
Abstract
Loneliness is an unpleasant and distressing feeling that a person experiences when he/she perceives that his/her social relationships are lacking in someway, either quantitatively or qualitatively; this can be linked to anxiety, depression, and suicide risk. Previous studies have found that certain personality traits (which are temporally stable and heritable) are predictors of loneliness. However, little empirical evidence is available on the brain structures associated with loneliness, as well as how personality traits impact the relationship between loneliness and brain structure. Thus, the current study used voxel-based morphometry to identify the brain structures underlying individual differences in loneliness (as measured by the UCLA Loneliness Scale) in a large sample, and then, applied multiple mediation analyses to explore the nature of the influence of personality traits on the relationship between loneliness and brain structure. The results showed that lonely individuals had greater regional gray matter volume in the left dorsolateral prefrontal cortex (DLPFC), which might reflect immature functioning in terms of emotional regulation. More importantly, we found that neuroticism and extraversion partially mediated the relationship between the left DLPFC and loneliness. In summary, through morphometric and multiple mediation analyses, this paper further validates the influence of both neuroticism and extraversion on loneliness.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
Loneliness is an unpleasant and distressing feeling that a person experiences when he/she perceives his/her social relationships to be lacking in someway, either quantitatively or qualitatively (Perlman and Peplau 1981, 1982). Obviously, the definition emphasizes the affective character of loneliness. Specifically, loneliness involves many emotional aspects such as sorrow, sadness and feelings of frustration, shame, and desperation (Shute and Howitt 1990). Loneliness is a common phenomenon in contemporary societies (Putnam 2000; McPherson et al. 2006; Kanai et al. 2012). Some surveys have reported that 15–30 % of people experience persistent feelings of loneliness (Sermat and Hartog 1980; Andersson 1982). In addition, loneliness very frequently occurs during earlier developmental periods (Peplau and Perlman 1982; Perlman and Landolt 1999). For example, in a large-scale investigation, Parlee (1979) found that 79 % of subjects aged younger than 18 years and 71 % of 18- to 24-year-olds reported feeling lonely sometimes or often. Although loneliness can be a normative experience, it can also be pathological (Asher and Paquette 2003). Numerous researchers have confirmed that loneliness is linked to anxiety, depression, and suicide risk (Moore and Schultz 1983; Koenig et al. 1994; Roberts et al. 1998; Lau et al. 1999; Johnson et al. 2001; Lasgaard 2007).
Moreover, many studies have investigated the relationship between loneliness and personality traits and found that neuroticism, extraversion, and psychoticism might be the predictors of loneliness (Hojat 1982; Stokes 1985; Saklofske and Yackulic 1989; Halamandaris and Power 1999; Cheng and Furnham 2002; Atak 2009). It is well known that neuroticism and extraversion are the most important and frequently studied traits of personality and are correlated with emotional experience (Williams 1990; Kardum and Hudek-Knežević 1996; Furnham and Cheng 1999). For example, neurotic individuals are typically described with negative emotional terms such as depression and anxiety, whereas extraverted individuals are typically described with positive emotional terms such as excitement and enthusiasm (Eysenck and Eysenck 1991). At the same time, psychoticism is proposed to indicate a predisposition toward coldness, aggressiveness, egocentrism, impersonality and antisocial nature, lack of empathy, creativity, impulsivity, and tough-mindedness (Eysenck and Eysenck 1985); thus, it is a trait which encompasses relatively powerful emotional content (Zuckerman 1989). These personality traits (neuroticism, extraversion, and psychoticism) are good candidates for research because their temporal stability and heritability make them likely to have an impact on function throughout the life cycle (Loehlin and Martin 2001; Birley et al. 2006; Bratko and Butkovic 2007; Ivkovic et al. 2007; Wray et al. 2007). However, so far, little empirical evidence is available on how personality impacts the relationship between loneliness and brain structure.
As of now, there are few neuroimaging studies about the neural processes related to loneliness. For example, Eisenberger et al. (2003) indicated that the anterior cingulate cortex (ACC; related to increased distress after social exclusion) and right ventral prefrontal cortex (PFC; correlated with diminished distressing after social exclusion) were active during the social pain participants felt during an episode of social exclusion. At the same time, some neuroimaging studies have revealed that neuroticism and extraversion were also significantly correlated with the lateral PFC and ACC (Canli et al. 2001; Eisenberger et al. 2005). Kross et al. (2011) also found that the secondary somatosensory cortex and dorsal posterior insula became active when participants thought about being rejected by their ex-partner. In a recent voxel-based morphometry (VBM) study, Kanai et al. (2012) showed that those who are more prone to loneliness had less gray matter volume (GMV) in the left posterior superior temporal sulcus (pSTS), which may reflect deficiencies in processing social cues (social perception plays a crucial role in shaping an individual’s experience of loneliness). Moreover, they also found that social network size, anxiety, and empathy independently contributed to loneliness. However, they did not explore the relationship between personality and loneliness. In addition, the development of top-down prefrontal regions during adolescence is immature relative to the development of bottom-up striatal regions implicated in the detection of salient cues in the environment (Somerville and Casey 2010); that is, characteristics of adolescence, such as susceptibility to social rejection or isolation from others, are associated with deficiencies in the development in these brain areas (e.g., PFC, pSTS), which can in turn lead to loneliness. More studies are needed to explore these brain structures and their functional roles. The current study was conducted with the aim of bridging these gaps in the literature.
The current study aimed to explore the relationship between brain structures and loneliness (measured by the UCLA Loneliness Scale) (Russell 1996) in a relatively large sample of normal Chinese young adults (N = 308). We used VBM to identify the brain structures underlying individual differences in loneliness. Moreover, even though it is believed that personality traits (neuroticism, extraversion and psychoticism) are predictors of loneliness (Hojat 1982; Stokes 1985; Saklofske and Yackulic 1989; Halamandaris and Power 1999; Cheng and Furnham 2002; Atak 2009), the nature of the influence of personality on the relationship between loneliness and brain structure is unclear. Thus, we further tried to explore whether personality mediated the relationship between brain structure and loneliness. Specifically, we hypothesized that: (1) loneliness is correlated with certain personality traits (neuroticism, extraversion, and psychoticism); (2) individual differences in loneliness are associated with the regional gray matter volume (rGMV) in the brain regions responsible for emotion (e.g., PFC) and social perception (e.g., pSTS); and (3) personality traits (neuroticism, extraversion, and psychoticism) mediate the relationship between specific brain regions (identified using VBM) and loneliness. It was thought that the use of morphometric results combined with a multiple mediation analysis could further validate the influence of personality traits on loneliness.
Materials and methods
Participants
A total of 308 right-handed, healthy volunteers (167 females; age range 18–27 years, mean ± SD 19.94 ± 1.27) participated in this study, which was conducted as a part of our ongoing project to examine the association between brain imaging, creativity, and mental health. All participants were undergraduate or postgraduate students from Southwest University, China. All participants completed the University of California Los Angeles (UCLA) Loneliness Scale (Russell 1996) and the Eysenck Personality Questionnaire (EPQ) (Eysenck and Eysenck 1969), which has emerged as an important psychometric theory in the field of personality. None of the participants had a history of neurological or psychiatric illness. This study was approved by the local ethics committee of Southwest China University and by the Institutional Human Participants Review Board of Southwest University Imaging Center for Brain Research. All participants provided written informed consent prior to taking part in the study.
Assessment of loneliness
The UCLA Loneliness Scale was designed to assess subjective feelings of loneliness or social isolation. The scale comprises 20 items, which are scored on a four-point Likert-type scale, where 1 = Never and 4 = Always; the total scores range from 20 (low level of loneliness) to 80 (high level of loneliness). An example statement is, “How often do you feel that there is no one you can turn to?” Items are counterbalanced to account for social desirability. The scale has adequate internal consistency and test–retest reliability (Russell 1996) and was shown to have a high level of internal consistency (Cronbach’s α = 0.83) in our sample.
Assessment of EPQ
The EPQ is a self-report questionnaire on three different personality dimensions— neuroticism, extraversion, and psychoticism—and a validity dimension—lie (Eysenck and Eysenck 1969; Chen 1983). Participants are required to respond with “true” or “false” in response to each of the 85 statements. Gender-specific normative data were used to calculate the t-scores for neuroticism, extraversion, and psychoticism, which were the main variables of interest. The EPQ has demonstrated adequate reliability and validity (Chen 1983). In addition, the scale has shown satisfactory internal consistency for neuroticism (Cronbach’s α = 0.82), extraversion (Cronbach’s α = 0.80), and psychoticism (Cronbach’s α = 0.72) in our sample.
MRI data acquisition
MRI scans were acquired on a 3.0T Siemens Trio MRI scanner (Siemens Medical, Erlangen, Germany). High-resolution T1-weighted anatomical images were acquired using a magnetization-prepared rapid gradient echo (MPRAGE) sequence (repetition time = 1,900 ms; echo time = 2.52 ms; inversion time = 900 ms; flip angle = 9°; resolution matrix = 256 × 256; slices = 176; thickness = 1.0 mm; voxel size = 1 × 1 × 1 mm).
Image processing for VBM
MRI scans were processed using the Statistical Parametric Mapping software (SPM8; Welcome Department of Cognitive Neurology, London, UK [www.fil.ion.ucl.ac/spm]) implemented in MATLAB 7.8 (MathWorks Inc., Natick, MA, USA). First, MRI scans were assessed by manual inspection for artifacts or gross anatomical abnormalities. For better registration, the reorientation of the images was manually set to the anterior commissure. Second, the images were segmented into three distinct tissue classes—gray matter, white matter, and cerebrospinal fluid—using the new segmentation tool in SPM8. Third, diffeomorphic anatomical registration through exponentiated lie (DARTEL) algebra was used for registration, normalization, and modulation (Ashburner and Friston 2005; Ashburner 2007). Fourth, to ensure that regional differences in the absolute amount of gray matter were conserved and to preserve the volume of tissue from each structure after warping, gray matter voxel values were modulated by multiplying the Jacobian determinants derived from the normalization (Good et al. 2002). The registered images were then transformed to Montreal Neurological Institute (MNI) space. Finally, the normalized, modulated gray matter images were smoothed with a 10-mm full-width at half-maximum Gaussian kernel to increase the signal-to-noise ratio.
Statistical analysis of VBM
Statistical analyses of GMV data were performed using SPM8. For whole-brain analyses, we performed multiple linear regression analysis with loneliness as the independent variable and voxel-wise GMV as the dependent variable. Loneliness was used as the variable of interest in the multiple linear regression analyses. Age and gender were treated as nuisance variables because both of them may affect brain structure (Good et al. 2002; Sowell et al. 2003), and total GMV (tGMV) was entered as a nuisance variable due to differences in head size (Bendfeldt et al. 2009). We also applied explicit masking with a population-specific optimal threshold using the masking toolbox in SPM8 to restrict the search volume within gray matter and white matter (http://www0.cs.ucl.ac.uk/staff/g.ridgway/masking/). This approach reduced the risk of false negatives caused by overly restrictive masking (in comparison with the use of absolute or relative threshold masking), as potentially interesting voxels were excluded from the statistical analysis (Ridgway et al. 2009). For all analyses, multiple comparison correction was carried out on the whole-brain data using the voxel-level family-wise error (FWE) approach and the corrected threshold was set at P < 0.05.
Furthermore, to examine the correlation between loneliness and regional brain volume, regions of interest (ROIs) were defined by a sphere with a radius of 6 mm, centered on the peak coordinate of the cluster obtained in the above analysis. The correlation between loneliness and the mean GM volume of the ROI was calculated while controlling for age, gender, and tGMV.
Multiple mediation analyses
To test how personality impacts the relationship between loneliness and individual differences in brain structures, we employed a multiple mediation model using the SPSS macros for bootstrapping (Preacher and Hayes 2008). Multiple mediation models comprise two parts: (1) investigating the total indirect effect, namely whether the set of mediators (personality traits) transmit the effect of X (brain structures) to Y (loneliness); (2) testing hypotheses regarding individual mediators in the context of a multiple mediator model (i.e., investigating the specific indirect effect associated with each putative mediator). The total effect (c) of the independent variable on the dependent variable is the sum of the direct effect (c′) and all of the specific indirect effects (a 1.b 1 + a 2.b 2 +⋯), namely, c = c′ + a 1.b 1 + a 2.b 2 +⋯. The total indirect effect can also be calculated as c–c′. Estimates of all paths are computed using ordinary least squares (OLS) regression. An indirect effect was considered to be significant if its 95 % bootstrap confidence intervals from 2,000 bootstrap samples did not include zero. The indirect, direct, and total effects of the brain structures on loneliness were calculated while controlling for age, gender, and tGMV effects.
Results
Descriptive data analysis
Age, gender, UCLA loneliness scores, and EPQ t-scores for this sample are shown in Table 1. The results revealed that loneliness was positively correlated with neuroticism (r = 0.44, P < 0.001) and psychoticism (r = 0.17, P < 0.01), and negatively related to extraversion (r = −0.45, P < 0.001).
VBM analysis
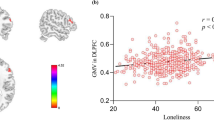
After controlling for age, gender, and tGMV, the results revealed a positive correlation between loneliness and the rGMV of a cluster, which mainly included areas in the left DLPFC (r = 0.27; cluster size = 76; t = 4.94; Brodmann areas, 9; MNI coordinates: −41, 12, 23; P = 0.007 corrected for multiple comparisons using the voxel-level FWE at the whole-brain level; see Fig. 1) (Benson et al. 2008; Causse et al. 2013). There were no negative correlations between loneliness and the rGMV of any brain area (P > 0.05, FWE corrected).

Correlation between regional gray matter volume (rGMV) and loneliness, as measured by the UCLA Loneliness Scale. a The left dorsolateral prefrontal cortex (DLPFC) exhibited significant positive correlation with loneliness (n = 308). The results are shown with a threshold of P[FWE-corr] = 0.007. b A scatter plot showing the correlation between loneliness and the rGMV of left DLPFC adjusted for age, gender, and total GMV (shown for illustrative purposes only)
Neuroticism and extraversion mediate the association between the left DLPFC and loneliness
To examine whether personality mediates the relationship between the brain structures identified in the aforementioned VBM study and loneliness, we conducted multiple mediation analyses. Table 1 provides Pearson’s correlations for the measures of EPQ (neuroticism, extraversion and psychoticism), loneliness, and the rGMV of left DLPFC identified in the VBM analysis. In addition, after controlling for age, gender, and tGMV effects, the correlations of the left DLPFC with loneliness, neuroticism, extraversion, and psychoticism were calculated. The correlations revealed that loneliness was significantly positively correlated with neuroticism (r = 0.44, P < 0.001) and psychoticism (r = 0.17, P < 0.01), and negatively associated with extraversion (r = −0.45, P < 0.001). Moreover, the rGMV of left DLPFC was positively correlated with neuroticism (r = 0.14, P < 0.05) and negatively correlated with extraversion (r = −0.12, P < 0.05). However, there was no correlation between the left DLPFC and psychoticism (P > 0.05) and the prerequisite for mediation was not satisfied, so it was concluded that psychoticism was not a mediator.
The total effect of the left DLPFC on loneliness was significant (c = 0.3381, P < 0.0001). After adjusting for the indirect effects of the mediators, the direct effect of the left DLPFC on loneliness was attenuated but remained significant (c′ = 0.1983, P = 0.0009). While these results suggest a partial mediation scenario, more recent statistical research suggests the importance of examining the indirect effects separately, given that the total effect is not necessary for mediation to occur (MacKinnon et al. 2000; Shrout and Bolger 2002; Preacher and Hayes 2008). Table 2 contains the parameter estimates for the total and specific indirect effects on the association between the left DLPFC and loneliness as mediated by neuroticism and extraversion. The total indirect effect and the indirect effects of neuroticism and extraversion were significant, as evidenced by the confidence intervals that did not contain zero. As illustrated in Fig. 2, the left DLPFC was significantly related to the two mediators, which in turn were significantly related to loneliness. Finally, there were no significant contrasts between the mediators, indicating that no mediator had a significantly greater indirect effect than other mediators on loneliness.

Paths linking the rGMV of left DLPFC to loneliness via neuroticism and extraversion. *P < 0.05, **P < 0.01
Discussion
This study investigated the association between brain structures and individual loneliness in a large sample and tried to explore which personality trait would be able to mediate the relationship between brain structure and loneliness. Our results showed that more lonely individuals had greater rGMV in the left DLPFC. Out of particular interest, we found that neuroticism and extraversion partially mediated the relationship between the left DLPFC and loneliness. The role of rGMV variations within the left DLPFC and the implication of the relationship between loneliness, the left DLPFC, and neuroticism (or extraversion) are discussed below.
Previous studies have indicated that the DLPFC plays a crucial role in working memory (Smith et al. 1998; Daskalakis et al. 2008; Hoppenbrouwers et al. 2013), executive functioning (Wagner et al. 2001), and emotion regulation (Banks et al. 2007; Rilling et al. 2007; Ruocco et al. 2013). The DLPFC is one of the brain regions implicated in emotional processing, particularly during down-regulation of negative emotional conditions (Davidson et al. 2000). For instance, certain aspects of emotional regulation have been associated with increased activity in prefrontal regions, which include the DLPFC and dorsomedial PFC (DMPFC) (Albaugh et al. 2013). Increased activity in the DLPFC was reported in an fMRI study during the processing of positive emotional stimuli in comparison with the processing of neutral and negative ones (Dolcos et al. 2004). Further, Ruocco et al. (2013) found bilateral reduction in activation within the DLPFC in bipolar disorder patients relative to control subjects, which likely reflects a diminished capacity for cognitive control in the modulation of subjectively experienced negative emotions. Furthermore, a number of studies using electrophysiological and/or functional neuroimaging techniques have reported consistently increased activity in the DLPFC under reappraisal conditions (modifying the intensity of emotional stimuli using cognitive strategies) (Ochsner et al. 2002; Kim and Hamann 2007). With regard to the involvement of the DLPFC in higher order cognitive functions such as working memory (Smith et al. 1998; Daskalakis et al. 2008), the findings of the previous studies suggest that lower working memory capacity is associated with less control over emotional responses and reduced emotional self-regulation following negative feedback (Schmeichel et al. 2008; Schmeichel and Demaree 2010). In brief, it is suggested that increased rGMV in the left DLPFC may contribute to loneliness through “inefficient emotion regulation” functions that are associated with this region (Dronkers et al. 2004; Binder et al. 2009; Turken and Dronkers 2011).
Recently, several studies have indicated that “the larger the volume, the better the function” premise does not always hold true (Kanai and Rees 2011; Takeuchi et al. 2011). For example, one of these studies reported an association between lower rostrolateral PFC volume and stronger self-referential ability, and a relationship between greater volume in the dorsal part of the ACC and increase in negative mood (Takeuchi et al. 2014b). Further, Kanai and Rees (2011) found that lower cortical volume can sometimes be associated with better task performance. Indeed, there is evidence that the gray matter of brain regions (e.g., prefrontal regions) can thin over time during the course of normal development (Sowell et al. 2003; Decety et al. 2004; Amodio and Frith 2006; Buckner et al. 2008; Schulte-Rüther et al. 2011). For instance, there is a decrease in GMV in the prefrontal regions during adolescence, which might represent more effective synaptic and/or neuronal pruning processes (i.e., the process of removing inefficient synapses and neurons over a lifetime) (Kanai and Rees 2011; Takeuchi et al. 2014a). Based on these findings, we think that the increased rGMV in the left DLPFC within prefrontal regions may be reflective of neuronal immaturity, which is mainly due to brain maturation characterized by cortical thinning such as effective synaptic pruning. Therefore, increased DLPFC volume is linked to more negative moods (inefficient emotion regulation), which in turn enhances an individual’s feeling of loneliness.
Interestingly, the current study found that neuroticism and extraversion partially mediated the relationship between the left DLPFC and loneliness. However, this mediation effect was not observed for psychoticism, which was also correlated with loneliness. The result might suggest that psychoticism contributes to loneliness, but its association with loneliness might be independent of the left DLPFC structure. Many previous papers have reported that neuroticism and extraversion show temporal stability and heritability (Birley et al. 2006; Bratko and Butkovic 2007; Ivkovic et al. 2007; Wray et al. 2007) and are strongly associated with loneliness (Stokes 1985; Saklofske and Yackulic 1989; Halamandaris and Power 1999; Atak 2009). In fact, our behavioral data also showed that the t-scores for neuroticism and extraversion were significantly related to loneliness. However, the nature of the influence of neuroticism and extraversion on the relationship between loneliness and brain structure is unclear. Based on the above discussion, we think that increased DLPFC volume is linked to more negative moods (inefficient emotion regulation); thus, a lack of inhibitory control (disrupted synaptic or neuronal pruning of the DLPFC) might facilitate the development of neuroticism and inhibit the development of extraversion, which is likely to exacerbate feelings of loneliness (e.g., negative thinking, social rejection, and isolation). Although we cannot determine the direction of causation among neuroticism (or extraversion), loneliness, and left DLPFC involvement, we hypothesize that the left DLPFC predicts neuroticism (or extraversion), which in turn influences and predicts loneliness.
Conclusions
The present study used VBM to examine the structural correlates of individual loneliness. We found a positive correlation between the rGMV of left DLPFC and loneliness. This finding indicated that inefficient emotional regulation may lead an individual to develop vulnerability to social rejection or isolation, thereby enhancing loneliness. In addition, we found that neuroticism and extraversion partially mediated the relationship between the left DLPFC and loneliness, that is, higher neuroticism or lower extraversion might be associated with increased rGMV in the left DLPFC, which may in turn be linked to individual loneliness. Neuroticism and extraversion might therefore play important roles in shaping an individual’s feeling of loneliness. In summary, through morphometric and multiple mediation analyses, this paper further validates the influence of both neuroticism and extraversion on loneliness. The implementation of longitudinal or intervention studies may further help to elucidate the complex relationships between loneliness, brain structure, and personality.
References
Albaugh MD, Ducharme S, Collins DL et al (2013) Evidence for a cerebral cortical thickness network anti-correlated with amygdalar volume in healthy youths: implications for the neural substrates of emotion regulation. Neuroimage 71:42–49
Amodio DM, Frith CD (2006) Meeting of minds: the medial frontal cortex and social cognition. Nat Rev Neurosci 7:268–277
Andersson L (1982) Interdisciplinary study of loneliness: with evaluation of social contacts as a means towards improving competence in old age. Acta Sociol 25:75–80
Ashburner J (2007) A fast diffeomorphic image registration algorithm. Neuroimage 38:95–113
Ashburner J, Friston KJ (2005) Unified segmentation. Neuroimage 26:839–851
Asher SR, Paquette JA (2003) Loneliness and peer relations in childhood. Curr Dir Psychol Sci 12:75–78
Atak H (2009) Big Five traits and loneliness among Turkish emerging adults. Int J Hum Soc Sci 4:749–753
Banks SJ, Eddy KT, Angstadt M, Nathan PJ, Phan KL (2007) Amygdala–frontal connectivity during emotion regulation. Soc Cogn Affect Neurosci 2:303–312
Bendfeldt K, Kuster P, Traud S et al (2009) Association of regional gray matter volume loss and progression of white matter lesions in multiple sclerosis—a longitudinal voxel-based morphometry study. Neuroimage 45:60–67
Benson BE, Willis MW, Ketter TA et al (2008) Interregional cerebral metabolic associativity during a continuous performance task (Part II): differential alterations in bipolar and unipolar disorders. Psychiatry Res 164:30–47
Binder JR, Desai RH, Graves WW, Conant LL (2009) Where is the semantic system? A critical review and meta-analysis of 120 functional neuroimaging studies. Cereb Cortex 19:2767–2796
Birley AJ, Gillespie NA, Heath AC, Sullivan PF, Boomsma DI, Martin NG (2006) Heritability and nineteen-year stability of long and short EPQ-R Neuroticism scales. Pers Individ Dif 40:737–747
Bratko D, Butkovic A (2007) Stability of genetic and environmental effects from adolescence to young adulthood: results of Croatian longitudinal twin study of personality. Twin Res Hum Genet 10:151
Buckner RL, Andrews-Hanna JR, Schacter DL (2008) The brain’s default network. Ann NY Acad Sci 1124:1–38
Canli T, Zhao Z, Desmond JE, Kang E, Gross J, Gabrieli JD (2001) An fMRI study of personality influences on brain reactivity to emotional stimuli. Behav Neurosci 115:33
Causse M, Péran P, Dehais F, Caravasso CF, Zeffiro T, Sabatini U, Pastor J (2013) Affective decision making under uncertainty during a plausible aviation task: an fMRI study. Neuroimage 71:19–29
Chen ZG (1983) Item analysis of eysenck personality questionnaire tested in Beijing-District (Article written in Chinese). Acta Psychol Sinica 2:211–218
Cheng H, Furnham A (2002) Personality, peer relations, and self-confidence as predictors of happiness and loneliness. J Adolesc 25:327–339
Daskalakis ZJ, Farzan F, Barr MS, Rusjan PM, Favalli G, Levinson AJ, Fitzgerald PB (2008) Evaluating the relationship between long interval cortical inhibition, working memory and gamma band activity in the dorsolateral prefrontal cortex. Clin EEG Neurosci 39:150–155
Davidson RJ, Putnam KM, Larson CL (2000) Dysfunction in the neural circuitry of emotion regulation—a possible prelude to violence. Science 289:591–594
Decety J, Jackson PL, Sommerville JA, Chaminade T, Meltzoff AN (2004) The neural bases of cooperation and competition: an fMRI investigation. Neuroimage 23:744–751
Dolcos F, LaBar KS, Cabeza R (2004) Dissociable effects of arousal and valence on prefrontal activity indexing emotional evaluation and subsequent memory: an event-related fMRI study. Neuroimage 23:64–74
Dronkers NF, Wilkins DP, Van Valin RD Jr, Redfern BB, Jaeger JJ (2004) Lesion analysis of the brain areas involved in language comprehension. Cognition 92:145–177
Eisenberger NI, Lieberman MD, Williams KD (2003) Does rejection hurt? An fMRI study of social exclusion. Science 302:290–292
Eisenberger NI, Lieberman MD, Satpute AB (2005) Personality from a controlled processing perspective: an fMRI study of neuroticism, extraversion, and self-consciousness. Cogn Affect Behav Neurosci 5:169–181
Eysenck SBG, Eysenck HJ (1969) Personality structure and measurement. Routledge & Kegan Paul, London
Eysenck HJ, Eysenck MW (1985) Personality and individual differences: a natural science approach. Plenum PressM, New York
Eysenck HJ, Eysenck SBG (1991) Manual of the Eysenck personality scales (EPS Adult). Hodder & Stoughton, London
Furnham A, Cheng H (1999) Personality as predictor of mental health and happiness in the East and West. Pers Individ Dif 27:395–403
Good CD, Johnsrude IS, Ashburner J, Henson RN, Fristen K, Frackowiak RS (2002) A voxel-based morphometric study of ageing in 465 normal adult human brains. In: 5th IEEE EMBS international summer school on biomedical imaging, 2002, IEEE, p 16
Halamandaris K, Power K (1999) Individual differences, social support and coping with the examination stress: a study of the psychosocial and academic adjustment of first year home students. Pers Individ Dif 26:665–685
Hojat M (1982) Loneliness as a function of selected personality variables. J Clin Psychol 38:137–141
Hoppenbrouwers SS, De Jesus DR, Stirpe T, Fitzgerald PB, Voineskos AN, Schutter DJ, Daskalakis ZJ (2013) Inhibitory deficits in the dorsolateral prefrontal cortex in psychopathic offenders. Cortex 49:1377–1385
Ivkovic V, Vitart V, Rudan I et al (2007) The Eysenck personality factors: psychometric structure, reliability, heritability and phenotypic and genetic correlations with psychological distress in an isolated Croatian population. Pers Individ Dif 42:123–133
Johnson HD, LaVoie JC, Spenceri MC, Mahoney-Wernli MA (2001) Peer conflict avoidance: associations with loneliness, social anxiety, and social avoidance. Psychol Rep 88:227–235
Kanai R, Rees G (2011) The structural basis of inter-individual differences in human behaviour and cognition. Nat Rev Neurosci 12:231–242
Kanai R, Bahrami B, Duchaine B, Janik A, Banissy MJ, Rees G (2012) Brain structure links loneliness to social perception. Curr Biol 22:1975–1979
Kardum I, Hudek-Knežević J (1996) The relationship between Eysenck’s personality traits, coping styles and moods. Pers Individ Dif 20:341–350
Kim SH, Hamann S (2007) Neural correlates of positive and negative emotion regulation. J Cogn Neurosci 19:776–798
Koenig LJ, Isaacs AM, Schwartz JAJ (1994) Sex differences in adolescent depression and loneliness: why are boys lonelier if girls are more depressed? J Res Pers 28:27–43
Kross E, Berman MG, Mischel W, Smith EE, Wager TD (2011) Social rejection shares somatosensory representations with physical pain. Proc Natl Acad Sci 108:6270–6275
Lasgaard M (2007) Reliability and validity of the Danish version of the UCLA Loneliness Scale. Pers Individ Dif 42:1359–1366
Lau S, Chan DWK, Lau PSY (1999) Facets of loneliness and depression among Chinese children and adolescents. J Soc Psychol 139:713–729
Loehlin J, Martin N (2001) Age changes in personality traits and their heritabilities during the adult years: evidence from Australian twin registry samples. Pers Individ Dif 30:1147–1160
MacKinnon DP, Krull JL, Lockwood CM (2000) Equivalence of the mediation, confounding and suppression effect. Prev Sci 1:173–181
McPherson M, Smith-Lovin L, Brashears ME (2006) Social isolation in America: changes in core discussion networks over two decades. Am Sociol Rev 71:353–375
Moore D, Schultz N Jr (1983) Loneliness at adolescence: correlates, attributions, and coping. J Youth Adolesc 12:95–100
Ochsner KN, Bunge SA, Gross JJ, Gabrieli JD (2002) Rethinking feelings: an fMRI study of the cognitive regulation of emotion. J Cogn Neurosci 14:1215–1229
Parlee MB (1979) The friendship bond. Psychology Today 13:43–54
Peplau LA, Perlman D (1982) Loneliness: a sourcebook of current theory, research, and therapy. Wiley, New York
Perlman D, Landolt MA (1999) Examination of loneliness in children-adolescents and in adults: two solitudes or unified enterprise? In: Rotenberg KJ, Hymel S (eds) Loneliness in childhood and adolescence. Cambridge University Press, Cambridge, pp 325–347
Perlman D, Peplau LA (1981) Toward a social psychology of loneliness. Pers Relatsh 3:31–56
Preacher KJ, Hayes AF (2008) Asymptotic and resampling strategies for assessing and comparing indirect effects in multiple mediator models. Behav Res Methods 40:879–891
Putnam RD (2000) Bowling alone: the collapse and revival of American community. Simon and Schuster, New York
Ridgway GR, Omar R, Ourselin S, Hill DL, Warren JD, Fox NC (2009) Issues with threshold masking in voxel-based morphometry of atrophied brains. Neuroimage 44:99–111
Rilling JK, Glenn AL, Jairam MR, Pagnoni G, Goldsmith DR, Elfenbein HA, Lilienfeld SO (2007) Neural correlates of social cooperation and non-cooperation as a function of psychopathy. Biol Psychiatry 61:1260–1271
Roberts RK, Roberts CR, Chen YR (1998) Suicidal thinking among adolescents with a history of attempted suicide. J Am Acad Child Adolesc Psychiatry 37:1294–1300
Ruocco AC, Amirthavasagam S, Choi-Kain LW, McMain SF (2013) Neural correlates of negative emotionality in borderline personality disorder: an activation-likelihood-estimation meta-analysis. Biol Psychiatry 73:153–160
Russell DW (1996) UCLA Loneliness Scale (Version 3): reliability, validity, and factor structure. J Pers Assess 66:20–40
Saklofske D, Yackulic R (1989) Personality predictors of loneliness. Pers Individ Dif 10:467–472
Schmeichel BJ, Demaree HA (2010) Working memory capacity and spontaneous emotion regulation: high capacity predicts self-enhancement in response to negative feedback. Emotion 10:739
Schmeichel BJ, Volokhov RN, Demaree HA (2008) Working memory capacity and the self-regulation of emotional expression and experience. J Pers Soc Psychol 95:1526
Schulte-Rüther M, Greimel E, Markowitsch HJ, Kamp-Becker I, Remschmidt H, Fink GR, Piefke M (2011) Dysfunctions in brain networks supporting empathy: an fMRI study in adults with autism spectrum disorders. Soc Neurosci 6:1–21
Sermat V, Hartog J (1980) Some situational and personality correlates of loneliness. In: Hartog J, Audy JR, Cohen YA (eds) The anatomy of loneliness. International Universities Press, New York, pp 305–318
Shrout PE, Bolger N (2002) Mediation in experimental and nonexperimental studies: new procedures and recommendations. Psychol Methods 7:422
Shute R, Howitt D (1990) Unravelling paradoxes in loneliness: research and elements of a social theory of loneliness. Soc Behav 5:169–184
Smith EE, Jonides J, Marshuetz C, Koeppe RA (1998) Components of verbal working memory: evidence from neuroimaging. Proc Natl Acad Sci 95:876–882
Somerville LH, Casey B (2010) Developmental neurobiology of cognitive control and motivational systems. Curr Opin Neurobiol 20:236–241
Sowell ER, Peterson BS, Thompson PM, Welcome SE, Henkenius AL, Toga AW (2003) Mapping cortical change across the human life span. Nat Neurosci 6:309–315
Stokes JP (1985) The relation of social network and individual difference variables to loneliness. J Pers Soc Psychol 48:981
Takeuchi H, Taki Y, Sassa Y, Hashizume H, Sekiguchi A, Fukushima A, Kawashima R (2011) Regional gray matter density associated with emotional intelligence: evidence from voxel-based morphometry. Hum Brain Mapp 32:1497–1510
Takeuchi H, Taki Y, Nouchi R et al (2014a) Regional gray matter density is associated with achievement motivation: evidence from voxel-based morphometry. Brain Struct Funct 219:71–83
Takeuchi H, Taki Y, Nouchi R et al (2014b) Anatomical correlates of quality of life: evidence from voxel‐based morphometry. Hum Brain Mapp 35:1834–1846
Turken AU, Dronkers NF (2011) The neural architecture of the language comprehension network: converging evidence from lesion and connectivity analyses. Front Syst Neurosci 5:1
Wagner AD, Maril A, Bjork RA, Schacter DL (2001) Prefrontal contributions to executive control: fMRI evidence for functional distinctions within lateral prefrontal cortex. Neuroimage 14:1337–1347
Williams D (1990) Effects of psychoticism, extraversion, and neuroticism in current mood: a statistical review of six studies. Pers Individ Dif 11:615–630
Wray NR, Birley AJ, Sullivan PF, Visscher PM, Martin NG (2007) Genetic and phenotypic stability of measures of neuroticism over 22 years. Twin Res Hum Genet 10:695–702
Zuckerman M (1989) Personality in the third dimension: a psychobiological approach. Pers Individ Dif 10:391–418
Acknowledgments
We thank Xi-Nian Zuo for his helpful comments on the data analysis. This work was supported by grants from the National Natural Science Foundation of China (31070900; 31271087), the Program for New Century Excellent Talents in University (2011) by the Ministry of Education, the Fundamental Research Funds for the Central Universities (SWU1209101) and the Key Discipline Fund of National 211 Project (TR201208-1).
Conflict of interest
The authors declare no competing interests.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
Kong, X., Wei, D., Li, W. et al. Neuroticism and extraversion mediate the association between loneliness and the dorsolateral prefrontal cortex. Exp Brain Res 233, 157–164 (2015). https://doi.org/10.1007/s00221-014-4097-4
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00221-014-4097-4