina A, Harris R, Liu D, Pohnert G (2005) Survey of the chemical defence potential of diatoms: screening of fifty one species for α, ß, γ, δ-unsaturated aldehydes. J Chem Ecol 31:1–10
Acknowledgments
I wish to thank Paul Wassmann for the organisation of the cruise, and P. Wassmann and Christian Wexels Riser for providing the unpublished data on chlorophyll-a, inorganic nutrients, phytoplankton species composition, copepod abundance and copepod faecal pellet production. Elena Arashkevich, Sigrun Jónasdóttir, Serge Poulet and C. Wexels Riser kindly commented the previous versions of the manuscript. Jörg Dutz assisted with the sampling and sorting of copepods. This study was financially supported by Academy of Finland, Carlsberg Foundation and a mobility grant from Nordic Academy for Advanced Study (NorFA), and was conducted during a cruise of the EU-project BIOHAB.
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by M. Kühl.
Rights and permissions
About this article
Cite this article
Koski, M. High reproduction of Calanus finmarchicus during a diatom-dominated spring bloom. Mar Biol 151, 1785–1798 (2007). https://doi.org/10.1007/s00227-007-0615-2
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00227-007-0615-2