
Abstract
Studying individuals with specialized training, such as dancers and musicians, provides an opportunity to investigate how intensive practice of sensorimotor skills affects behavioural performance across various domains. While several studies have found that musicians have improved motor, perceptual and sensorimotor integration skills compared to untrained controls, fewer studies have examined the effect of dance training on such skills. Moreover, no study has specifically compared the effects of dance versus music training on perceptual or sensorimotor performance. To this aim, in the present study, expert dancers, expert musicians and untrained controls were tested on a range of perceptual and sensorimotor tasks designed to discriminate performance profiles across groups. Dancers performed better than musicians and controls on a dance imitation task (involving whole-body movement), but musicians performed better than dancers and controls on a musical melody discrimination task as well as on a rhythm synchronization task (involving finger tapping). These results indicate that long-term intensive dance and music training are associated with distinct enhancements in sensorimotor skills. This novel work advances knowledge of the effects of long-term dance versus music training and has potential applications in therapies for motor disorders.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Dance and music involve concentrated training of specific perceptual and sensorimotor skills that are typically initiated at a young age, are long term and can be quantified. As such, expert dancers and musicians are an ideal population in which to investigate the behavioural effects of intensive sensorimotor training. The behavioural correlates of music training have been well studied (for a review, see Herholz and Zatorre 2012), and there has been growing interest to study dance as well (for a review, see Bläsing et al. 2012 or Karpati et al. 2015). Both dance and music training have similarities, such as the importance of sensorimotor integration, as well as their artistic components and the easily quantifiable nature of the training process. However, music training generally focuses on producing sound using effector-specific movements, while dance training commonly focuses on following sound using whole-body movements. To date, no studies have directly compared the behavioural correlates of dance and music training. Comparing both forms of training is important to better understand how intensive sensorimotor training affects performance across a variety of tasks, and what effects might be specific to each form of training. Such knowledge may then be used to guide future research in clinical practice such as the development of dance- and music-based therapies for a variety of conditions such as Parkinson’s disease (Mandelbaum and Lo 2014; Duncan and Earhart 2012) and stroke (Schneider et al. 2007). To these aims, in the present study, the behavioural correlates of long-term dance and music training were compared.
Behavioural differences between dancers and non-dancers as well as between musicians and non-musicians have been found using tasks in a variety of domains including motor control, perception and sensorimotor integration. In terms of motor control, both dancers and musicians have shown enhanced performance relative to their untrained counterparts. Dancers have improved balance (Crotts et al. 1996) and postural control (Rein et al. 2011) compared to non-dancers. Musicians show advantages in hand and finger movements compared to non-musicians (Fernandes and de Barros 2012; Verheul and Geuze 2004). These results may be related to the finding that both dancers (Thullier and Moufti 2004) and musicians (Inui and Ichihara 2001) can better optimize and combine movements than untrained controls.
In addition to motor tasks, the correlates of long-term dance and music training have been examined using perceptual tasks. Perceptual studies in dancers have focused on the visual domain using dance-related stimuli. Relative to non-dancers, dancers can better detect differences in moving point-light displays (Calvo-Merino et al. 2010) and show faster eye movements when viewing a dance film (Stevens et al. 2010). In contrast, perceptual studies in musicians have focused on the auditory domain (for a review, see Herholz and Zatorre 2012); especially relevant to the present study are findings that musicians can better detect differences in rhythmic and melodic stimuli than non-musicians (Foster and Zatorre 2010; Fujioka et al. 2004; Rammsayer and Altenmuller 2006).
Sensorimotor integration is a crucial aspect of both dance and music training. Dancers combine auditory information from music, along with visual information from observing their own and others’ movements, with their motor output during performance. Musicians combine visual information from reading music or following a conductor, as well as auditory information from the output of their own and others’ instruments, with the movements required to produce the desired sound. The forms of sensorimotor integration most relevant to dance and music include audiomotor and visuomotor integration. In the audiomotor domain, dancers more accurately synchronize knee bending to an auditory beat than non-dancers (Miura et al. 2011, 2013), and musicians more accurately synchronize hand/finger tapping to an auditory stimulus than non-musicians (Repp 2010; Chen et al. 2008; Bailey et al. 2014). In the visuomotor domain, dancers show increased ability compared to non-dancers in synchronizing with observed dance movements in the absence of auditory cues (Washburn et al. 2014). Musicians have better visuomotor integration compared to non-musicians, for example, in terms of motor reproduction of visually presented temporal intervals (Aagten-Murphy et al. 2014).
Some studies in dancers and musicians have also examined performance on multi-modal tasks across the auditory, visual and motor domains. These studies are especially informative since dance performance and music performance involve the integration of all three of these domains. Sofianidis et al. (2012) observed that dancers were more stable and coordinated than non-dancers in a dance synchronization task with visual and auditory cues. In musicians, audio-visuomotor performance has been mainly studied using interference paradigms (Drost et al. 2005; Keller and Koch 2008; Taylor and Witt 2015), such as playing chords in response to visual cues while hearing irrelevant auditory stimuli (Drost et al. 2005). Musicians show a greater interference effect of incongruent auditory and visual information relative to non-musicians, signalling that this action-effect representation in multi-modal integration is affected by training (Drost et al. 2005).
Taken together, this literature suggests that both dancers and musicians show enhancements in a range of sensorimotor skills compared to their untrained counterparts. However, it is not yet known whether the sensorimotor enhancements associated with long-term dance training are similar to or distinct from those associated with long-term music training. To this aim, the main objective of the present study was to investigate the behavioural correlates of long-term dance versus music training by examining sensorimotor integration in expert dancers, expert musicians and untrained controls.
In the present study, all participants were tested on a battery of behavioural tasks ranging from more dance-relevant to more music-relevant tasks. This type of behavioural battery provides the opportunity to investigate performance that is directly relevant to the trained skill (e.g. dancers performing a dance-related task), as well as transfer effects to the other skill (e.g. dancers performing a music-related task). A dance imitation task was the most dance relevant, in which participants executed whole-body dance movements simultaneously with visual and auditory stimuli. The synchronization of whole-body movements to sensory stimuli is a main component of dance training, while music training generally relies less on whole-body movements and more on effector-specific movements. Compared to dance-related tasks used in previous studies (Cross et al. 2009; Tachibana et al. 2011; Ono et al. 2014), this task includes more ecologically valid dance movements since they require use of the whole-body rather than only the lower limbs. A rhythm synchronization task, in which participants tapped a finger in synchrony with auditory stimuli, was relevant to both dance and music since rhythm is a crucial aspect of both types of training. Dancers often synchronize their movements with the rhythmic aspect of auditory and visual stimuli, while musicians produce sound in accordance with perceived rhythmic stimuli. A melody discrimination task was the most music-relevant task, which required participants to determine whether pairs of melodies were the same or different. Musicians are often trained to distinguish between pitches and melodies in order to be able to analyse performed or heard music in a critical manner, while this is less relevant to dance training as dancers generally follow rather than produce sound. In addition, cognitive tasks (Digit Span, Letter-Number Sequencing and syllable sequence discrimination) were administered in order to test for group differences in auditory working memory (Wechsler 1997; Foster and Zatorre 2010).
On the dance imitation task, dancers were expected to outperform both musicians and controls since dance training focuses on performing whole-body movements often in synchrony with auditory and visual stimuli. This is supported by the finding that dancers perform better than non-dancers on a visuomotor dance imitation task (Washburn et al. 2014). On the rhythm synchronization task, both musicians and dancers were expected to outperform controls. Musicians have previously shown enhanced performance relative to non-musicians on the same rhythm synchronization task used here (Chen et al. 2008; Bailey et al. 2014), while dancers have shown better audiomotor integration in a rhythmic context than non-dancers (Miura et al. 2011, 2013). Since both dancers and musicians have demonstrated enhanced rhythmic abilities versus untrained controls, it was expected that they would perform similarly on the rhythm synchronization task. On the melody discrimination task, musicians were expected to outperform both dancers and controls. Music training often involves learning to distinguish between melodies, such as identifying whether a produced sound was correct or incorrect, and has been previously shown to provide advantages relative to non-musicians on the same melody task used here (Foster and Zatorre 2010).
Methods
Participants
Three groups of participants (aged 18–40 years) were recruited for this study: expert dancers (N = 20), expert musicians (N = 19) and a control group of non-musicians/non-dancers (N = 20) (Table 1). Dancers and musicians were either currently practicing as professionals or students involved in professional training programs. Their training was assessed via a detailed questionnaire developed in our laboratories (Coffey et al. 2011; Bailey and Penhune 2010). Dancers and musicians had on average approximately 15 years of experience in their respective disciplines, and controls had on average <1 year of experience in dance, music, figure skating and aerobics. All participants were physically active (i.e. biking, running or practicing other physical exercises). Dancers were currently practicing contemporary dance as their principal style, but had a variety of training backgrounds including ballet, tap, jazz, swing and ballroom. Dancers whose main style was too similar to the dance task used here (i.e. urban, street or hip hop) were excluded. Musicians had various instrumental backgrounds, including keyboard instruments, strings, woodwinds, brass and percussion. None of the musicians had absolute pitch. Since the dance task was based on a video game, participants were screened for experience with dance video games; 56 out of 59 participants reported that they never or rarely (up to three times per year) played dance video games. The remaining three participants (one dancer and two musicians) reported a maximum 4 months of experience with dance video games. The groups did not differ in age, sex distribution, body mass index (BMI) or level of education (Table 1). Participants had no past or current learning or developmental disorder, neurological or psychiatric condition, or alcohol or substance abuse. This study was approved by the Research Ethics Board at the Montreal Neurological Institute and Hospital, and written informed consent was obtained from all participants.
Tasks
Dance imitation
The whole-body dance imitation task (Fig. 1) consisted of performing sequences of dance movements from a video game, Dance Central for Xbox Kinect version 1 (Harmonix, http://www.harmonixmusic.com). Participants viewed an avatar performing dance movements to music on a large television screen directly in front of them, and were asked to mirror the avatar’s movements in real time. A rectangular area corresponding to the field of view of the Kinect sensor was marked on the floor, and participants were instructed to keep their movements within this area. Music was presented through speakers on each side of the television. Movements were recorded by the Kinect infrared light sensor (http://www.xbox.com/en-ca/Kinect; US Patent No. 20100197399), which has been shown to validly represent movement and posture (Clark et al. 2012, 2013).

Dance imitation task. The participants stand facing the television screen where they view an avatar dancing using whole-body movements, and are asked to mirror the avatar’s movements in real time. Participants’ movements are registered by the Kinect sensor
Prior to performing the test sequences, participants were required to achieve at least 60 % correct moves on a familiarization dance sequence in order to be sure that they could perform the task. One control participant did not reach this threshold and was thus excluded from the sample.
Participants performed seven test dance sequences covering a range of difficulty levels (labelled as sequences 1–7, with 1 being the easiest and 7 being the most difficult). These dance sequences were chosen from the game through pilot testing in order to minimize floor effects in the non-dancers and ceiling effects in the dancers. Level of difficulty was rated within the game and depended on the number of different moves, rate of change and complexity (e.g. speed and number of body parts required). The dance task was divided into two blocks: (1) the familiarization sequence followed by sequences 1–4 and (2) a warm-up sequence (repeat of sequence 3) followed by sequences 5–7. Familiarization and warm-up sequences were not included in the analyses. The Kinect system provided a score of the per cent of correct moves made by each participant for each sequence. This score was based on the movement of the correct body parts in the correct direction within a restricted time window. The manufacturer does not disclose additional details about how scores are calculated.
Rhythm synchronization
In the rhythm synchronization task (Fig. 2), as used by Bailey and Penhune (2010), participants were asked to listen to and tap in synchrony with auditory rhythms at three levels of metrical complexity. Each trial consisted of two presentations of the same stimulus. Participants were instructed to listen to the first presentation and tap in synchrony with the second on a computer mouse button. Participants completed two blocks of 36 trials each. Stimuli were 6 s in duration and included 11 woodblock sounds of 200 ms each. Each stimulus included five onset-to-onset intervals of 250 ms, three 500-ms intervals, and one interval of each of 750, 1000 and 1500 ms. The order of these intervals differed in each stimulus to create three levels of increasing rhythmic complexity corresponding to decreasing metrical structure: metric simple (MS), metric complex (MC) and non-metric (NM). Two rhythms of each complexity were used. The six rhythms were randomly ordered, and each was presented six times per block. The task was scored using the absolute value inter-tap interval (ITI) deviation, which is the ratio of the time between two taps (ITI) to the time between the two corresponding sounds in the stimulus (inter-stimulus interval) and measures the accuracy in reproducing the temporal intervals between sounds in the stimulus. Lower ITI values indicate higher interval reproduction accuracy and therefore better performance. For more details about the stimuli and scoring of this task, please see Bailey and Penhune (2010).

Examples of stimuli and scoring of the rhythm synchronization task. Participants listen to and tap in synchrony with stimuli of three rhythmic complexities. Inter-tap interval (ITI) deviation is calculated as the ratio of the ITI to the corresponding inter-stimulus interval (figure adapted from Bailey and Penhune 2010)
Melody and syllable sequence discrimination
The melody discrimination task (Fig. 3), designed by Foster and Zatorre (2010), required participants to determine whether two melodic stimuli were the same or different. It consisted of two sub-tasks, simple and transposed melody discrimination, each consisting of two blocks of 30 trials each. The simple task was always performed before the transposed task. In the simple task, two melodies containing 5–13 low-pass-filtered harmonic tones were presented. Tones were 320 ms in duration and included notes in the Western major scale ranging in pitch from C4 to E6. In half the trials, these melodies were identical (“same” trials). In the other half, one tone in the second melody was altered in pitch by up to five semitones (“different” trials). Key and melodic contour were not affected by this alteration. Following presentation of each melody pair, participants were asked whether the presented melodies were the same or different. The transposed task was similar to the simple task, except that all second melodies were transposed four semitones higher in pitch than the first melodies and the pitch alterations in the “different” trials had a magnitude of one semitone, which produced a note outside of the melody’s key. This task was scored using per cent of trials correct.

Examples of stimuli from the melody and syllable sequence discrimination tasks, in which participants hear pairs of melodies or syllable sequences and are asked to determine whether they are the same or different. Arrows show alterations in the “different” stimuli (figure adapted from Foster and Zatorre 2010)
The syllable sequence discrimination task (Fig. 3), also designed by Foster and Zatorre (2010), is a control task for the melody discrimination task described above. By comparing performance on this task to that of the melody discrimination task, it can be determined whether group differences on the melody task represent music-specific skills or may be due to group differences in working memory. On each trial, participants heard two syllable sequences and were asked to determine whether they were the same or different. Syllables consisted of one consonant with one vowel and were recorded in monotone from one speaker. Sequences had no meaning in English and contained 5–13 syllables of 320 ms each. On half the trials, the two sequences were identical. On the other half, one syllable was changed to a different syllable in the second sequence. Participants completed two blocks of 30 trials each. This task was scored using per cent of trials correct.
Cognitive tasks
To test for group differences in auditory working memory, participants completed two subtasks of the Weschler Adult Intelligence Scale (WAIS) III: Digit Span and Letter-Number Sequencing (Wechsler 1997). Scores on each task were converted to scaled scores using standard protocols.
Procedure
Each participant completed all tasks in one or two sessions. The rhythm, melody and syllable tasks were administered using Presentation software (Neurobehavioural Systems, http://www.neurobs.com), and auditory stimuli for these tasks were delivered using Sennheiser HD 25-1 II headphones at a comfortable volume. Participants were randomly assigned to one of four task orders, and the task order distribution was matched between groups. The non-dance tasks were performed between the dance task blocks to minimize the effect of fatigue on dance task performance.
Analyses
Group characteristics
Age, BMI, level of education and years of dance and music training were compared between groups (dancer, musician or control) using one-factor ANOVAs with group as the between-subjects factor. Post hoc pairwise comparisons were conducted where appropriate and corrected for multiple comparisons using the Bonferroni method.
Within-task analyses
Linear fixed-effects models with repeated measures were used to analyse each task using its respective score. This technique uses a restricted maximum likelihood estimation method, which provides identical estimates to ANOVA when applied to balanced data (IBM Corporation 2012), but has important differences: it allows the specification of the relationship between repeated measures (i.e. covariance structure) so that the sphericity assumption does not have to be satisfied, and it allows the inclusion of subjects with missing data. These models included group as a between-subjects factor and task-specific within-subject repeated measures using a compound symmetry covariance structure. Task-specific repeated measures included sequence difficulty in the dance task, rhythm complexity in the rhythm task, simple versus transposed melody discrimination subtasks, and syllable sequence versus melody discrimination tasks. Group comparisons in the cognitive tasks were analysed using univariate ANCOVAs. Post hoc pairwise comparisons were conducted where appropriate and were corrected for multiple comparisons using the Bonferroni method. To control for effects of age and sex on task performance, these variables were included as covariates of no interest in all analyses.
Between-task analyses
To allow for between-task comparisons, overall scores for the dance, rhythm and melody tasks were converted to z-scores (i.e. standardized scores). Since lower scores on the rhythm task correspond to better performance, z-scores for this task were multiplied by −1 so that higher z-scores corresponded to higher performance, which was consistent with the directionality of the other task scores. A linear fixed-effects model was conducted on these data, with group as a between-subjects fixed factor and task as a within-subjects repeated measure using an unstructured covariance matrix. To control for effects of age and sex on task performance, these variables were included as covariates of no interest.
Results
Group characteristics
One-factor ANOVAs conducted on age, BMI and level of education with group (dancer, musician or control) as the between-subjects factor revealed no significant differences between groups [BMI: F(2,55) = 0.4, p = 0.68; age: F(2,56) = 1, p = 0.28; education: F(2,56) = 0.6, p = 0.57]. One-factor ANOVAs conducted on years of dance and music training with group as the between-subjects factor revealed significant differences for both training types [music: F(2,55) = 271 p < 0.0001; dance: F(2,56) = 135 p < 0.0001]. Post hoc pairwise comparisons confirmed that musicians and controls did not differ in years of dance training (p = 1) and dancers and controls did not differ in years of music training (p = 0.27).
Dance imitation
Linear fixed-effects analysis using group as a between-subjects factor and sequence difficulty as a within-subjects repeated measure showed significant main effects of group [F(2,54) = 85, p < 0.0001] and sequence difficulty [F(6,334) = 154, p < 0.0001] as well as an interaction between group and difficulty [F(12,334) = 6, p < 0.0001]. Post hoc pairwise comparisons suggest that overall, dancers had the highest per cent moves correct, followed by musicians, and controls had the lowest per cent moves correct (p ≤ 0.003). Decomposition of the interaction suggests that on the easiest and three most difficult sequences, dancers performed better than both musicians and controls (p < 0.0001) with no difference between musicians and controls (p > 0.18). On the remaining three intermediate sequences, dancers performed better than both other groups, and musicians performed better than controls (p ≤ 0.01).
Rhythm synchronization
Linear fixed-effects analysis using group as a between-subjects factor and stimulus complexity (MS, MC or NM) as a within-subjects repeated measure revealed significant main effects of group [F(2,54) = 13, p < 0.0001] and stimulus complexity [F(2,112) = 42, p < 0.0001], with no significant group by complexity interaction [F(4,112) = 0.4, p > 0.8]. Musicians showed lower ITI deviation (better performance) than both dancers and controls (p ≤ 0.007), and no difference was observed between dancers and controls (p = 0.19). Across groups, the lowest ITI deviations were observed for MS rhythms, followed by MC rhythms, and highest ITI deviations were observed in the NM condition (p < 0.0001). In six participants (4 dancers, 1 musician, 1 control), one block of this task was discarded due to at least ten instances where the participant tapped during the listening stimulus instead of the tapping stimulus. Since there was an overall learning effect between the two blocks on this task (paired t test comparing block 1 and block 2: t = 6.28, p < 0.0001), the scores of these six participants were calculated using their single block score and an adjustment to compensate for the learning effect (estimated by regression analysis of the remaining participants).
Melody and syllable sequence discrimination
Linear fixed-effects analysis using group as a between-subjects factor and melody versus syllable discrimination tasks as a within-subjects repeated measure revealed significant main effects of group [F(2,54) = 9, p < 0.0001] and task [F(1,56) = 7, p = 0.014], as well as a significant interaction between group and task [F(2,56) = 29, p < 0.0001]. Post hoc pairwise comparisons on the interaction showed that musicians performed better than both other groups (p < 0.0001) with no difference between dancers and controls (p = 1) in the melody task, but no group differences were found in the syllable task (p > 0.7).
Linear fixed-effects analysis using group as a between-subjects factor and simple versus transposed melody discrimination tasks as a within-subjects repeated measure showed significant main effects of group [F(2,54) = 28, p < 0.0001] and task [F(1,56) = 38, p < 0.0001], and a significant interaction between group and task [F(2,56) = 4, p = 0.017]. Decomposition of the interaction revealed that musicians performed better than both dancers and controls (p < 0.0001), with no difference between dancers and controls (p ≥ 0.14), on both simple and transposed tasks. Both dancers and musicians performed significantly better on the simple compared to the transposed task (p ≤ 0.003), while controls did not show this difference (p = 0.095).
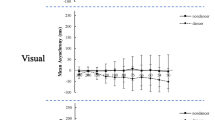
Z-score analysis (Fig. 4)
Linear fixed-effects analysis on the z-scores of the dance, rhythm and melody tasks (Fig. 4) with group as a between-subjects factor and task as a within-subjects repeated measure revealed a significant main effect of group [F(2,51) = 26, p < 0.0001] and an interaction between group and task [F(4,56) = 30, p < 0.0001]. No main effect of task was found [F(2,56) = 0.001, p = 0.99].

Performance across the three groups (dancers are shown as filled circles, musicians as unfilled squares and controls as striped triangles) as measured by z-scores (adjusted for the covariates of age and sex) are shown across tasks from the most dance-related task (dance imitation), the rhythm synchronization, to the most music-related task (melody discrimination) as well as the syllable control task. As shown, dancers perform best on the dance task, and musicians perform best on the melody task
Post hoc comparisons revealed that across tasks, dancers and musicians showed similar z-scores (p = 0.24), and both groups showed higher z-scores than controls (p < 0.0001). Decomposition of the group by task interaction showed identical group differences as found in the individual task analyses. Post hoc analyses on the task by group interaction showed that dancers had higher z-scores on the dance task than both other tasks (p < 0.0001), and scored similarly on the rhythm and melody tasks (p = 0.4). Musicians had similar z-scores on the melody and rhythm tasks (p = 0.49), both of which were higher than their z-scores on the dance task (p < 0.0001). Controls scored similarly on all three tasks (p > 0.4).
Cognitive tasks
Univariate ANCOVAs revealed no differences between groups in the Digit Span task [F(2,54) = 1, p = 0.41]. A significant effect of group was found for the Letter-Number Sequencing task [F(2,54) = 5, p = 0.013), and post hoc pairwise comparisons showed that musicians performed better than both dancers and controls (p < 0.03) and no difference was observed between dancers and controls (p = 1).
Discussion
In the present study, performance on a battery of sensorimotor tasks ranging from more dance-related to more music-related as well as cognitive tasks was compared between expert dancers, expert musicians and untrained controls. On the dance imitation task, dancers performed best, followed by musicians, whereas controls scored the lowest. On the rhythm synchronization and melody discrimination tasks, musicians performed best, while dancers and controls performed similarly. This work investigates the behavioural profiles of expert dancers and musicians and thereby increases understanding of the specificity of the effects of long-term dance versus music training.
Dance imitation
The dance-related task used in this study was a whole-body dance video game imitation task that required audio-visuomotor integration. Participants viewed an avatar dancing to music and were asked to imitate the avatar’s movement in real time. As expected, dancers performed better than both controls and musicians on the dance task. This result indicates that dancers have increased audio-visuomotor integration abilities involving whole-body movements relative to non-dancers. These findings are consistent with previous results from a study by Washburn et al. (2014) in which dancers performed better than non-dancers in a visuomotor interpersonal dance imitation task. These findings also support the work of Sofianidis et al. (2012) in which dancers were more stable and coordinated than non-dancers in a dance-related task involving the auditory, visual and motor domains. These results are likely explained by the fact that dance training involves performing whole-body movements in synchrony with auditory stimuli as well as other dancers. Dancers are trained to integrate the movements of multiple body parts (Thullier and Moufti 2004), and often learn choreography by observing and imitating movements. The advantage for dancers in this task may have been associated with an enhanced ability to engage the action observation network based on previous experience. This network of brain regions is implicated in action observation, understanding and imitation (Rizzolatti and Craighero 2004; Iacoboni 2005; Caspers et al. 2010). Consistent with these functions, dancers showed increasing engagement of this system as they learned a new dance sequence (Cross et al. 2006).
Musicians also performed better than untrained controls on the dance task, but only at the intermediate difficulty level. This indicates that music training showed some transfer to the dance imitation task. This finding supports previous work showing that musicians perform better than non-musicians in tasks involving sensorimotor integration, specifically the synchronization of finger or hand/arm movements to auditory (Chen et al. 2008; Repp 2010; Bailey et al. 2014) or visual (Spilka et al. 2010) stimuli. Even though the specific types of movements (i.e. whole-body vs. effector-specific) and stimuli may differ between music and dance contexts, the overlap may be enough to provide musicians with an advantage in dance. It is also possible that musical training allows musicians to make better temporal predictions from the music used in the dance task, allowing them to make more synchronized movements. Finally, as for the dancers, musicians may have enhanced abilities to engage the action observation network during this task which may facilitate their ability to imitate and synchronize movements with the avatar. This is supported by findings of enhancements in the mirror neuron system in musicians compared to non-musicians (Bangert et al. 2006; Pau et al. 2013; Proverbio et al. 2014).
Rhythm synchronization
The rhythm synchronization task lies between the dance imitation and melody discrimination tasks on the continuum from more dance related to more music related, since rhythm is a crucial component of both dance and music training. Participants were asked to synchronize finger taps to auditory rhythmic stimuli of varying metrical complexities. Across all participants, performance decreased with increasing metrical complexity, consistent with previous studies using the same task (Chen et al. 2008; Bailey and Penhune 2010) and other similar paradigms (Grahn and Brett 2007). As expected, musicians performed better than untrained controls on the rhythm synchronization task, consistent with previous reports of enhanced rhythm synchronization in musicians versus non-musicians on this same task (Chen et al. 2008; Bailey et al. 2014) as well as other work showing that musicians are less variable than non-musicians when tapping in synchrony with a changing auditory stimulus (Repp 2010). Musicians also performed better than dancers on the rhythm synchronization task, but dancers did not perform better than untrained controls. This may be due to the fact the rhythm task relied on finger tapping rather than whole-body movements and that finger movements are more prevalent in music than dance training.
Melody and syllable sequence discrimination
In the most music-related task, melody discrimination, participants were asked to determine whether pairs of melodies were the same or different. As expected, musicians performed better than both dancers and controls on this task. The syllable sequence discrimination task consisted of the same design except using non-musical syllable sequences instead of melodies, and was administered as a control for the melody discrimination task. No group differences were found in the syllable task. These results are consistent with those of Foster and Zatorre (2010), who found that musicians performed better than non-musicians on the same melody discrimination task used here, and both groups performed similarly on the syllable sequence discrimination task. The lack of group differences on the syllable task suggests that the differences observed in the melody task are related to music-specific advantages in the musician group and not to auditory working memory differences between groups. These advantages are likely due to the fact that melody discrimination is a crucial aspect of music training. Musicians must be able to hear pitch differences between melodies in order to determine whether they have correctly performed a musical sequence.
Overall, dancers performed similar to controls on the melody task. However, like musicians but unlike controls, they performed significantly better on the simple compared to the transposed condition of the task. This suggests that dancers’ exposure to music during training may have some effect on their ability to process melodic information.
Cognitive tasks
Cognitive tasks (Digit Span and Letter-Number Sequencing) were administered to test for group differences in auditory working memory. No performance differences were found between dancers, musicians and controls on the Digit Span task; however, musicians performed better than both dancers and controls on the Letter-Number Sequencing task. These results are consistent with previous findings in musicians that years of music training was positively correlated with performance on the Letter-Number Sequencing task, which places greater demands on auditory working memory (Bailey and Penhune 2010) relative to the other tasks used. Potential explanations include training-associated enhanced working memory ability in musicians, or that individuals who pursued music training may have pre-existing enhancements in such cognitive skills.
Overall results
This study is the first to compare the behavioural correlates of dance versus music training and compare task performance between expert dancers and musicians. It expands on work comparing each of these groups to their untrained counterparts by demonstrating that dancers and musicians have distinct behavioural profiles relative to untrained controls as well as to each other. It also builds on the literature by applying tasks across the continuum from dance relevant to music relevant in the same sample. Individual dance- or music-relevant tasks have been used independently (e.g. Washburn et al. 2014; Bailey and Penhune 2010; Foster and Zatorre 2010); however, combining them allows for the investigation of a larger range of sensorimotor abilities.
Taken together, the results of the present study suggest that dancers and musicians have distinct sensorimotor enhancements relative to untrained controls. Dancers showed enhanced sensorimotor integration involving whole-body movements compared to both musicians and controls as demonstrated by their higher scores on the dance imitation task. Dancers performed similarly to controls on the rhythm synchronization task, but performed better on the simple than transposed melody discrimination (as observed in musicians but not controls), suggesting the possibility of some effect of dance training on melodic processing. As expected, musicians showed enhancements in melody discrimination and rhythm synchronization, but they also performed better than untrained controls on the dance imitation task, suggesting that music training may show some transfer to whole-body movement.
These findings have implications for sensorimotor training in general. They suggest that learning a specific skill is associated with improvements in tasks with a strong relation to that skill and that transference to less-related tasks may occur.
Future directions
Further work on the topic of the behavioural correlates of dance and music training will continue to advance knowledge on the specificity of these skills. To build on the present study, longitudinal work comparing task performance before and after dance and music training would be especially informative as this would distinguish skill enhancement caused by training from that which may have been present before training and predisposed certain individuals to pursue training. Another important avenue for further study is the investigation of a sensitive period in the context of dance. Sensitive periods have been demonstrated for music (for a review, see Penhune 2011), but have not yet been investigated for dance. This topic could be examined by comparing task performance between groups of early- and late-trained dancers, and would provide insight into the interaction between development and learning dance-related skills such as sensorimotor integration involving whole-body movement. In addition to studying the behavioural correlates of dance versus music training, comparing the brain structural and functional correlates of these two types of training can further our understanding of their effects and provide support and explanations for the observed behavioural correlates. Our laboratory is currently investigating the grey matter (Karpati et al. 2014) and white matter (Giacosa et al. 2014) structural correlates of dance versus music training. A clinical avenue for future work is the development of dance- and music-based therapies for a variety of conditions including Parkinson’s disease (Mandelbaum and Lo 2014; Duncan and Earhart 2012) and stroke (Schneider et al. 2007). Evidence to date has demonstrated the efficacy of dance- and music-related interventions separately, and understanding how each may target specific skills may assist in designing protocols for specific symptoms or populations.
Conclusions
This study demonstrates that both dance and music training provide advantages in sensorimotor tasks, and each is associated with a unique behavioural profile. Dancers and musicians both showed a large advantage in tasks related to their trained skill, and some transference between skills was observed. This study expands on previous work that investigated the behavioural correlates of dance or music training separately, and is the first study to directly compare these two types of training. This work advances the knowledge of the specificity of dance versus music training on sensorimotor skills and can be applied to the development of arts-based therapies for motor disorders. By understanding the similar and different effects of dance versus music training, one could more accurately design a therapy protocol using one or both of these types of training to target specific skills.
References
Aagten-Murphy D, Cappagli G, Burr D (2014) Musical training generalises across modalities and reveals efficient and adaptive mechanisms for reproducing temporal intervals. Acta Psychol 147:25–33. doi:10.1016/j.actpsy.2013.10.007
Bailey JA, Penhune VB (2010) Rhythm synchronization performance and auditory working memory in early- and late-trained musicians. Exp Brain Res 204:91–101. doi:10.1007/s00221-010-2299-y
Bailey JA, Zatorre RJ, Penhune VB (2014) Early musical training is linked to gray matter structure in the ventral premotor cortex and auditory–motor rhythm synchronization performance. J Cogn Neurosci 26:755–767. doi:10.1162/jocn_a_00527
Bangert M, Peschel T, Schlaug G, Rotte M, Drescher D, Hinrichs H, Heinze HJ, Altenmüller E (2006) Shared networks for auditory and motor processing in professional pianists: evidence from fMRI conjunction. NeuroImage 30:917–926
Bläsing B, Calvo-Merino B, Cross ES, Jola C, Honisch J, Stevens CJ (2012) Neurocognitive control in dance perception and performance. Acta Psychol 139:300–308. doi:10.1016/j.actpsy.2011.12.005
Calvo-Merino B, Ehrenberg S, Leung D, Haggard P (2010) Experts see it all: configural effects in action observation. Psychol Res 74:400–406. doi:10.1007/s00426-009-0262-y
Caspers S, Zilles K, Laird AR, Eickhoff SB (2010) ALE meta-analysis of action observation and imitation in the human brain. NeuroImage 50:1148–1167. doi:10.1016/j.neuroimage.2009.12.112
Chen JL, Penhune VB, Zatorre RJ (2008) Moving on time: brain network for auditory-motor synchronization is modulated by rhythm complexity and musical training. J Cogn Neurosci 20:226–239. doi:10.1162/jocn.2008.20018
Clark RA, Pua YH, Fortin K, Ritchie C, Webster KE, Denehy L, Bryant AL (2012) Validity of the Microsoft Kinect for assessment of postural control. Gait Posture 36:372–377. doi:10.1016/j.gaitpost.2012.03.033
Clark RA, Bower KJ, Mentiplay BF, Paterson K, Pua YH (2013) Concurrent validity of the Microsoft Kinect for assessment of spatiotemporal gait variables. J Biomech 46:2722–2725. doi:10.1016/j.jbiomech.2013.08.011
Coffey EBJ, Herholz SC, Scala S, Zatorre RJ (2011) Montreal Music History Questionnaire: a tool for the assessment of music-related experience in music cognition research. In: Conference proceedings from The Neurosciences and Music IV, Edinburgh, UK
Cross ES, Hamilton AF, Grafton ST (2006) Building a motor simulation de novo: observation of dance by dancers. NeuroImage 31:1257–1267. doi:10.1016/j.neuroimage.2006.01.033
Cross ES, Kraemer DJ, Hamilton AF, Kelley WM, Grafton ST (2009) Sensitivity of the action observation network to physical and observational learning. Cereb Cortex 19:315–326. doi:10.1093/cercor/bhn083
Crotts D, Thompson B, Nahom M, Ryan S, Newton RA (1996) Balance abilities of professional dancers on select balance tests. J Orthop Sport Phys 23:12–17. doi:10.2519/jospt.1996.23.1.12
Drost UC, Rieger M, Brass M, Gunter TC, Prinz W (2005) Action-effect coupling in pianists. Psychol Res 69:233–241. doi:10.1007/s00426-004-0175-8
Duncan RP, Earhart GM (2012) Randomized controlled trial of community-based dancing to modify disease progression in Parkinson disease. Neurorehab Neural Repair 26:132–143. doi:10.1177/1545968311421614
Fernandes LF, de Barros RM (2012) Grip pattern and finger coordination differences between pianists and non-pianists. J Electromyogr Kines 22:412–418. doi:10.1016/j.jelekin.2012.02.007
Foster NE, Zatorre RJ (2010) A role for the intraparietal sulcus in transforming musical pitch information. Cereb Cortex 20:1350–1359. doi:10.1093/cercor/bhp199
Fujioka T, Trainor LJ, Ross B, Kakigi R, Pantev C (2004) Musical training enhances automatic encoding of melodic contour and interval structure. J Cogn Neurosci 16:1010–1021. doi:10.1162/0898929041502706
Giacosa C, Karpati FJ, Foster NEV, Penhune VB, Hyde KL (2014) White matter differences in dancers and musicians. In: Conference proceedings from The Neurosciences and Music V, Dijon, France
Grahn JA, Brett M (2007) Rhythm and beat perception in motor areas of the brain. J Cogn Neurosci 19:893–906. doi:10.1162/jocn.2007.19.5.893
Herholz SC, Zatorre RJ (2012) Musical training as a framework for brain plasticity: behavior, function, and structure. Neuron 76:486–502. doi:10.1016/j.neuron.2012.10.011
Iacoboni M (2005) Neural mechanisms of imitation. Curr Opin Neurobiol 15:632–637
IBM Corporation (2012) Restricted maximum likelihood method. SPSS Statistics 21.0.0. http://www-01.ibm.com/support/knowledgecenter/#!/SSLVMB_21.0.0/com.ibm.spss.statistics.cs/varcomp_reml_method.htm. Accessed 10 Aug 2015
Inui N, Ichihara T (2001) Comparison of the relation between timing and force control during finger-tapping sequences by pianists and non pianists. Mot Control 5:385–398
Karpati FJ, Giacosa C, Foster NE, Penhune VB, Hyde KL (2014) The specificity of dance versus music training on gray matter structure. In: Conference proceedings from the 20th annual meeting of the Organization for Human Brain Mapping, Hamburg, Germany
Karpati FJ, Giacosa C, Foster NE, Penhune VB, Hyde KL (2015) Dance and the brain: a review. Ann N Y Acad Sci 1337:140–146. doi:10.1111/nyas.12632
Keller PE, Koch I (2008) Action planning in sequential skills: relations to music performance. Q J Exp Psychol 61:275–291. doi:10.1080/17470210601160864
Mandelbaum R, Lo AC (2014) Examining dance as an intervention in Parkinson’s disease: a systematic review. Am J Dance Ther 36:160–175. doi:10.1007/s10465-014-9181-6
Miura A, Kudo K, Ohtsuki T, Kanehisa H (2011) Coordination modes in sensorimotor synchronization of whole-body movement: a study of street dancers and non-dancers. Hum Mov Sci 30:1260–1271. doi:10.1016/j.humov.2010.08.006
Miura A, Kudo K, Nakazawa K (2013) Action-perception coordination dynamics of whole-body rhythmic movement in stance: a comparison study of street dancers and non-dancers. Neurosci Lett 544:157–162. doi:10.1016/j.neulet.2013.04.005
Ono Y et al (2014) Frontotemporal oxyhemoglobin dynamics predict performance accuracy of dance simulation gameplay: temporal characteristics of top-down and bottom-up cortical activities. NeuroImage 85:461–470. doi:10.1016/j.neuroimage.2013.05.071
Pau S, Jahn G, Sakreida K, Domin M, Lotze M (2013) Encoding and recall of finger sequences in experienced pianists compared with musicale naïve controls: a combined behavioral and functional imaging study. NeuroImage 64:379–387
Penhune VB (2011) Sensitive periods in human development: evidence from musical training. Cortex 47:1126–1137. doi:10.1016/j.cortex.2011.05.010
Proverbio AM, Calbi M, Manfredi M, Zani A (2014) Audio-visuomotor processing in the musician’s brain: an ERP study on professional violinists and clarinetists. Sci Rep 4:5866
Rammsayer T, Altenmuller E (2006) Temporal information processing in musicians and nonmusicians. Music Percept 24:37–48. doi:10.1525/mp.2006.24.1.37
Rein S, Fabian T, Zwipp H, Rammelt S, Weindel S (2011) Postural control and functional ankle stability in professional and amateur dancers. Clin Neurophysiol 122:1602–1610. doi:10.1016/j.clinph.2011.01.004
Repp BH (2010) Sensorimotor synchronization and perception of timing: effects of music training and task experience. Hum Mov Sci 29:200–213. doi:10.1016/j.humov.2009.08.002
Rizzolatti G, Craighero L (2004) The mirror-neuron system. Annu Rev Neurosci 27:169–192
Schneider S, Schonle PW, Altenmuller E, Munte TF (2007) Using musical instruments to improve motor skill recovery following a stroke. J Neurol 254:1339–1346. doi:10.1007/s00415-006-0523-2
Sofianidis G, Hatzitaki V, McKinley P (2012) Effects of expertise and auditory guidance on traditional dance performance. J Dance Med Sci 16:57–64
Spilka MJ, Steele CJ, Penhune VB (2010) Gesture imitation in musicians and non-musicians. Exp Brain Res 204:549–558. doi:10.1007/s00221-010-2322-3
Stevens C, Winskel H, Howell C, Vidal LM, Latimer C, Milne-Home J (2010) Perceiving dance: schematic expectations guide experts’ scanning of a contemporary dance film. J Dance Med Sci 14:19–25
Tachibana A, Noah JA, Bronner S, Ono Y, Onozuka M (2011) Parietal and temporal activity during a multimodal dance video game: an fNIRS study. Neurosci Lett 503:125–130. doi:10.1016/j.neulet.2011.08.023
Taylor JE, Witt JK (2015) Listening to music primes space: pianists, but not novices, simulate heard actions. Psychol Res 79:175–182. doi:10.1007/s00426-014-0544-x
Thullier F, Moufti H (2004) Multi-joint coordination in ballet dancers. Neurosci Lett 369:80–84. doi:10.1016/j.neulet.2004.08.011
Verheul MH, Geuze RH (2004) Bimanual coordination and musical experience: the role of intrinsic dynamics and behavioral information. Mot Control 8:270–291
Washburn A, DeMarco M, de Vries S, Ariyabuddhiphongs K, Schmidt RC, Richardson MJ, Riley MA (2014) Dancers entrain more effectively than non-dancers to another actor’s movements. Front Hum Neurosci 8:800. doi:10.3389/fnhum.2014.00800
Wechsler D (1997) Wechsler adult intelligence scale, 3rd edn. Harcourt Assessment, San Antonio
Acknowledgments
We would like to thank our participants for their time, as well as Alessia Di Cesare, Veronica Yuk, and Dana Boebinger for their assistance in the data collection process. This work was funded by a grant from the Natural Sciences and Engineering Council of Canada (NSERC) to Dr. Krista Hyde.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
The authors declare that they have no conflict of interest. All procedures performed in studies involving human participants were in accordance with the ethical standards of the institutional and/or national research committee and with the 1964 Helsinki Declaration and its later amendments or comparable ethical standards. Informed consent was obtained from all individual participants included in the study. Additional informed consent was obtained from all individual participants for whom identifying information is included in this article.
Rights and permissions
About this article

Cite this article
Karpati, F.J., Giacosa, C., Foster, N.E.V. et al. Sensorimotor integration is enhanced in dancers and musicians. Exp Brain Res 234, 893–903 (2016). https://doi.org/10.1007/s00221-015-4524-1
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00221-015-4524-1