
Abstract
The gonosomatic or gonadosomatic index [GSI=(ovary weight/fish weight)×100] has been widely used in fisheries science and experimental reproductive studies as a simple, low-cost measure of reproductive condition. However, its properties have not been fully evaluated, and several pitfalls, such as size-dependence and changes in ovarian allometry, may invalidate its use. In the present study, we examined ovarian allometry and the appropriateness of GSI for assessing ovarian activity in the Mediterranean sardine Sardina pilchardus. The analysis was based on a large sample of histologically scored females collected over an annual cycle in the eastern Mediterranean (Aegean and Ionian Seas). First, we examined GSI dependence on fish size by comparing ovary weight–on–fish weight relationships in different stages of oocyte development. The effects of recent spawning (incidence of postovulatory follicles) and intensity of follicular atresia were also addressed. In a subsequent step of the analysis, we applied generalized linear models (GLMs) to analyze the relationship between GSI and histological stage, taking into account the parallel effects of additional factors related to geographical region, month of capture, fish length and energetic reserves (fat stage, hepatosomatic index—HSI). Ovarian growth was isometric in all stages of oocyte development and states of atresia, but altered to positive allometric at the stage of oocyte hydration. Oocyte growth and intensity of atresia significantly affected GSI. Fish length, geographical region, month of capture, fat stage and HSI did not substantially affect GSI, further strengthening the appropriateness of the index for the Mediterranean sardine. Finally, we provide first evidence, based mainly on available (but yet limited) published information for other fish species, that: (1) the pattern of ovarian allometry may not be altered by fecundity variations and (2) the pattern of ovarian allometry may reflect the pattern of oocyte growth (i.e. isometric vs. allometric ovarian growth could reflect group-synchronous vs. -asynchronous oocyte development). Alternatively, the pattern of ovarian allometry could reflect the presence or absence of size dependency in the relative fecundity (eggs per gram of body weight) and/or egg size. The isometric ovarian growth in the Mediterranean sardine could be attributed to the lack of such size-dependent effects.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Fish reproductive cycles are coupled with pronounced changes in gonadal size. For a given stage of development, gonadal size also increases with fish size. A widely used method to index reproductive condition has been to express gonadal weight as a percentage of body weight (DeVlamming et al. 1982). The resulting gonosomatic or gonadosomatic index (GSI) provides a simple measure of the extent of gonadal development, alternative to more precise but laborious methods such as gonadal histology or oocyte diameter measurements (West 1990).
Despite its wide use, the validity of GSI has often been questioned (DeVlamming et al. 1982; Erickson et al. 1985b; Garcia-Berthou and Moreno-Amich 1993; DeMartini and Lau 1999). It has generally been recommended that the index should not be used without proper validation (DeVlamming et al. 1982). The most serious problem is that, for many fish species, GSI is correlated with fish size (West 1990). Consequently, its use in among-sample comparisons will produce biased estimates of reproductive condition when samples are composed of different-sized individuals. A more general model, the relative gonadal index (RGI), then, would be more appropriate; however, the allometric coefficient of the gonad weight–on–fish weight relationship should not change with stage of gonadal development (Erickson et al. 1985b).
When addressing the question of GSI validity for female fish, it is necessary to consider the main factors that determine ovarian weight, namely, the stage of oocyte development (mean oocyte size) and fish fecundity (DeVlamming et al. 1982; Jons and Miranda 1997). Both ovarian maturation and fish fecundity are linked to energy reserves and, hence, food supply. Many fish species transfer energy from stored fat to reproductive tissue, but others primarily utilize energy stored in the liver during reproduction (Milton et al. 1994, and references therein). Energy availability can cause variations in egg production (e.g. Coward and Bromage 1999; Marshall et al. 1999; Somarakis 2004), with likely impacts on ovarian allometry.
In the present study, we validated the use of GSI for the Mediterranean sardine, Sardina pilchardus. The analysis was based on a large sample of histologically scored females collected over an annual cycle in the eastern Mediterranean (Aegean and Ionian Seas). First, we examined GSI dependence on fish size. The effects of oocyte development, recent spawning and intensity of follicular atresia were considered. In a subsequent step of the analysis, we applied generalized linear models to analyze the relationship between GSI and histological stage, taking into account the parallel effects of additional factors related to geographical region, season, fish size and energetic reserves.
Materials and methods
Sample collection and laboratory analysis
Samples of adult sardines (n=134) were collected on a monthly basis from coastal locations of central Greece, from September 1999 to August 2000. Sampling was carried out on board the commercial fleet of Chalkis and Piraeus, operating in the central Aegean Sea, and Patras, operating in the central Ionian Sea (Fig. 1). Each sample consisted of a random collection of 1.5–2 kg of sardines. Fish were fixed onboard in 10% neutral buffered formalin. Sample information is provided in Table 1.

Map of the central Aegean and Ionian Seas (eastern Mediterranean) showing the sardine sampling ports
In the laboratory, 15–20 randomly selected females from each sample (or all females when their number was <15) were measured (total length, 0.1 cm). Gonads, liver and viscera were removed, and gutted weights (W) were recorded (0.01 g). The amount of visceral fat was assessed macroscopically according to a seven-point scale based on the staging system of Nikolsky (1963), slightly modified for sardine (Table 2). Both ovaries and liver were dried of surface moisture and weighed (0.1 mg). Gonosomatic and hepatosomatic indices were calculated as the ratio of gonad (Wg) and liver (Wl) weight to gutted weight, respectively, expressed as a percentage, i.e. GSI=[(Wg/W)×100] and HSI=[(Wl/W)×100]. Somatic indices based on gutted, rather than total fish weight, were considered more appropriate, because they do not include gonad and/or liver weight in either numerator and denominator and are not influenced by the amount of food in fish stomachs.
Histological analysis
A total of 1467 females were examined histologically. A piece of tissue was removed from the center of each ovary, dehydrated, cleared in xylol and embedded in paraffin. Sections (4–6 μm) were cut and stained with Mayer’s hematoxylin and eosin Y (Clark 1981). Histological scoring included the maturation stage of the most advanced batch of oocytes (Murayama et al. 1994; Ganias et al. 2004), the state of atresia [atretic states 0, 1, 2 and 3 according to Hunter and Macewitz (1985)], and the incidence of postovulatory follicles (POFs; Ganias et al. 2003). The ovaries were classified into a total of 18 histological stages (Table 3). A total of 917 females (63%) had ovaries without any atretic or postovulatory follicles. Ovulated females were very limited in number (n=4, Table 3), and they were not considered in the subsequent analyses.
The allometry of ovaries
According to allometric theory (Gould 1966), in a random sample of j=1,...n i fish from some population in histological stage i, ovarian weight, Wgij, and fish weight, W ij , follow the relationship:
where a i and b i are parameters and e j are normally distributed random errors with a mean of 0 and a variance of σ2. Therefore, exp(e j ) has a log-normal distribution. From standard properties of the log-normal distribution (Gutreuter 1990), the conditional expectation (E) of Wgij given W ij is:
and the expectation of the relative gonadal index [or the traditionally used GSI in the case of isometry (b i =1)]:
We estimated a i and b i on a logarithmic scale for each histological stage i:
and examined whether GSI was dependent on somatic weight by testing for isometry, i.e. b i =1 (t-tests, Erickson et al. 1985b). Furthermore, ANCOVA models were used to test for among-stage differences in the regression parameters. Among-stage differences in mean GSI (log-transformed values) were tested with ANOVA followed by Newman–Keuls multiple comparisons.
GLM analysis
In a subsequent step, generalized linear models (GLMs; McCullagh and Nelder 1983) were used to analyze variations in GSI and the relative importance of different variables in explaining those variations. The predictor variables considered were: histological stage, region (Aegean Sea—Ionian Sea), month of capture, fish length, fat stage (Table 2) and HSI.
The choice of the most appropriate link function and error distribution was made on the basis of residual plots. A log-link function was found to be adequate in all cases, implying that explanatory variables had a multiplicative effect on the response parameter. A gamma error structure model was the most appropriate for the analysis of the data.
Variable selection proceeded by forward entry, and the Akaike information criterion (AIC; Chambers and Hastie 1992) was used to detect the relative importance of each variable in explaining variations and to determine which variables should be included in the final model and in which order. The AIC statistic accounts simultaneously for the degrees of freedom used and the goodness of fit, such that a smaller AIC statistic corresponds to a better model in the sense of smaller residual deviance penalized by the number of parameters that are estimated in fitting the model. This approach facilitated the creation of a model that included only the most important (in terms of significance and explanatory power) variables. Analysis of deviance to evaluate the significance (F-test) of the variables included in the final model was also performed by a stepwise procedure.
The models were fitted by means of the statistical package S-PLUS as detailed by Venables and Ripley (1997).
Results
Gonadal allometry and GSI by histological stage
The weight of the ovary was significantly related to body weight in all histological stages, except stage 6 (i.e. in ovaries at the primary yolk globule stage with state-1 atresia) (Table 4). All models had very good fit to data in terms of residuals (homoscedastic, normally distributed). The slopes b i for all stages did not differ significantly from unity, except for the stage of hydration (stage 17), where a strong positive allometry was found (Table 4). This is illustrated in Fig. 2 for all stages of ovarian development, with no incidence of atresia and POFs (stages 1, 3, 5, 12, 14, 16 and 17; see Table 3). Thus, prior to hydration, gonadal growth in sardine was isometric and GSI was independent of body weight.

Sardina pilchardus. Regression lines for the relationships of ovarian weight–on–body weight for different stages of ovarian development
Excluding hydrated females, which presented allometric ovarian growth, slopes of all remaining models (histological stages 1–16) were homogeneous, but intercepts differed significantly (ANCOVA models—slopes: F=1.29, P>0.05; intercepts: F=498.28, P<<0.01). In each individual histological stage, GSI values were log-normally distributed (Kolmogorov–Smirnov tests, P>0.05 in all cases), which was in agreement with GSI expectation (see “Materials and methods” section). Analysis of variance on the log-transformed GSI values (F=499.63, P<<0.01) and subsequent a posteriori comparisons (Table 5) showed that:
-
1.
In females without POFs and/or atresia (comparison of stages 1, 3, 5, 12, 14 and 16), the stage of development of the advanced clutch had a significant effect on GSI up to the tertiary yolk-globule stage (end of vitellogenesis) (see also Fig. 2). Mean GSI of tertiary yolk globule females (stage 14) did not differ significantly from migratory nucleus females (stage 16).
-
2.
In each individual stage of vitellogenesis, state-1 atresia (<50% α-stage atretic follicles) did not affect GSI significantly (comparisons: stages 5–6, stages 9–12, stages 14–15, but also stages 7–8 and 10–11). However, in the migratory nucleus stage, state-1 atresia significantly reduced GSI (i.e. stage 13 had a lower mean GSI compared to stage 16).
-
3.
Atretic state-2 females (stage 4: 50–100% α-atretic follicles) had significantly lower GSI than females with vitellogenic ovaries at any other histological stage (stages 5–16).
-
4.
Atretic state-3 females (stage 3: non-vitellogenic oocytes—β-atretic follicles) had GSI values similar to the yolk-vesicle stage females (stage 2).
-
5.
The presence of postovulatory follicles (indicative of recent spawning) significantly increased GSI in the primary yolk globule stage (comparisons of stages 5–8 and stages 6–7), which was not the case for the secondary yolk globule stage (stages 10–12 and stages 9–11).
GLM analysis
The analysis of GSI variation in relation to histological stage, fish length, geographic region, month of capture, fat stage and HSI indicated that the most important predictor variable was histological stage (Table 6). Actually, it explained >80% of the global GSI variation. The final model fitted also included the month and HSI variables. These two variables were significant at the 0.05 level, but explained <4% and 1% of total variation, respectively. A subsequent further examination of the data set showed that the effect of month was solely due to variation in GSI of histological stage 1 (primary oocyte stage—immature) outside the spawning period. Specifically, GSI of primary females showed a progressive decrease from a mean value of about 0.6 in May to a value of about 0.3 in July–August and a subsequent increase to 0.6 in November (analysis not shown).
The effect of each significant predictor on GSI is shown on the y-axis for the different predictor levels (x-axis) (Fig. 3). Negative values on the y-axis indicate that, at the corresponding levels, the model predicts GSI that is lower than the mean estimated by the model. The opposite holds for positive values. In that sense, histological stages higher than 7–8 had a positive effect on GSI (i.e. from about the secondary yolk globule stage onwards). Similarly, the months of November–March (months 1–5) had a positive effect on GSI. November–March were the main spawning months for sardine in the studied seas (Ganias 2003).

Sardina pilchardus. GLM-derived significant effects on sardine GSI. Each plot represents the contribution of the corresponding variable to the fitted predictor. The fitted values were adjusted to average zero, and the broken lines indicate two standard errors. Bar widths correspond to the observation frequency at each variable level (months 1–12 November–October)
Discussion
The traditional GSI model [GSI=(Wg/W)×100] has been widely used in fisheries science and experimental reproductive studies, but its properties have generally been poorly evaluated. In a first attempt to evaluate the underlying assumptions of the index, DeVlamming et al. (1982) considered that the use of GSI assumes a linear functional relationship of gonad weight–on–fish weight, with zero intercept and normally distributed errors. These assumptions contradict the principles of allometric theory (Gould 1966), which, for a pair of body parts, predict a nonlinear relationship with log-normally distributed error. The relative gonadal index (RGI=Wg/Wb, Erickson et al. 1985b) is thus the generally applicable model, with expectation (E) that is merely a function of the variance of the allometric equation (see “Materials and methods” section). The traditional GSI model is valid when b=1, i.e. ovarian growth is isometric.
Morphometric ratios may present difficulties (e.g. size dependence, unusual and possibly intractable distributions); this is particularly pronounced in situations when the denominator of the ratio is highly variable (Bolger and Connolly 1989; and references therein). DeMartini and Lau (1999) considered that variations in somatic weight related to energy allocation may be independent of ovary weight and, hence, complicate the use of gonadal indices. However, somatic weight varies much less than ovarian weight. For example, the coefficients of variation of log-transformed somatic weight and ovarian weight for Mediterranean sardine were about 12% and 113%, respectively.
More importantly, gonadal indices assume that the allometric pattern is independent of gonadal cycling, i.e. the allometric coefficient b does not change with stage of ovarian development and state of follicular atresia. In the Mediterranean sardine, ovarian growth was isometric in all histological stages, except in the stage of oocyte hydration, when a positive allometry was established. Thus, the use of GSI was valid for all but the hydrated females. It is important to emphasize that neither atresia nor recent spawning (incidence of postovulatory follicles) affected this pattern of isometric ovarian growth in the Mediterranean sardine.
Investigations of ovarian allometry in relation to stage of oocyte development have generally been few in number and variable in terms of sample sizes, assessment of ovarian development and number of developmental stages used. All species studied so far had either isometric (Taylor et al. 1998) or allometric (DeVlamming et al. 1982; Erickson et al. 1985a, 1985b; Davis and West 1993; Shapiro et al. 1993; DeMartini and Lau 1999; Kwok and Ni 1999) gonadal growth, with homogeneous coefficients of allometry throughout ovarian development. Minor exceptions are possibly due to inadequate sample size [i.e. Trichiurus nanhaiensis (Kwok and Ni 1999), see also Erickson et al. (1985a)]. The present study is, to our knowledge, the first report of an alteration in the pattern of gonadal allometry (from isometry to positive allometry) at a specific stage of oocyte maturation. Positive allometry of hydrated ovaries could be explained in terms of the intrusion of low specific-weight fluid in mature oocytes (Wallace and Selman 1981; Thorsen and Fyhn 1996) and corresponding increase in the wet weight/dry weight ratio of the ovaries. The addition of dry mass seemed to stop at the end of vitellogenesis; hence, no detectable GSI difference was found between the tertiary yolk globule stage (completion of vitellogenesis) and the migratory nucleus stage (first stage of final maturation). In each consecutive stage of the primary and secondary growth phase (sensu West 1990), i.e. up to the tertiary yolk globule stage, the increase in ovarian weight (and thus GSI) was significant (Fig. 2).
State-1 atresia did not exert any significant effect on GSI except for the final stage of maturation (migratory nucleus stage). In contrast, females in atretic state-2 had a lower GSI than females in state-0 and state-1. We therefore conclude that for follicular atresia to significantly affect GSI, its intensity must be high, which is generally indicative of the cessation of spawning (Hunter and Macewitz 1985).
The presence of postovulatory follicles did not affect GSI, except in the primary yolk globule stage. During the peak spawning months, the latter stage was mainly found in ovaries having numerous, large-sized, early POFs (Ganias et al. 2004). Females at the primary yolk globule stage with no POFs were mainly collected at the beginning of the spawning period (October). These fish might not yet have started to spawn, and their recruiting ovaries might have had lower yolk deposits than already active fish at the same stage of oocyte development.
In contrast to most species with indeterminate fecundity, the Mediterranean sardine exhibits a group-synchronous pattern of oocyte development (Ganias et al. 2004). In another group-synchronous spawner with indeterminate fecundity, the common snook (Centropomus undecimalis), ovarian growth was also isometric (Taylor et al. 1998). This contrasts to several other species, clearly shown or well known to exhibit the asynchronous pattern of oocyte development, in which the ovary grows allometrically, with coefficient b significantly different from unity [lutjanids (Davis and West 1993; DeMartini and Lau 1999), serranids (Shapiro et al. 1993; authors’ unpublished data), branchiostegids (Erickson et al. 1985a), engraulids (Osorio and Acero 1996; authors’ unpublished data)]. We believe that this contrast might provide the first evidence that a relationship exists between the pattern of oocyte development (clutch recruitment) and the pattern of ovarian allometry, i.e. isometric versus allometric ovarian growth could reflect group-synchronous versus asynchronous oocyte development.
Alternatively, the isometric versus allometric pattern of ovarian growth might be related to a respective absence or presence of size-dependent effects on fecundity and/or egg size. According to Jons and Miranda (1997), fecundity and oocyte size (weight) are the main factors affecting ovarian weight. In the Mediterranean sardine, oocyte diameter at the completion of vitellogenesis (tertiary yolk globule and migration nucleus stages) does not change significantly with fish size (Ganias 2003). Furthermore, the relative batch fecundity [number of eggs per gram body weight (Hunter et al. 1985)] does not present any fish-size dependency (Ganias et al. 2004). The absence of such size-dependent effects could also explain the isometric pattern of ovarian growth in the Mediterranean sardine.
Summarizing, the histological stages that were shown to significantly affect GSI in non-hydrated sardine females were: (1) all five stages of the primary and secondary growth phase; (2) the presence of POFs at the primary yolk globule stage; (3) atresia-1 at the migratory nucleus stage; (4) atresia-2; and (5) atresia-3. As subsequently shown by the GLM analysis, histological stage accounted for >80% of GSI variation. Other parameters did not enter the models, except for month, which explained an additional ~3% of variation, and HSI, which explained <1%. The former small amount of variation (the month effect) concerned the immature stage during the non-reproductive period (see “Results” section). The slight (however significant) effect of HSI is attributable to a positive effect of high hepatic reserves on sardine batch fecundity as shown in Ganias (2003). The finding that fish length, geographical region, month of capture, fat stage and HSI did not generally exert important effects on GSI, further strengthen the suitability of GSI for assessing ovarian activity in the Mediterranean sardine.
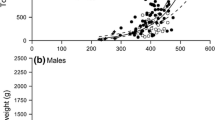
Remaining variation in GSI, not explained by histological stage, can be attributed to fecundity, which is known to independently affect ovarian weight (Jons and Miranda 1997). In general, mean batch fecundity estimates for the Mediterranean sardine show low variability with regard to season/region/year (Ganias 2003; Ganias et al. 2004). However, residual variation of the fecundity–on–somatic weight relationship is usually very large, as in most other fish species. We believe, and will provide some evidence, that fecundity variation (individual, regional, inter-seasonal, inter-annual) does not generally alter the ovarian weight–on–fish weight allometric coefficient. It is most likely related to the variance about the allometric regression. This is shown by a study on European anchovy (Engraulis encrasicolus) in the same region (Aegean and Ionian Seas). In a Daily Egg Production Method survey (Somarakis et al. 2002), anchovy showed a remarkable difference in batch fecundity between Aegean and Ionian stocks (Fig. 4a). At the same time, difference in ovarian allometry was not significant (Fig. 4b) and ovarian egg density showed no correlation with GSI (Fig. 4c) or RGI. The interplay between fecundity and spawning frequency seemed to be important, since the spawning frequency estimates showed an opposite trend (Aegean Sea: 0.15; Ionian Sea: 0.06) and resulting values for daily specific fecundity were almost equal for the two seas (about 18 eggs g−1 of reproducing stock) (Somarakis et al. 2002). Further evidence that gonadal allometry is unaffected by changes in fecundity has been provided by DeMartini (1991) for Seriphus politus. Batch fecundity was lower for this species during a major El Niño year (1984), compared with preceding (1979/1980) or following (1985/1986) pairs of years. However, the respective ovarian weight–on–fish weight relationships had homogenous allometric coefficients. The resulting gonadal condition (mean RGI) was significantly lower in 1984, indicating a lower intercept for the allometric equation during the El Niño year.

Engraulis encrasicolus. June 1999. a Batch fecundity-on-ovary free weight relationships (heterogeneous slopes, P<0.001). b Ovary weight-on-somatic weight allometric relationships for hydrated females (homogeneous slopes, P>0.05). c Egg density (batch fecundity/ovary weight) versus GSI (non-significant relationship, P>0.05). All weights in grams [filled circles, continuous lines Aegean Sea stock; open circles, broken lines Ionian Sea stock; data from Somarakis et al. (2002)]
The present paper is a case study for a single species and does not claim greater generality than it, in fact, has. The hypotheses formulated here, namely, that the pattern of ovarian allometry may not be altered by fecundity variations and that the isometric versus allometric growth pattern may reflect group-synchronous versus asynchronous oocyte development, or, alternatively, the respective absence or presence of size dependency in fecundity and/or egg size were based on carefully gathered, yet very limited, available information. We believe that future tests of these hypotheses will likely provide new insights into the reproductive biology of fishes.
References
Bolger T, Connolly PL (1989) The selection of suitable indices for the measurement and analysis of fish condition. J Fish Biol 34:171–182
Chambers JM, Hastie TJ (1992) Statistical models in S. Chapman and Hall, London
Clark C (1981) Staining procedures. Williams and Wilking, Baltimore
Coward K, Bromage N (1999) Spawning frequency, egg size and ovarian histology in groups of Tilapia zillii maintained upon two distinct food ration sizes from first-feeding to sexual maturity. Aquat Living Resour 12:11–22
Davis TLO, West GJ (1993) Maturation, reproductive seasonality, fecundity, and spawning frequency in Lutjanus vittus (Quoy and Gaimard) from the north west shelf of Australia. Fish Bull (Wash DC) 91:224–236
DeMartini EE (1991) Annual variations in fecundity, egg size, and the gonadal and somatic conditions of queenfish Seriphus politus (Sciaenidae). Fish Bull (Wash DC) 89:9–18
DeMartini EE, Lau BB (1999) Morphometric criteria for estimating sexual maturity in two snappers, Etelis carbunculus and Pristipomoides sieboldii. Fish Bull (Wash DC) 97:449–458
DeVlamming V, Grossman G, Chapman F (1982) On the use of the gonosomatic index. Comp Biochem Physiol A 73:31–39
Erickson DL, Harris MJ, Grossman GD (1985a) Ovarian cycling of tilefish, Lopholatilus chamaeleonticeps Goode and Bean, from the South Atlantic Bight, USA. J Fish Biol 27:131–146
Erickson DL, Hightower JE, Grossman GD (1985b) The relative gonadal index: an alternative index for quantification of reproductive condition. Comp Biochem Physiol A 81:117–120
Ganias K (2003) Oceanographic and biological study of sardine Sardina pilchardus (Walb., 1792) ichthyoplankton production in coastal waters of central Greece. PhD thesis, University of Thessaly, Magnesia
Ganias K, Somarakis S, Machias A, Theodorou A (2003) Evaluation of spawning frequency in a Mediterranean sardine population. Mar Biol 142:1169–1179
Ganias K, Somarakis S, Machias A, Theodorou A (2004) Pattern of oocyte development and batch fecundity in the Mediterranean sardine. Fish Res (Amst) 67:13–23
Garcia-Berthou E, Moreno-Amich R (1993) Multivariate analysis of covariance in morphometric studies of the reproductive cycle. Can J Fish Aquat Sci 50:1394–1399
Gould SL (1966) Allometry and size in ontogeny and phylogeny. Biol Rev 41:587–640
Gutreuter S (1990) Comments. Properties of relative weight and other condition indices. Trans Am Fish Soc 119:1049–1051
Hunter JR, Macewitz B (1985) Measurement of spawning frequency in multiple spawning fishes. In: Lasker R (ed) An egg production method for estimating spawning biomass of pelagic fish: application to the northern anchovy, Engraulis mordax. NOAA (Natl Ocean Atmos Adm) Tech Rep NMFS (Natl Mar Fish Serv) 36:79–93
Hunter JR, Lo NCH, Leong HJ (1985) Batch fecundity in multiple spawning fishes. In: Lasker R (ed) An egg production method for estimating spawning biomass of pelagic fish: application to the northern anchovy, Engraulis mordax. NOAA (Natl Ocean Atmos Adm) Tech Rep NMFS (Natl Mar Fish Serv) 36:67–77
Jons GD, Miranda LE (1997) Ovarian weight as an index of fecundity, maturity, and spawning periodicity. J Fish Biol 50:150–156
Kwok KY, Ni I-H (1999) Reproduction of cutlassfishes Trichiurus spp. from the South China Sea. Mar Ecol Prog Ser 176:39–47
Marshall CT, Yaragina N, Lambert Y, Kjesbu O (1999) Total lipid energy as a proxy for total egg production by fish stocks. Nature 402:288–290
McCullagh P, Nelder JA (1983) Generalized linear models. Chapman and Hall, London
Milton DA, Blaber SJM, Rawlinson NJF (1994) Reproductive biology and egg production of three species of Clupeidae from Kiribati, tropical central Pacific. Fish Bull (Wash DC) 92:102–121
Murayama T, Shirashi M, Aoki I (1994) Changes in ovarian development and plasma levels of sex hormones in the wild female Japanese sardine (Sardinops melanostictus) during the spawning period. J Fish Biol 45:235–245
Nikolsky GV (1963) The ecology of fishes. Academic, London
Osorio AC, Acero A (1996) Reproduction of Anchovia clupeoides and Anchoa parva (Pisces: Engraulidae) in two Caribbean swamps of Colombia. Rev Biol Trop 44:781–793
Shapiro DY, Savody Y, McGehee MA (1993) Periodicity of sex change and reproduction in the red hind, Epinephelus guttatus, a protogynous grouper. Bull Mar Sci 53:1151–1162
Somarakis S (2004) Marked interannual differences in reproductive parameters and daily egg production of anchovy (Engraulis encrasicolus) in the northern Aegean Sea. Belg J Zool[Suppl] (in press)
Somarakis S, Koutsikopoulos C, Machias A, Tsimenides N (2002) Applying the daily egg production method (DEPM) to small stocks in highly heterogeneous seas. Fish Res (Amst) 55:193–204
Taylor RG, Grier HJ, Whittington JA (1998) Spawning rhythms of common snook in Florida. J Fish Biol 53:502–520
Thorsen A, Fyhn H (1996) Final oocyte maturation in vivo and in vitro in marine fishes with pelagic eggs; yolk protein hydrolysis and free amino acid content. J Fish Biol 48:1195–1209
Venables WN, Ripley BD (1997) Modern applied statistics with S-PLUS, 2nd edn. Springer, New York Heidelberg Berlin
Wallace AR, Selman K (1981) Cellular and dynamic aspects of oocyte growth in teleosts. Am Zool 21:325–343
West G (1990) Methods of assessing ovarian development in fishes: a review. Aust J Mar Freshw Res 41:199–222
Acknowledgements
This study was partially funded by an EU-DG XIV study project (contract no. 98/039). Thanks are due to the coordinator of the project, Dr. A. Machias. The experimental work complied with the current laws of Greece.
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by O. Kinne, Oldendorf/Luhe
Rights and permissions
About this article
Cite this article
Somarakis, S., Ganias, K., Tserpes, G. et al. Ovarian allometry and the use of the gonosomatic index: a case study in the Mediterranean sardine, Sardina pilchardus. Marine Biology 146, 181–189 (2004). https://doi.org/10.1007/s00227-004-1419-2
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00227-004-1419-2