
Abstract
It has been proposed in the past that adaptation to rotated visual feedback is based on directionally tuned modules. Here, we investigate whether adaptation depends on the number of modules that are concurrently activated. To disambiguate the number of modules and their spatial overlap, we decided to vary the number of target directions and their spacing independently. In light of recent work on the existence of fast and slow adaptive processes, we analyzed the role of target number and spacing separately for the first eight movements under rotated visual feedback and for later movements. We found that during the first eight movements, adaptation progressed three times faster when targets were spaced across a 42° rather than across a 360° range, irrespective of their number. During the subsequent movements, adaptation progressed 1.66 times faster with two than with eight targets, irrespective of their spacing. This differential dependence of early and late adaptation on target metrics confirms the existence of adaptive processes with different time scales and suggests that those processes differ not only by their clocking speed, but also by their functional properties. Specifically, the speed of fast processes seems to be constrained by the directional tuning width of adaptive modules, and the speed of slow processes by the number of presentations per target direction.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Several lines of evidence indicate that sensorimotor adaptation to rotated vision is directionally selective. Thus, adaptation established while pointing at targets in certain directions does not transfer freely to new directions; rather, transfer falls off with increasing difference between the new and the trained direction (Imamizu et al. 1995; Krakauer et al. 2000; Roby-Brami and Burnod 1995). When the starting point of hand movements is modified, adaptation is found to be anchored with respect to target direction rather than position (Wang and Sainburg 2005). Further support for directional selectivity comes from the finding that subjects can concurrently adapt to different rotations for different target directions, with no loss of adaptation speed or magnitude (Ghahramani and Wolpert 1997; Woolley et al. 2007). It has therefore been concluded that adaptation to rotated vision is based on directionally tuned adaptive modules (Roby-Brami and Burnod 1995; Tanaka et al. 2009).
Directional selectivity may not be the only notable characteristic of adaptation to rotated vision. Another critical factor could be the number of distinct target directions in which an adaptive module must handle. The earlier observation, that the adaptation rate decreases with an increasing number of target directions (Krakauer et al. 2000), would be compatible with such a view. However, that study co-varied the number of targets and the range of directions across which those targets were spaced and thus, does not allow an independent assessment of the role of target number. We therefore decided to vary these two metrics of the target array separately in the present study.
The time course of adaptive improvement is not well described by a single exponential function. It rather seems to follow multiple time scales, one in the order of a few trials and another in the order of several tens of trials (Krakauer et al. 2000; Smith et al. 2006; Snoddy 1926). The notion of multiple time scales and of cue-dependent modules has recently been combined in a model, which consists of a singular fast process, arranged in parallel with a slow process that contains several cue-dependent modules (Lee and Schweighofer 2009). We reasoned that the fast and slow process might depend differently on target metrics and therefore, decided to scrutinize the role of target number and range separately for the first few and for the remaining trials.
Methods
Subjects
Thirty female and 34 male volunteers participated after signing an informed consent statement. They were 23.2 ± 2.1 years old, right-handed, and had no prior experience with similar research. None of them reported sensorimotor disorders, and no such disorders were evident to the experimenters, except that some subjects wore corrective eyeglasses.
Apparatus
Subjects were seated on a 72-cm high table, bent forward, and looked vertically down. Looking through a mirror, they saw the virtual image of a 19′′ TFT monitor superposed on the table’s surface. The distance between their eyes and the virtual image was about 40 cm, depending on body height. Visual targets were displayed on the monitor under the control of a computer (CPU: Intel® Corel™2 Duo CPU E7500 2.93 GHz; GPU: Radeon X550 Series 256 MB; RAM: 1.96 GB). The display was synchronized with the monitor frame rate of 80 Hz using the “retrace” signal, which is generated at the end of each frame.
Due to the mirror and a shroud extending from the mirror to their neck, subjects were unable to see their hand as it moved across the table toward the perceived target locations. However, index fingertip position was registered by the Fastrak® motion analysis system with a resolution of 1 mm and 8.3 ms, respectively and was displayed as a cursor along with the targets, thus providing continuous visual feedback in the plane of finger movement.
Procedures
Subjects first saw a central starting point. After they had pointed at it with their invisible index fingertip for a randomized intervalFootnote 1 of 0.5–2 s, it was replaced by a target located on an imagined circle of 12 cm radius about the starting point. Target direction varied from trial to trial in a quasi-random sequence, which ensured that all targets appeared equally often within a cycle of eight targets. Two seconds later, or until subjects had pointed at the target for 0.5 s, it was replaced by the starting point. That point was again displayed until subjects had pointed at it for 0.5–2 s, the next target was then presented, etc.
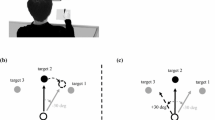
The experiment was subdivided into episodes of 30-s duration, separated by rest breaks of 5 s. Each session started with a baseline phase of four episodes with normal visual feedback. The subsequent adaptation phase consisted of 33 episodes in which feedback was distorted by a clockwise rotation about the starting point. Subjects were split into eight groups of equal size, differing with respect to the magnitude of rotation (30 or 60°), the number of target directions (2 or 8), and the range of directions across which the targets were spaced (42° arc or full 360° circle about the starting point). Specifically, the possible target directions were ±21° for some subjects, ±3, ±9, ±15, and ±21° for others, 0 and 180° for yet others, and 0, ±45, ±90, ±135, and 180° for the remaining subjects, where 0° denotes “from the starting point forwards, away from the subject”.
Data analysis
From the registered data, we determined the initial error of each response as the difference between target and hand direction 150 ms after movement onset,Footnote 2 i.e., before visual feedback could become effective. Initial errors of the first eight movements toward a target in the adaptation phase were considered to be indicative of fast adaptation processes.Footnote 3 They were normalized by subtracting the baseline and dividing by the rotation magnitude; this normalization rescaled the response size but preserved possible differences in the time course of errors. The outcome was then submitted to an analysis of variance (ANOVA) with the between-factors rotation magnitude, target number, and target spacing, as well as the within-factor movement (8 levels). For a second analysis, the mean initial errors of each adaptation episode were normalized with respect to the mean error of all subjects from the same group in the first adaptation episode; this normalization was introduced to remove effects already accounted for by the above ANOVA. The outcome was submitted to an ANOVA with the same between-factors and the within-factor episode (33 levels). The degrees of freedom were Greenhouse–Geisser-adjusted where necessary.
Results
Statistical analysis of the first eight movements yielded a significant effect of rotation (F(1,56) = 6.08, P < 0.05): the error was consistently smaller under the 30° rotation (0.62 ± 0.26) than under the 60° rotation (0.76 ± 0.26), which suggests that subjects compensated a larger proportion of the 30° than of the 60° perturbation. We further found highly significant effects of spacing (F(1,56) = 12.85, P < 0.001) and movement (F(7,392) = 4.27, P < 0.001): as illustrated in Fig. 1, adaptive improvement was substantially stronger with the 42° range than with the 360° range. Defining “improvement” as the difference between the first movement and the mean of the seventh and eighth movement, we yielded a score of 0.535 for the 42° range, but only of 0.181 for the 360° range. Thus, improvement was almost exactly three times faster with the narrow than with the wide target spacing. ANOVA of the first eight movements revealed no other significant effects, notably no main effect or interaction of number.

Time course of adaptation over the first eight movements of the adaptation phase in subjects tested with a 42° (triangles) and a 360° (circles) target spacing. Symbols represent across-subject means and error bars standard deviations of pointing errors normalized with respect to rotation magnitude
Statistical analysis of all adaptation episodes yielded significant effects of episode (F(32,1792) = 47.24, P < 0.001), episode × rotation (F(32,1792) = 2.85, P < 0.01), and episode × number (F(32,1792) = 2.64, P < 0.01). The effects of episode and episode*rotation are depicted in Fig. 2: the error decreased at a faster rate and toward a higher plateau with the 30° than with the 60° rotation. The effects of episode and episode*number are shown in Fig. 3: normalized error decreased at a faster rate with two than with eight targets. This difference of adaptation rates was quantified by an iterative-regression algorithm, which expanded the 2-target curve in time to minimize the root mean square difference between both curves. We thus yielded a scaling factor of 1.66, i.e., adaptation was 1.66 times faster with two than with eight targets. ANOVA of all adaptation episodes revealed no other significant effects, notably no main effect or interactions of spacing.

Time course of adaptation over all episodes of the adaptation phase in subjects tested with a 30° (filled symbols) and a 60° (open symbols) rotation of visual feedback. Symbols represent across-subject means and error bars standard deviations of pointing errors normalized with respect to the first adaptation episode

Time course of adaptation over all episodes of the adaptation phase in subjects tested with two (open symbols) and with eight (filled symbols) targets. Symbols represent across-subject means and error bars standard deviations of pointing errors normalized with respect to the first adaptation episode
To confirm that the reported relationship between adaptation rate and target number but not target spacing is not an artifact of our normalization procedure, we additionally calculated the adaptation rate from the non-normalized data of each group. To this end, we determined how many episodes each group needed to reduce the initial error below half of what it was during the first adaptation episode. We thus yielded a mean of 3.75 episodes for groups with two targets and of 7.75 episodes for groups with 8 targets; rearranging the groups, we yielded a mean of 5.75 episodes for those with a 360° target spread and also 5.75 episodes for those with a 42° spread. This outcome endorses the above dependence of the adaptation rate on target number but not target spacing.
Discussion
The primary goal of the present work was to scrutinize whether the speed of adaptation to rotated feedback depends on the number of target directions or rather on the range across which targets are spread. Our data suggest that both metrics play a role. An analysis focusing on the first few movements revealed a strong dependence on spacing, but not on the number. In contrast, an analysis of the remaining movements yielded a marked dependence on the number, but not on spacing. This differential dependence of early and late adaptation on target metrics supports the view that the sensorimotor system utilizes multiple adaptive processes with different time scales (Krakauer et al. 2000; Smith et al. 2006; Snoddy 1926). Moreover, it documents that those processes differ not only with respect to their clocking speed, but also with respect to their functional characteristics.
Our observation, that fast processes depend on the spacing but not on the number of target directions, suggests that they rely on modules with limited directional tuning curves: when targets are spaced across a wide range, they activate more modules but each of them less often, thus reducing adaptive improvement. Since early adaptation with 360° spacing was only one-third of that with 42° spacing, we posit that the wider spacing activated three times as many modules than the narrower spacing. Assuming a parsimonious system that does not employ more modules than necessary, we then have:
and
where k is the number, and w the tuning width of the presumed modules. Equations 1–4 are only satisfied for k = 1 and 120° ≤ w < 180°, which suggests that the tuning curves of adaptive modules are fairly wide, corresponding roughly to those of single cells found in the motor cortex (Georgopoulos et al. 1988), parietal area 5 (Kalaska et al. 1983), parietal area 7 (Andersen et al. 1990), and the superior colliculi (Ottes et al. 1986).
Slow adaptive processes seem to be directionally tuned as well, since transfer to unpracticed targets at the end of the adaptation phase is limited to neighboring directions. The tuning width can be estimated from published graphs as about 150° (Imamizu et al. 1995), 80° (Roby-Brami and Burnod 1995), or 60° (Krakauer et al. 2000). A modeling approach predicted the latter dataset well with a half-width of 23°, which corresponds to a tuning width of 46°, but the authors cautioned that their model is not very sensitive to the actual parameter values (Tanaka et al. 2009). Thus, summing up the tuning curves of slow adaptive processes may be between 50 and 150° wide, which is similar to the width determined above for fast processes. However, the directional tuning of slow processes—unlike that of fast processes—seems not to constrain adaptation speed.
If slow processes were constrained by tuning curves of 50–150° width, adaptation with two or eight targets spaced across 42° should be faster than that with two or eight targets spaced across 360°. Our data provide two lines of evidence that this was not the case. First, ANOVA did not yield the predicted effect of spacing. Second, our time-scaling algorithm (see "Results") revealed that adaptation was fastest with two targets spaced across 42° and slowed down for the other target arrays by a factor of 1.45 (two targets, 360°), 1.61 (eight targets, 360°), or 2.91 (eight targets, 42°); thus, adaptation with the latter target array was slowest—contrary to the above prediction. Since the only reliable ANOVA effect was that of number, we conclude that the rate of slow adaptive processes is mainly constrained by the number of trained target directions, probably because an increase in this number implies fewer presentations per target.
In addition to the above findings, our study also revealed differences between adaptation to a visual rotation of 30° and to a rotation of 60°. Even after normalizing for rotation magnitude, the error with a 30° rotation was initially smaller, decreased at a higher speed, and reached a higher plateau than the error with a 60° rotation. This finding is not in conflict with the hypothesis that adaptation to different rotations is achieved by common processes (Abeele and Bock 2001; Bock et al. 2003), since those processes may depend on rotation magnitude in a non-linear fashion. It would be desirable to probe the presumed non-linearity in experiments, which compare adaptation to more than two rotation magnitudes.
Notes
Randomization was performed with the in C++ statement “rand” after seeding with “srand”.
Movement onset was defined by a velocity threshold of 30 mm/s.
We decided for eight movements to represent fast adaptive processes since all target directions were presented equally often within eight movements (see "Procedures"). This is slightly less than the ten or twelve movements selected in earlier studies on adaptation to mirror reversals (Stratton et al. 2007) and force fields (Smith et al. 2006), respectively. Since the first eight adaptation movements in the present study took about 30 s, our choice is comparable to the value determined for fast adaptation to visual rotations in psychophysical experiments (Inoue et al. 2000). We are therefore confident that our analysis didn’t exceed the temporal limits of fast processes.
References
Abeele S, Bock O (2001) Mechanisms for sensorimotor adaptation to rotated visual input. Exp Brain Res 139:248–253
Andersen RA, Bracewell RM, Barash S, Gnadt JW, Fogassi L (1990) Eye position effects on visual, memory, and saccade-related activity in areas LIP and 7a of macaque. J Neurosci 10:1176–1196
Bock O, Abeele S, Eversheim U (2003) Human adaptation to rotated vision: interplay of a continuous and a discrete process. Exp Brain Res 152:528–532
Georgopoulos AP, Kettner RE, Schwartz AB (1988) Primate motor cortex and free arm movements to visual targets in three-dimensional space. II. Coding of the direction of movement by a neuronal population. J Neurosci 8:2928–2937
Ghahramani Z, Wolpert D (1997) Modular decomposition in visuomotor learning. Nature 386:392–395
Imamizu H, Uno Y, Kawato M (1995) Internal representations of the motor apparatus: implications from generalization n visuomotor learning. J Exp Psychol Hum Percept Perform 21:1174–1198
Inoue K, Kawashima R, Satoh K, Kinomura S, Sugiura M, Goto R, Ito M, Fukuda H (2000) A PET study of visuomotor learning under optical rotation. NeuroImage 11:505–516
Kalaska JF, Caminiti R, Georgopoulos AP (1983) Cortical mechanisms related to the direction of two-dimensional arm movements: relations in parietal area 5 and comparison with motor cortex. Exp Brain Res 51:247–260
Krakauer J, Pine Z, Ghilardi M-F, Ghez C (2000) Learning of visuomotor transformations for vectorial planning of reaching trajectories. J Neurosci 20:8916–8924
Lee J, Schweighofer N (2009) Dual adaptation supports a parallel architecture of motor memory. J Neurosci 29:10396–10404
Ottes F, Van Gibergen J, Eggermont J (1986) Visuomotor fields of the superior colliculus: a quantitative model. Vision Res 26:857–873
Roby-Brami A, Burnod Y (1995) Learning a new visuomotor transformation: error correction and generalization. Cogn Brain Res 2:229–242
Smith M, Ghazizadeh A, Shadmehr R (2006) Interacting adaptive processes with different timescales underlie short-term motor learning. PLoS Biol 4:1035–1043
Snoddy G (1926) Learning and stability. J Appl Psychol 10:1–36
Stratton S, Liu Y-T, Hong S, Mayer-Kress G, Newell K (2007) Snoddy (1926) revisited: time scales of motor learning. J Motor Behav 39:503–515
Tanaka H, Sejnowski T, Krakauer J (2009) Adaptation to visuomotor rotation through interaction between posterior parietal and motor cortical areas. J Neurophysiol 102:2921–2932
Wang J, Sainburg R (2005) Adaptation to visuomotor rotations remaps movement vectors, not final positions. J Neurosci 25:4024–4030
Woolley D, Tresilian J, Carson R, Rick S (2007) Dual adaptation to two opposing visuomotor rotations when each is associated with different regions of workspace. Exp Brain Res 179:155–165
Acknowledgments
This work was supported by the German Space Agency DLR on behalf of the Bundesministeriu für Wirtschaft und Technologie (Grant 50WB0625). Thanks are due to Dipl.-Ing. (FH) L. Geisen for software development and to V. Rohde, K. Friederichs, and M. Hinrichs for their help with data collection and analysis.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Bock, O., Schmitz, G. Adaptation to rotated visual feedback depends on the number and spread of target directions. Exp Brain Res 209, 409–413 (2011). https://doi.org/10.1007/s00221-011-2564-8
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00221-011-2564-8