
Abstract
During adaptive locomotion, vision is used to guide the lead limb; however, the individual must rely on knowledge of obstacle height and position, termed obstacle memory, to guide the trail limb. Previous research has demonstrated that visual sampling of the obstacle during approach was adequate to provide obstacle height information, but online visual update of distance to the obstacle was required to plan and implement appropriate foot placement. Our purpose was to determine whether obstacle height memory, coupled with a visible obstacle position cue, could successfully guide the foot during obstacle crossing. Subjects first stepped over an obstacle for 25 trials; then, the obstacle was removed, but its position was marked with high-contrast tape; subjects were instructed to step over the obstacle as if it was still there (termed “virtual obstacle”) for 25 trials. No changes in foot placement were observed; therefore, the position cue provided salient online information to guide foot placement. Average failure rates (subject would have contacted the virtual obstacle if it was present) were 9 and 47 % (lead and trail limb, respectively). Therefore, action was impaired for both limbs when guided by obstacle height memory, but action was impaired to a greater extent for the trail limb. Therefore, viewing the obstacle during approach appears to facilitate the memory needed to guide obstacle crossing, particularly for the trail limb. This is likely because the lead limb is visible in the peripheral visual field during crossing, but the trail limb is not.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
It is well recognized that locomotor tasks are completed under continuous control based on visual information (e.g., Lee et al. 1982; Patla 1998). When stepping over an obstacle, the first limb (leading limb) is visible in the lower visual field, and online visual information is used to control the lead limb trajectory (Patla et al. 1996; Patla 1998; Mohagheghi et al. 2004; Rietdyk and Rhea 2006, 2011). However, when the trailing limb clears the obstacle, the limb and the obstacle are not visible, so the individual must rely on knowledge of obstacle characteristics to control the trail limb trajectory. These characteristics likely include spatial characteristics, such as height, position, and depth (Patla and Rietdyk 1993), and perceived characteristics, such as fragility (Patla et al. 1996). This visuospatial knowledge has been termed as “stored obstacle representation” (Lajoie et al. 2012) or an “obstacle memory” (McVea and Pearson 2006, 2007; Whishaw et al. 2009; Shinya et al. 2012).
The concept that a representation is used to guide motor output is controversial, especially when vision is available (e.g., Warren 2006; Anson et al. 2010). However, when vision is not available, retained knowledge of spatial characteristics of a target is used to control upper limb reaching tasks (Milner et al. 2003; Binsted et al. 2006; Heath et al. 2010). Similarly, in locomotor research, it has been demonstrated that quadrupeds retain obstacle characteristics for long period of time. In one set of studies, cats stepped over an obstacle with the forelimbs and paused to eat; during the pause, the obstacle was lowered. When gait resumed, the hind limb trajectories clearly demonstrated that the cat remembered the obstacle and modified the trajectory based on the obstacle size and position (McVea and Pearson 2006, 2007). Similar findings were observed in horses (Whishaw et al. 2009). In humans, Lajoie et al. (2012) demonstrated that the trail leg trajectory was scaled appropriately to obstacle height after straddling an obstacle for up to 2 min. In the preceding studies, the obstacle was visible prior to and during lead limb crossing, which may have helped establish the memory. However, participants also successfully crossed obstacles when an obstacle was viewed during approach and when vision was removed during the last three steps before obstacle crossing (Mohagheghi et al. 2004). When vision was removed earlier (five steps before obstacle crossing), subjects were only 50 % successful (Patla and Greig 2006). In the latter study, the main cause of failure was not inappropriate limb elevation, but rather incorrect foot placement. The authors concluded that while initial visual sampling was adequate to provide obstacle height information, online visual update of distance to the obstacle was required to plan and implement appropriate foot placement. Our goal was to extend this line of research to determine whether action can be accurately guided by an obstacle memory when online visual distance to the obstacle was available.
In the present study, subjects were instructed to step over an obstacle that was not physically present (termed a virtual obstacle), in the same manner as upper limb aiming paradigms where the target is initially visible, but is not visible during the aiming movement (Elliott 1988; Binsted and Heath 2005; Heath 2005). Subjects stepped over an actual obstacle 25 times (epoch 1) before they stepped over the virtual obstacle 25 times (epoch 2). Obstacle clearance performance was quantified at two levels: (1) whether the subject would have contacted the virtual obstacle if it had been present and (2) differences in trajectory characteristics when crossing a real versus virtual obstacle. The average obstacle contact rate is about 1–2 % in young, healthy subjects in a research setting (Mohagheghi et al. 2004; Berard and Vallis 2006; Rietdyk and Rhea 2006; Rhea and Rietdyk 2007; Heijnen et al. 2012a), so we quantified successful performance as a virtual obstacle contact rate of 5 % or less. Given the empirical support for long-lasting obstacle memories, the hypotheses were developed in support of the obstacle memory successfully guiding action. We hypothesized that (1) subjects would successfully clear the virtual obstacle at least 95 % of the time and (2) trajectory characteristics would be similar for the real and virtual obstacles. Further, we hypothesized that failure rate and trajectory characteristics would not change during the course of epoch 2. This would demonstrate that the obstacle memory did not decay over the 9-min epoch.
After 19 subjects were collected, it was noted that the trail limb failure rate (47 %) was about four times greater than the lead limb failure rate (9 %). Therefore, we added a condition to the following 21 subjects to determine whether the high trail limb failure rate was due to inadequate instructions for the trail limb. We found that trail limb failure remained higher than lead limb failure. As the failure rate was substantially larger than hypothesized, we also completed a second study to determine whether subjects would scale the trajectories to different heights of virtual obstacles. We found that subjects scaled the lead and trail limb trajectories to the virtual obstacle height, confirming that an obstacle memory was being used to guide the trajectory despite high failure rates.
Methods
Experiment 1
Forty-one subjects were recruited from a university population, and two were excluded due to data collection problems, resulting in 39 total subjects (22.1 ± 2.4 years, 18 males). Subjects were free from any impediments to normal locomotion, as verified by self-report. All subjects signed a consent form approved by the local institutional review board. Subjects were instrumented with eight infrared emitting diodes (IREDs). Six IREDs were placed on the lateral aspect of the left foot at the distal phalanx of the third toe, calcaneus, and malleolus and on the medial aspect of the right foot at the distal phalanx of the first toe, calcaneus, and malleolus. Two IREDs were placed on the lateral side of the head. One IRED was placed on the lateral side of the obstacle. Two Optotrak 3020 sensors (NDI, Canada) recorded the position data of the IREDs at 60 Hz. The obstacle was composed of masonite (painted flat black) with two supports (L-brackets) mounted on the leading face of the obstacle, such that if the subject contacted the obstacle, it would fall forward without arresting the swing limb. The obstacle was 100 cm wide by 0.3 cm deep, and the height was 25 % of the subject’s leg length (range 19.5–26.0 cm in 0.5 cm increments). For each subject, before the experiment began, the starting position was adjusted such that the right foot was naturally the lead foot (first foot to cross obstacle). The right limb was set as the lead foot because we have previously observed that when subjects self-select the lead limb, occasionally a subject will switch between right and left as the lead limb; these intermixed trajectories were qualitatively different for some subjects (unpublished observations). These differences would have confounded the comparison across real and virtual obstacles if the subject used different lead limbs for the real and virtual obstacles; thus, we used the right limb as the lead limb to eliminate this confound. After the starting position was determined, subjects were instructed to always cross the obstacle with the right foot first. The obstacle position was marked with masking tape (100 cm long). Two obstacle conditions were observed as follows: the obstacle was in place (real obstacle) or the obstacle was not in place (“virtual obstacle” located at masking tape).
Before data collection, subjects were instructed as follows: “The obstacle will be in place for the first 25 trials and will be removed for next 25 trials. When the obstacle is not there, you will be asked to step over the tape as if the obstacle was still there.” In the first 25 trials (epoch 1), subjects walked down an 8-m walkway at a self-selected pace, stepped over the obstacle in the middle of the walkway, and continued walking. The obstacle was removed in the second epoch of 25 trials, but the obstacle position tape remained (epoch 2). At the beginning of epoch 2, subjects were instructed “Step over the piece of tape as if the obstacle was still in place and cross the obstacle with your right leg first.” The obstacle was returned to the walkway for the third epoch of 25 trials (epoch 3). Epoch 3 was used to ensure that any changes across real and virtual trajectories (epochs 1 and 2) were not a simple adaptation due to repeated crossings (Rhea et al. 2010; Heijnen et al. 2012a). A fourth 25-trial epoch was added for the latter 21 subjects to investigate the influence of instruction on the dependent variables (epoch 4). The obstacle was removed in epoch 4 as in epoch 2, and subjects were instructed “Step over the piece of tape as if the obstacle was still in place. Make sure that you cross the obstacle with your right leg first and remember to also step over the obstacle with the left leg.”
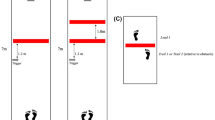
Data were analyzed with MATLAB 2010a software (MathWorks Inc., MA, USA). Data were filtered off-line at 8 Hz with a fourth-order zero-phase-shift low-pass Butterworth digital filter (Winter 2009). Dependent variables were minimum foot clearance (MFC), toe peak elevation, toe peak position relative to the obstacle, horizontal distance, stride length (SL), and failure rate. Variability measures were calculated as the standard deviation. High limb velocity during crossing can compromise clearance accuracy (up to 17 % error), so spatial resolution was increased with a cubic interpolation algorithm (Heijnen et al. 2012b). Failure could result from either the forefoot or rearfoot region of the foot passing through the obstacle (Chen et al. 1991; Thies et al. 2011; Heijnen et al. 2012a; Loverro et al. 2013; Telonio et al. 2013). Therefore, both toe and heel clearances were calculated: Toe/heel clearance was the vertical distance between the IREDs on the toe/heel and obstacle as the toe/heel crossed the obstacle. The minimum of the toe and heel clearance for each trial was quantified as MFC. A negative MFC indicated failure; failure magnitude was quantified as the average of the negative MFC. Toe peak was the maximum vertical distance between the toe and the ground. Toe peak position was the anterior–posterior distance of the IREDs on the toe relative to the obstacle at toe peak. A negative value indicated that toe peak occurred before the toe crossed the obstacle (e.g., subject 19, virtual obstacle trajectories of lead limb, Fig. 1), and a positive value indicated that toe peak occurred after the toe crossed the obstacle. Horizontal distance was calculated as the anterior–posterior distance between the IREDs on the toe and obstacle at toe-off. SL was calculated as the anterior–posterior distance between the toe marker at toe-off and the subsequent toe-off of the same limb. Overall failure rate was calculated as the percentage of obstacle contacts if the obstacle had been in place in the virtual obstacle condition; failure was determined for the lead and trail limbs separately. Subjects were classified as successful, achieving a failure rate of 5 % or less, or not successful. If the obstacle memory degraded during epoch 2, we would expect the failure rate and MFC to change. To quantify this, trial-specific failure rate was calculated as the percent of subjects who failed in each of the trials in epoch 2 and a linear regression was performed. Further, a linear regression of MFC for all trials in epoch 2 was calculated for each subject individually. All variables were calculated for both the lead and the trail limb.

Toe trajectories of the lead (left panel) and trail limb (right panel) for five subjects. Subjects stepped over a real obstacle in epoch 1 (black lines) and crossed a “virtual” obstacle in epoch 2 (gray dashed lines)
Experiment 2
Twenty-four subjects were recruited from a university population (20.2 ± 0.9 years, 8 males). Subjects were free from any impediments to normal locomotion, as verified by self-report. All subjects signed a consent form approved by the local institutional review board. Instrumentation and methods were identical to Experiment 1 with the following exceptions: two obstacle heights were examined, 15 and 25 cm. The obstacles were presented in a blocked manner, with order of presentation counterbalanced. For example, when the small obstacle was presented first, epochs 1–4 were as follows: real 15 cm, virtual 15 cm, real 25 cm, and virtual 25 cm, respectively. Toe peak was compared between the 15- and 25-cm virtual obstacles.
Rationale for using real and virtual obstacles in a series of epochs
The virtual obstacle approach was adapted from the common experimental paradigm to examine visual control of reaching: The target is initially visible but disappears before the target is reached (Elliott 1988; Milner et al. 2003; Binsted and Heath 2005; Heath 2005). In the same manner, subjects here were instructed to step over an obstacle that was not physically present. This preserved vision of the environment and allowed the examination of memory-guided action for both the lead and trail limbs. To create the obstacle memory, the subject could simply have been shown the actual obstacle. However, walking upstairs facilitated the stair height memory compared to when information was obtained by vision alone (Shinya et al. 2012). Therefore, to increase the likelihood of generating a robust memory, subjects stepped over an actual obstacle 25 times (epoch 1) before they stepped over the virtual obstacle 25 times (epoch 2). The 25 trials in epoch 2 also allowed us to examine whether the memory degraded over time, as it takes about 9 min to collect 25 trials in young healthy adults.
A within-subject ANOVA was used to examine the effect of epoch (four levels) on each dependent variable. A generalized linear mixed model was used to allow the residuals to vary (GLIMMIX in SAS 9.3, Cary, NC, USA). Due to the large number of dependent variables, the p value was set to 0.01. Tukey–Kramer post hoc analyses were used to determine whether behavior changed due to repeated exposures within the real obstacle condition (epoch 1 vs 3), to determine whether behavior changed for real versus virtual obstacles (epoch 1 vs 2), and to determine whether toe peak changed for the virtual 25-cm obstacle versus the virtual 15-cm obstacle (Experiment 2). A generalized linear mixed model was used to test for differences in failure rate.
Results
Experiment 1
Contacts with the real obstacle
Ten contacts with the real obstacle were observed in nine subjects, for a contact rate of 0.5 %. Ninety percent of the contacts were with the trail limb. A large increase in toe clearance has been observed in subsequent trials after obstacle contact (Alexander et al. 2011; Rhea and Rietdyk 2011; Heijnen et al. 2012a), so these nine subjects were excluded from further analyses to ensure that any changes in clearance were due to the independent variable manipulation, and not in response to the contact. The remaining 30 subjects were included in further analyses, 15 have observations in epochs 1–3, and the remaining 15 have observations in epochs 1–4.
Qualitative comparisons of real and virtual trajectories
Lead toe trajectories for the real and virtual obstacles were similar, but a generally lower elevation and an earlier peak are noted in most subjects (Fig. 1, right panel, subjects 8, 13, and 19). Failures are demonstrated by the trajectory passing through the obstacle (e.g., subject 8); note that failures also resulted from the heel trajectory passing through the obstacle (not shown). Marked differences in the trail limb trajectories were readily apparent, with high intersubject variability (Fig. 1, right panel). Subjects 8 and 13 demonstrate large undershoot, and subjects 2 and 19 demonstrate large overshoot, with subject 2 pulling the limb backwards. When subjects were given further instruction with the trail limb, some subjects improved (e.g., subject 22, Fig. 2, right panel), but the majority demonstrated the same general trajectories with the virtual obstacle (e.g., subjects 24 and 41, Fig. 2).

Toe trajectories of the lead (left panel) and trail limb (right panel) for three subjects. Subjects stepped over a “virtual” obstacle in epoch 2 (black dashed lines) and epoch 4 (gray dashed lines); in epoch 4, subjects received more instruction than in epoch 2
Failure rate with the virtual obstacle
Overall failure rates with the virtual obstacle (epoch 2) were 9 and 47 %, for the lead and trail limb, respectively (Table 1; Fig. 1). Therefore, hypothesis 1, that subjects would successfully clear the virtual obstacle at least 95 % of the time, was rejected for both the lead and trail limb. The failure rate was significantly higher for the trail limb versus the lead limb (p < 0.001). Trial-specific failures did not change during epoch 2 for the lead limb, but trail failure increased 10 % in a linear fashion during epoch 2 (p = 0.001; Fig. 3). Failures during epoch 2 were examined for individual subjects, and it was observed that the subjects who were initially successful tended to remain successful and subjects who were initially unsuccessful tended to remain unsuccessful; therefore, these two groups did not affect the failure rate. However, there were four subjects (13 %) who changed from unsuccessful to successful during epoch 2, and these four subjects were responsible for the change in average trail trial-specific failure. Therefore, although the significant increase in trail failure during epoch 2 appears to indicate memory decay, it was being driven by only 13 % of the subjects.

Failure rate calculated for each trial during epoch 2 (virtual obstacle). Closed circles represent percent of subjects who failed with the lead limb, and open circles represent percent of subjects who failed with the trail limb. Failures did not change for the lead limb during the epoch, but trail failure significantly increased (p = 0.001)
Classification of successful and unsuccessful subjects
Nineteen subjects (63 %) were classified as lead limb successful (achieved a lead failure rate of 5 % or less in epoch 2 (e.g., subjects 2, 13 & 32 Fig. 1), and 11 (46 %) were classified as lead limb unsuccessful (e.g., subject 8, 19 Fig. 1; Table 1). Thirteen subjects (43 %) were classified as trail limb successful (e.g., subjects 2, 19 & 32 Fig. 1; Table 1), and 17 (67 %) were classified as trail limb unsuccessful (e.g., subjects 8 & 13 Fig. 1). Ten subjects (33 %) were classified as successful with both the lead and trail limb (e.g., subjects 2 & 32 in Fig. 1; Table 1). For those subjects with failed trials, the average magnitude of the failure was 1.7 ± 1.0 and 8.4 ± 2.4 cm for the lead and trail limb, respectively.
Failure rate as a function of instruction
This analysis included only the latter 15 subjects with observations in epochs 2 and 4, so epoch 2 average failure rates are slightly different from those reported above for all 30 subjects. Lead limb failure rate decreased from 14 to 7 % from epochs 2 to 4, respectively (p = 0.004; Table 1; Fig. 2). Trail limb failure rate decreased from 57 to 47 % from epochs 2 to 4, respectively (p < 0.001). Therefore, hypothesis 4, that trail limb failure rate will decrease with more specific instruction, was accepted. However, lead limb failure rate also decreased to a similar extent. Note the high variability in the improvement: trail limb failure rate improved more than 20 % with instruction for three subjects (subjects 22, 23, and 38 improved 84, 60, and 27 %, respectively), but subject 41 had 20 % higher failure (Table 1; Fig. 2).
Change in minimum foot clearance during epoch 2
There was no change in lead MFC for the majority of the subjects (80 %) during epoch 2. Of the six subjects (20 %) with slopes that were significantly different from zero in epoch 2, MFC increased for four subjects and decreased for two subjects. Similarly, there was no change in trail MFC for the majority of subjects (83 %). Of the five subjects (17 %) with slopes that were significantly different from zero: MFC increased for two subjects and decreased for three subjects. Since there was no consistent change in lead or trail MFC for the majority of subjects, the average of all 25 trials from epoch 2 was used for the remaining analyses. Therefore, there was no evidence of memory decay in the MFC of either lead or trail limb in epoch 2.
Adaptation effects for real obstacle; epoch 1 versus 3
Epoch 1 was not different from epoch 3 for all measures, demonstrating that subjects did not adapt their behavior as a function of repeated observations with the real obstacle (Figs. 4, 5). Therefore, any differences between epoch 2 and epoch 1 are due to the virtual obstacle manipulation.

Mean (left panel) and variability (right panel) of dependent variables during epochs 1–4 (E1, E2, E3, and E4). E1 and E3 have a real obstacle, E2 and E4 have a virtual obstacle. Lead minimum foot clearance (a), trail minimum foot clearance (b), lead toe peak elevation (c), trail toe peak elevation (d), variability of lead minimum foot clearance (e), variability of trail minimum foot clearance (f), variability of lead toe peak elevation (g), and variability of trail toe peak elevation (h). Error bars indicate standard error. Different letters indicate significant differences (p < 0.01). NS indicates no significant effect of epoch

Mean (left panel) and variability (right panel) of dependent variables during epoch 1–4 (E1, E2, E3, and E4). E1 and E3 have a real obstacle, E2 and E4 have a virtual obstacle. Lead toe peak position (a), trail toe peak position (b), variability of lead toe peak position (c), and variability of trail toe peak position (d). Error bars indicate standard error. Different letters indicate significant differences (p < 0.01). NS indicates no significant effect of epoch
Real versus virtual obstacle; epoch 1 versus 2
Post hoc analyses revealed that the following measures all decreased for the virtual obstacle (p ≤ 0.001 for all measures): lead and trail MFC (Fig. 4a, b), trail toe peak (Fig. 4d), and lead toe peak position (Fig. 5a). The following variability measures increased for both the lead and trail limbs (p ≤ 0.001): MFC variability (Fig. 4e, f), toe peak variability (Fig. 4g, h), and toe peak position variability (Fig. 5c, d). There were no differences in horizontal distance or stride length.
Experiment 2
Contacts with the real obstacle
Six contacts with the real obstacle were observed in six subjects, for a contact rate of 0.5 %; all contacts were with the trail limb. These six subjects were excluded from further analyses, resulting in 18 subjects. Toe peak was examined to determine whether subjects elevated the foot the same amount for real and virtual obstacles. Similar to Experiment 1 (Fig. 4c), lead toe peak was not different for real versus virtual obstacles for both the 15- and 25-cm obstacles. Trail toe peak was not different for real versus virtual obstacles for the 15-cm obstacle, but was lower for the 25-cm virtual obstacle compared to the large real obstacle (p < 0.001), consistent with Experiment 1 (Fig. 4d). Next, toe peak of the virtual trajectories was compared for 15- versus 25-cm obstacles, to determine whether the limb elevation when guided by memory was scaled to the obstacle height. Toe peak was significantly higher for the 25-cm versus 15-cm virtual obstacle for the lead foot (38.7 ± 3.2 vs 30.2 ± 3.1 cm, p < 0.001) and the trail foot (40.7 ± 5.5 vs 32.3 ± 4.9 cm, p < 0.001). These changes confirmed that subjects were scaling the trajectory to the height of the virtual obstacle.
Discussion
The aim of this paper was to determine whether an obstacle memory could guide action. The obstacle was not present and thus did not provide online visual guidance. However, unlike previous research (Mohagheghi et al. 2004; Patla and Greig 2006), vision of the environment was preserved, including a visible obstacle position cue, so that we specifically examined whether obstacle height memory could accurately guide action. No changes in foot placement were observed; therefore, the position cue provided salient online information to guide foot placement. However, the failure rates of 9 and 47 % for the lead and trail limb, respectively, indicate that the ability to successfully clear the virtual obstacle was compromised when relying on an obstacle memory. The higher variability of the trajectory characteristics demonstrate reduced precision when relying on an obstacle memory. In order to accept the preceding interpretations, it is important to demonstrate that subjects were genuinely attempting to clear the obstacle and that an obstacle memory was formed. Genuine attempts to clear the obstacle are confirmed by the observation that lead toe peak height was not different for real and virtual obstacles (Fig. 4c) and the clear attempts of subjects 2 and 19 to clear the obstacle (Fig. 1). The observation that both lead and trail toe peaks were scaled to the height of the virtual obstacle in Experiment 2 confirms that an obstacle memory had been formed. Therefore, it appears that participants were using an obstacle memory to guide action, but the action was compromised, perhaps due to an imprecise obstacle memory. We first describe why subjects failed with the lead limb, why they failed more frequently with the trail limb, and then consider why action was not successfully guided by an obstacle height memory, given the empirical support for long-lasting obstacle memories (McVea and Pearson 2006, 2007; Whishaw et al. 2009; Lajoie et al. 2012; Shinya et al. 2012).
The lead limb trajectories were qualitatively similar for real and virtual obstacles (Figs. 1, 2), although differences in the kinematics appear to reflect a position misjudgment and reduced precision. While toe peak was not different, the position of the toe peak within the stride was shifted backwards (Fig. 5a), and the clearance was reduced for the virtual obstacle (Fig. 4a), due to a steeper descent after the peak (Fig. 1, subjects 8, 13, and 19). These changes may reflect a position misjudgment where the subject perceived that the virtual obstacle was closer to the stance foot. An obstacle position misjudgment was unexpected because the position of the lower edge was provided with a high contrast length of masking tape and there was no difference in foot placement. If information regarding obstacle height and position of the lower edge is available, it is reasonable to expect that the position of the top edge is also available. However, the 9 % failure, the decrease in clearance, and the shifted location of the toe peak are not consistent with this expectation. It should be pointed out that while the average lead limb failure rate was 9, 63 % of subjects did achieve 5 % failure or less. However, the higher variability of clearance, toe peak, and toe peak position (Figs. 4e, g, 5c) for the virtual obstacle demonstrate reduced precision. Further, the average clearance of the failed trials was 1.7 cm, which is moderately high (about 8 % of the obstacle height). Overall, these findings indicate that an obstacle height memory provided some success with the lead limb, although not as high as predicted, but the action was compromised.
The trail limb failure rate (47 %) was almost ten times greater than the predicted failure rate of 5 % (Table 1). What is most striking during trail limb crossing of a virtual obstacle is the wide variety of behavior apparent in the trajectories (Figs. 1, 2). Subject 2 moved the foot backwards up to 25 cm after toe-off and elevated the toe up to 70 cm for a 19.5-cm obstacle. The peaks of subjects 8 and 13 only reach about half the height of the obstacle. Subject 19 increased toe clearance 250 %, and subject 32 adopted a trail limb trajectory with a triangular shape that was more similar to lead limb trajectories. The high trail limb failure rate, coupled with the qualitative changes in the trajectories (Fig. 1) and the large quantitative changes in the means and variability of the trajectory characteristics (Figs. 4, 5), clearly demonstrates that relying on an obstacle height memory compromised the control of the trail limb trajectory.
These failures and high trajectory variability are strikingly different from the successes observed in the previous literature (McVea and Pearson 2006, 2007; Whishaw et al. 2009; Lajoie et al. 2012). The main difference with the preceding studies is that the obstacle was visible during approach and/or lead limb crossing. Therefore, for both the lead and trail limb, it appears that the obstacle must be viewed during approach to form a memory that can successfully guide the action. It was not adequate to view the position (masking tape) and combine that information with obstacle height memory. Memories formed during approach and memories formed from previous experience may reside in separate systems for spatial representation (Milner and Goodale 1995; Milner et al. 2003); these separate systems would explain the differences between this study and previous obstacle crossing research. The dorsal system is responsible for the immediate guidance of action, while the ventral system is involved in delayed guidance of action. Previous research, where the obstacle was viewed during approach, would likely involve memory related to the dorsal visual stream. The dorsal visual stream projects to the parietal cortex, and neurons in the parietal cortex are active transiently when an animal steps over an obstacle (Drew et al. 2008) and remain active when an animal straddles the obstacle (Lajoie et al. 2010). The current paradigm would have relied on visual information that lasts longer than the transient information available within the dorsal stream. The more persistent—and less precise—visual information of the ventral stream would be used (Milner et al. 2003), leading to a high failure rate and reduced precision in trajectory control. This interpretation is similar to that of Shinya et al. (2012): Climbing stairs after vision was diverted for a few seconds appears to involve the less precise ventral system.
There is little evidence that the obstacle memory degraded during the course of epoch 2 (about 9 min), which likely reflects that participants were already relying on the less precise ventral system from the first trial in epoch 2. The only support for memory decay was the increase in trail failure (Fig. 3), but the increase was driven by only four subjects (13 %). The lack of decay was likely due to the relatively long interval, approximately 30 s, between crossing the last real obstacle in epoch 1 and the first virtual obstacle in epoch 2. In a similar approach with stair climbing, maximum toe clearance increased most within a 2-s period between diverting vision from the stair and step initiation. Therefore, the paradigm adopted here did not allow for evidence of decay. The lack of change reflects that the less precise obstacle memory, presumably from the ventral system, was relatively stable over the 9-min interval.
The high failure rates in the virtual obstacle condition support Gibson’s argument that dynamic visual sampling, achieved during the approach to the obstacle, is beneficial for the guidance of action (Gibson 1958, 1966). The obstacle memory would likely be a static representation and would likely be devoid of the rich information gained by viewing the obstacle while moving through the environment. These observations also build on previous research that demonstrated that vision of the interface of the obstacle and walkway (the lower edge) is important for successful crossing (Rietdyk and Rhea 2011). Therefore, it appears that the full obstacle must be visible, top and lower edge, at least three steps before crossing the obstacle in order to successfully guide the limb trajectory.
If the lead limb trajectory was used to calibrate or control the trail limb, one would expect that the failure rates would be similar for the two limbs, but substantially different failure rates were observed (Table 1; Figs. 1, 3). These differences add to the converging evidence that the limbs are controlled independently during obstacle crossing in humans (Patla et al. 1996; Niang and McFadyen 2004; Yang et al. 2004; Rhea and Rietdyk 2011). The observation that an obstacle memory was more successful at guiding the lead than the trail limb can be interpreted two ways. First, the instructions in epoch 2 reminded the subject to cross the obstacle with the right (lead) limb first, but did not specifically refer to the trail limb. In dual-task paradigms, subjects perform better in the task that they are instructed to attend to (Siu and Woollacott 2007; Kelly et al. 2010; Yogev-Seligmann et al. 2010). Since the original instruction referred to the lead limb, but not the trail limb, the instruction may have resulted in the subject paying more attention to the lead limb. When instruction referred to both lead and trail limb in epoch 4, failure rate decreased significantly for both limbs (Table 1; Fig. 2). Therefore, subjects apparently perceived that both lead and trail trajectories were not adequate and compensated with both limbs. However, the trail limb failure rate was still higher. Therefore, the observations do not support instruction as a plausible explanation for the high trail limb failure rate. The second explanation is that information of limb position relative to the obstacle (termed exproprioception) was compromised more for the trail limb than the lead limb. The trail limb action is guided by obstacle memory combined with kinesthetic information regarding current limb position and motion. The lead limb action is also guided by memory combined with kinesthetic information, but online visual information (the thigh is visible in the lower periphery) is also available and is likely used to update and calibrate the movement during the swing phase (Patla 1998; Rietdyk and Rhea 2006). Therefore, viewing the obstacle during approach is more critical for successful trail limb crossing than lead limb crossing.
More frequent trail contacts, observed here with virtual obstacles, are also observed with real obstacles. Therefore, the current findings may provide insight into the causes of trail limb failures with real obstacles. Trips can occur due to unexpected changes in surface height, but they also occur when an individual perceived an obstacle, but failed to elevate the limb adequately. In the lab setting, when young, healthy adults contact a visible, stationary obstacle under the conditions of normal lighting and full vision, the contact rate is about 1–2 % (Rietdyk and Rhea 2006; Rhea and Rietdyk 2007), and the trail foot is the contact foot 67–100 % of the time (Mohagheghi et al. 2004; Berard and Vallis 2006; Rietdyk and Rhea 2006, 2011; Rhea and Rietdyk 2007, 2011; Heijnen et al. 2012a). Note that the contacts with the real obstacles in the current paradigm have a similar rate (0.5 %) and were also mostly trail limb contacts (90 & 100 %). One potential mechanism behind these trail limb contacts with a real obstacle is that visual fixations during approach were absent or inadequate. There is a fair amount of inter and intrasubject variability in obstacle fixations during approach to an obstacle, and in up to 33 % of the trials, subjects did not fixate on the obstacle at all (Patla and Vickers 1997). This would result in insufficient visual information to successfully guide the action with the dorsal system, and the individual would presumably be forced to rely on the less precise ventral system. Lack of adequate visual information should compromise both lead and trail limb trajectories, but the lead limb trajectory can be updated by online visual information while the trail limb cannot, ultimately resulting in higher trail limb contact rates.
The virtual obstacle trail limb trajectories reported here for young healthy subjects have similarities with real obstacle trajectories described in balance-compromised subjects in two studies. First, subject 2 (Fig. 1) and another subject (not shown) demonstrated backwards displacement of the trail limb after toe-off for the virtual obstacle. A similar backward horizontal overcorrection has been observed in older women when taking a single step over an obstacle, and this was interpreted as a larger clearance margin to maintain safety (Berg and Blasi 2000). Extrapolating the interpretations described here for young adults with virtual obstacles to older adults with real obstacles, it is also possible that the backwards foot displacement of older women may be related to compromised ability to gather, store, and/or use obstacle information in the single step task. Second, subject 32 (Fig. 1) and another subject (not shown) had triangular shaped trail limb trajectories for the trail obstacle; the shape is similar to the trail limb trajectory of a 4-year-old girl with early bilateral lesion of the occipital cortex (Amicuzi et al. 2006). It was concluded that the lesion eliminated the detection of visual information that specified how to interact with the obstacle. This conclusion could be extended to the current findings that vision of the obstacle during approach specifies how to interact with the obstacle; while an obstacle memory may provide height information, it is not adequate for guiding the trail limb trajectory.
In summary, when an obstacle memory was not formed during the current approach, the control of the trajectory was impaired, ultimately resulting in a high failure rate. The failure rate was four times higher for the trail limb than the lead limb. Since the lead limb is visible in the lower periphery during crossing, vision of the limb, combined with stored height information, can be used to guide the lead limb more successfully than the trail limb. However, 9 % lead limb failure is relatively high, given that lead limb failures in young, healthy adults are rarely observed (Mohagheghi et al. 2004; Berard and Vallis 2006; Rietdyk and Rhea 2006; Rhea and Rietdyk 2007; Heijnen et al. 2012a). Action was impaired to a greater extent for the trail limb, which is likely due to the fact that the trail limb is not visible during crossing. These results emphasize that the dynamic visual input gained during approach is critical for success.
References
Alexander MS, Flodin BW, Marigold DS (2011) Prism adaptation and generalization during visually guided locomotor tasks. J Neurophysiol 106:860–871
Amicuzi I, Stortini M, Petrarca M et al (2006) Visual recognition and visually guided action after early bilateral lesion of occipital cortex: a behavioral study of a 4.6-year-old girl. Neurocase 12:263–279
Anson JG, Burgess R, Scott RL (2010) Vision and movement planning. In: Elliott D, Khan MA (eds) Vision and goal-directed movement: neurobehav perspect. Human Kinetics, Champaign, pp 59–78
Berard JR, Vallis LA (2006) Characteristics of single and double obstacle avoidance strategies: a comparison between adults and children. Exp Brain Res 175:21–31
Berg WP, Blasi ER (2000) Stepping performance during obstacle clearance in women: age differences and the association with lower extremity strength in older women. J Am Geriatr Soc 48:1414–1423
Binsted G, Heath M (2005) No evidence of a lower visual field specialization for visuomotor control. Exp Brain Res 162:89–94
Binsted G, Rolheiser TM, Chua R (2006) Decay in visuomotor representations during manual aiming. J Mot Behav 38:82–87
Chen HC, Ashton-Miller JA, Alexander NB, Schultz AB (1991) Stepping over obstacles: gait patterns of healthy young and old adults. J Gerontol 46:M196–M203
Drew T, Andujar J-E, Lajoie K, Yakovenko S (2008) Cortical mechanisms involved in visuomotor coordination during precision walking. Brain Res Rev 57:199–211
Elliott D (1988) The influence of visual target and limb information on manual aiming. Can J Psychol 42:57
Gibson JJ (1958) Visually controlled locomotion and visual orientation in animals. Br J Psychol 49:182–194
Gibson JJ (1966) The senses considered as perceptual systems. Houghton Mifflin, Boston
Heath M (2005) Role of limb and target vision in the online control of memory-guided reaches. Motor Control 9:281–311
Heath M, Neely K, Krigolson O, Binsted G, Elliott D (2010) Memory-guided reaching: what the visuomotor system knows and how long it knows it. In: Elliott D, Khan MA (eds) Vision and goal-directed movement: neurobehav perspect. Human Kinetics, Champaign, pp 79–96
Heijnen MJH, Muir BC, Rietdyk S (2012a) Factors leading to obstacle contact during adaptive locomotion. Exp Brain Res 223:219–231
Heijnen MJH, Muir BC, Rietdyk S (2012b) Interpolation techniques to reduce error in measurement of toe clearance during obstacle avoidance. J Biomech 45:196–198
Kelly VE, Janke AA, Shumway-Cook A (2010) Effects of instructed focus and task difficulty on concurrent walking and cognitive task performance in healthy young adults. Exp Brain Res 207:65–73
Lajoie K, Andujar J-É, Pearson K, Drew T (2010) Neurons in area 5 of the posterior parietal cortex in the cat contribute to interlimb coordination during visually guided locomotion: a role in working memory. J Neurophysiol 103:2234–2254
Lajoie K, Bloomfield LW, Nelson FJ, Suh JJ, Marigold DS (2012) The contribution of vision, proprioception, and efference copy in storing a neural representation for guiding trail leg trajectory over an obstacle. J Neurophysiol 107:2283–2293
Lee DN, Lishman JR, Thomson JA (1982) Regulation of gait in long jumping. J Exp Psychol Hum Percept Perform 8:448–459
Loverro KL, Mueske NM, Hamel KA (2013) Location of minimum foot clearance on the shoe and with respect to the obstacle changes with locomotor task. J Biomech 46:1842–1850
McVea DA, Pearson KG (2006) Long-lasting memories of obstacles guide leg movements in the walking cat. J Neurosci 26:1175–1178
McVea DA, Pearson KG (2007) Stepping of the forelegs over obstacles establishes long-lasting memories in cats. Curr Biol 17:R621–R623
Milner AD, Goodale MA (1995) The visual brain in action. Oxford University Press, New York
Milner A, Dijkerman H, McIntosh R, Rossetti Y, Pisella L (2003) Delayed reaching and grasping in patients with optic ataxia. Prog Brain Res 142:225–242
Mohagheghi AA, Moraes R, Patla AE (2004) The effects of distant and on-line visual information on the control of approach phase and step over an obstacle during locomotion. Exp Brain Res 155:459–468
Niang A, McFadyen B (2004) Adaptations in bilateral mechanical power patterns during obstacle avoidance reveal distinct control strategies for limb elevation versus limb progression. Mot Control 8:160
Patla AE (1998) How is human gait controlled by vision. Ecol Psychol 10:287–302
Patla AE, Greig M (2006) Any way you look at it, successful obstacle negotiation needs visually guided on-line foot placement regulation during the approach phase. Neurosci Lett 397:110–114
Patla A, Rietdyk S (1993) Visual control of limb trajectory over obstacles during locomotion: effect of obstacle height and width. Gait Posture 1:45–60
Patla AE, Vickers JN (1997) Where and when do we look as we approach and step over an obstacle in the travel path? NeuroReport 8:3661–3665
Patla AE, Rietdyk S, Martin C, Prentice S (1996) Locomotor patterns of the leading and the trailing limbs as solid and fragile obstacles are stepped over: some insights into the role of vision during locomotion. J Mot Behav 28:35–47
Rhea CK, Rietdyk S (2007) Visual exteroceptive information provided during obstacle crossing did not modify the lower limb trajectory. Neurosci Lett 418:60–65
Rhea CK, Rietdyk S (2011) Influence of an unexpected perturbation on adaptive gait behavior. Gait Posture 34:439–441
Rhea CK, Rietdyk S, Haddad JM (2010) Locomotor adaptation versus perceptual adaptation when stepping over an obstacle with a height illusion. PLoS ONE 5:e11544
Rietdyk S, Rhea CK (2006) Control of adaptive locomotion: effect of visual obstruction and visual cues in the environment. Exp Brain Res 169:272–278
Rietdyk S, Rhea CK (2011) The effect of the visual characteristics of obstacles on risk of tripping and gait parameters during locomotion. Ophthalmic Physiol Opt 31:302–310
Shinya M, Popescu A, Marchak C, Maraj B, Pearson K (2012) Enhancing memory of stair height by the motor experience of stepping. Exp Brain Res 223:405–414
Siu K-C, Woollacott MH (2007) Attentional demands of postural control: the ability to selectively allocate information-processing resources. Gait Posture 25:121–126
Telonio A, Blanchet S, Maganaris C, Baltzopoulos V, McFadyen B (2013) The detailed measurement of foot clearance by young adults during stair decent. J Biomech 46:1400–1402
Thies S, Jones R, Kenney L, Howard D, Baker R (2011) Effects of ramp negotiation, paving type and shoe sole geometry on toe clearance in young adults. J Biomech 44:2679–2684
Warren WH (2006) The dynamics of perception and action. Psychol Rev 113:358–389
Whishaw IQ, Sacrey LA, Gorny B (2009) Hind limb stepping over obstacles in the horse guided by place-object memory. Behav Brain Res 198:372–379
Winter DA (2009) Biomechanics and motor control of human movement. Wiley, Hoboken
Yang JF, Lam T, Pang MYC, Lamont E, Musselman K, Seinen E (2004) Infant stepping: a window to the behaviour of the human pattern generator for walking. Can J Physiol Pharmacol 82(8):662–674
Yogev-Seligmann G, Rotem-Galili Y, Mirelman A, Dickstein R, Giladi N, Hausdorff JM (2010) How does explicit prioritization alter walking during dual-task performance? Effects of age and sex on gait speed and variability. Phys Ther 90:177–186
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
Heijnen, M.J.H., Romine, N.L., Stumpf, D.M. et al. Memory-guided obstacle crossing: more failures were observed for the trail limb versus lead limb. Exp Brain Res 232, 2131–2142 (2014). https://doi.org/10.1007/s00221-014-3903-3
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00221-014-3903-3