
Abstract
We required healthy subjects to recognize visually presented one’s own or others’ hands in egocentric or allocentric perspective. Both right- and left-handers were faster in recognizing dominant hands in egocentric perspective and others’ non-dominant hand in allocentric perspective. These findings demonstrated that body-specific information contributes to sense of ownership, and that the “peri-dominant-hand space” is the preferred reference frame to distinguish self from not-self body parts.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
In recent years, recognition of body and body parts has been demonstrated to recruit devoted neural circuits, in which the Extrastriate Body Area (EBA) in the lateral occipitotemporal cortex plays a pivotal role (for a review see Peelen and Downing 2007). Neural activity in such circuits appears to be modulated by visual perspective and handedness. On one hand, the right EBA is more strongly activated by the observation of body parts (e.g., hands) presented from an allocentric viewpoint (i.e., as if looking at someone else’s body) than in egocentric perspective (i.e., as if looking at one’s own body; Chan et al. 2004; Saxe et al. 2006). This enhanced response to allocentric perspective suggests the involvement of EBA in perceiving others’ body (Peelen and Downing 2007), whereas recognition of one’s own body seems to require the contribution of a network including parietal and frontal areas (Hodzic et al. 2009a, b).
On the other hand, left- and right-handers can show opposite lateralization in the activity of body-specific visual areas (Willems et al. 2010b). This finding adds to those demonstrating that handedness affects neural correlates of action observation (Willems and Hagoort 2009), motor imagery (Willems et al. 2009), and mental representation of action verbs (Willems et al. 2010a). Taken together, these convergent lines of evidence support the so-called body-specificity hypothesis according to which right-handed and left-handed individuals have different mental representations of action- and body-related information, due to the different way in which they interact with the environment (Casasanto 2009).
On the basis of the data reviewed above, we aimed at verifying whether handedness and visual perspective can modulate visual discrimination of self and not-self body parts on behavioral grounds. Until now, only a few studies have directly investigated the ability to discriminate self from others’ body parts, most of them addressing the neural correlates of self face recognition (for a review, Gillihan and Farah 2005). In the present paper, we required right-handed and left-handed healthy subjects to explicitly recognize their own or others’ hands presented according to the egocentric or the allocentric perspective.
The body-specificity hypothesis would predict that handedness affects self/not-self discrimination. More precisely, this hypothesis would foresee right- and left-handers to show opposite advantages in recognizing hands, with the left hand preferred in left-handers and the right hand in right-handers in egocentric perspective. Moreover, the present experimental paradigm will allow to verify whether body-specificity can affect others’ body parts processing too. Actually, allocentric perspective is better consistent with looking at someone else’s body; if body-specificity applies to such a condition, one could expect that handedness also affects others’ body part recognition, with an asymmetry in recognizing right or left others’ hand. Alternatively, both left- and right-handed subjects might show an advantage in the recognition of other’s right hand; such a pattern would not be compatible with an extension of body-specificity to recognition of others’ body parts but rather with a simple effect of visual familiarity (because most people are right-handed others’ right hand is most often attended to in common social interactions).
Materials and methods
Subjects
Fifty healthy male volunteers, aged 18–30 years (mean age = 22.6, SD = ±3.2), were screened for the study. To the purpose of selecting subjects with definite hand preference, we enrolled only subjects with scores on the Edinburgh Handedness Inventory (EHI, Oldfield 1971) ranging from 80 to 100; we excluded subjects wearing rings or having easily recognizable marks (e.g., tattoos, scars). On this basis, we selected 20 right-handed subjects (EHI score = 88.6 ± 6.2) and 16 left-handers (EHI score = −86.5 ± 5.9). All participants were unaware of purposes and predictions of the experiment at the time of testing. The study was conducted in accordance with the ethical standards of the 1964 Declaration of Helsinki, and an informed consent was obtained from all participants.
Stimuli and procedure
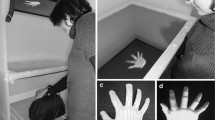
Stimuli were full-color photos of left and right hands of participating subjects, viewed from back in a neutral, frontal, upright posture against a non-reflecting black background. When subjects were screened for handedness, pictures were taken by means of a digital camera (5.2 million effective pixels) kept at a constant distance (40 cm) from the target. The original images of the hands (one picture per hand) in upright orientation (0°, consistent with the perspective of the viewer, i.e., egocentric) were digitally manipulated to obtain the hand images in opposite orientation (180°, consistent with looking at someone else’s body, i.e., allocentric). The hand images were large approximately 9.5 cm along the widest axis (10.7° of visual angle at a viewing distance of 50 cm).
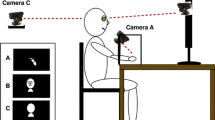
During the experiment, each subject was shown the four pictures of his own hands plus 8 pictures of others’ hands from two different subjects, not involved in the experiment and whose hands did not present salient features. The set of 12 pictures was repeatedly presented for a total of 240 trials (80 self hand trials and 160 others’ hand trials); hand laterality (left or right), hand ownership (self or not-self), and visual perspective (egocentric or allocentric) were randomised across trials. In each trial, a fixation point (800 ms) was followed by a centrally presented hand stimulus until a response was emitted. Participants were required to decide whether they were presented with their own or others’ hands and to press one of two keys on a foot pedal. Response key was counterbalanced across participants, with half responding with their right foot to “my hand” and with their left foot to “others’ hand”, and the other half with the opposite mapping.
Participants were sitting 50 cm from a computer screen with their head on a chin rest and their hands placed palm-down on their thighs, out of sight. Subjects were instructed to refrain from moving their head and fingers, and the experimenter (seated behind participants) checked that subjects complied with this instruction for the whole task. Reaction times (RTs) and accuracy were recorded.
Results
Overall accuracy was high (93%; see Table 1 for details on accuracy data) and showed a significant negative correlation with overall RTs (877.91 ms; Pearson’s correlation: r = –.577, P = .0001). This finding ruled out a trade-off between speed and accuracy, thus allowing to perform statistical analyses on RTs data only.
A 2 × 2 × 2 × 2 mixed ANOVA was conducted on correct RTs, with hand ownership (self, not-self), hand laterality (left, right), and visual perspective (egocentric, allocentric) as within-subject factors and with group (right-handers, left-handers) as between-subject factor.
Results (see Supplementary table for a detailed presentation of ANOVA results) showed significant first-order interactions between hand laterality and group [F(1,19) = 5.823, P = .021, η2 = .146], and between hand ownership and visual perspective [F(1,34) = 26.404, P = .0001, η2 = .437]; second-order interactions among hand ownership, hand laterality and group [F(1,34) = 5.144, P = .030, η2 = .131], and among hand laterality, visual perspective and group [F(1,34) = 16.259, P = .0001, η2 = .324] were also significant. More relevant here, the third-order interaction among hand ownership, hand laterality, visual perspective and group was statistically significant [F(1,34) = 13.915, P = .001, η2 = .290]. No main effects and no other interactions were statistically significant.
Post hoc comparisons (paired t-tests) performed on the hand laterality × group interaction showed in right-handers a not significant advantage in recognizing right (859.34) with respect to left-hand stimuli (882.46; t = −1.801, P = .088), and in left-handers a not significant advantage in recognizing left (877.19) versus right-hand stimuli (892.65; t = −1.979, P = .067).
Post hoc comparisons performed on the hand ownership × visual perspective interaction showed that recognition of self hands was faster in egocentric (854.14) than in allocentric perspective (917.31; t = −4.773, P = .0001), whereas recognition of not-self hands was faster in allocentric (851.11) than in egocentric perspective (889.11; t = 2.942, P = .006).
Post hoc comparisons performed on the hand ownership × hand laterality × group interaction showed that right-handers were faster in recognizing right (861.51) than left self hand (901.95; t = −2.244, P = .037), without differences between right (857.17) or left (862.96) not-self hand (t = −.305, P = .764). Left-handed subjects were significantly faster in recognizing left (870.62) than right (908.82) self hand, without differences between left (883.76) or right (876.47) not-self hand (t = −.611, P = .550).
Post hoc comparisons performed on the hand laterality × visual perspective × group interaction showed that right-handers were significantly faster in recognizing right-hand stimuli in egocentric (837.85) than in allocentric (880.83) perspective (t = −2.549, P = .020), without differences in recognizing left-hand stimuli in allocentric (865.44) or egocentric (899.47) perspective (t = 1.601, P = .126). Left-handed subjects recognized left-hand stimuli faster in egocentric (847.66) than in allocentric (906.72) perspective (t = −3.488, P = .003), without differences in recognizing right-hand stimuli in allocentric (883.84) or egocentric (901.45) perspective (t = 1.246, P = .232).
Most importantly, post hoc comparisons on the hand ownership × hand laterality × visual perspective × group interaction (Fig. 1) showed that in egocentric perspective right-handed subjects were significantly faster in recognizing right than left self hand (t = −2.230, P = .038), whereas the difference between recognition of right or left not-self hand fell short of significance level (t = −2.004, P = .060). When dealing with stimuli in allocentric perspective, right-handers were significantly faster in recognizing left than right others’ hand (t = 2.999, P = .007), whereas there were not significant differences between left and right self hand recognition (t = −1.272, P = .219). Left-handers were significantly faster in recognizing left than right self hand in egocentric perspective (t = 2.788, P = .014), with a marginally significant difference between recognition of left or right others’ hand (t = 2.106, P = .058). When dealing with stimuli in allocentric perspective, left-handers were significantly faster in recognizing right than left others’ hand (t = −3.314, P = .005), without significant differences between left or right self hand (t = 1.230, P = .238).

Mean RTs (bars represent standard errors) to hand stimuli are plotted by hand ownership (self, not-self), hand laterality (right hand, left hand), visual perspective (egocentric, allocentric) and group (right-handers, left-handers). * Significant at P < .05
Discussion
The present study demonstrates that explicit visual recognition of both one’s own and others’ hands fits the body-specificity hypothesis. Actually, hand dominance significantly modulated the effect of visual perspective on task performance, as revealed by the high-order interaction among hand ownership, hand laterality, visual perspective and group. In synthesis, in egocentric perspective, both right- and left-handers were faster in recognizing dominant hands, with a stronger advantage when dealing with self than with not-self hands, whereas in allocentric perspective, they were faster in identifying not-self non-dominant hands (i.e., others’ left hands in right-handers and others’ right hands in left-handers).
The influence of handedness on recognition of others’ body parts respected a specular rather than an anatomic principle, since recognition of others’ hands in allocentric perspective was advantaged for the (non-dominant) hand, which is represented in the side specular to one’s own dominant hand. The phenomenon of specular correspondence has not been previously investigated in recognition of self versus not-self static body parts, whereas it has been repeatedly reported in studies on imitation and action observation (Alaerts et al. 2009; Bekkering et al. 2000; Gleissner et al. 2000; Koski et al. 2003). For instance, in a recent TMS study, Alaerts et al. (2009) demonstrated that the mirror matching also manifests (with higher responses of the motor cortex) during mere observation of specular with respect to anatomically correspondent hand actions. Behavioral studies suggest that specular correspondence applies to mental simulation of actions too. Conson et al. (2009) required right-handed subjects to judge laterality of hand stimuli preceded by digits cuing attention to the left or the right side of space. Subjects were faster in recognizing egocentric right hand and allocentric left-hand stimuli when preceded by a cue priming the right side of egocentric space, whereas they were faster in recognizing egocentric left hand and allocentric right-hand stimuli preceded by a pre-activation of the left side of egocentric space. Such results showed that subjects mentally align their body midline with the midline of the imagined body of other (Ottoboni et al. 2005), with “my” right hand and “your” left hand both sharing the right side of “my” egocentric space.
The present findings demonstrated that specular correspondence is not limited to motor-related phenomena but also applies to visual recognition of body parts. It is possible to hypothesize that handedness defines the reference frame used for the egocentric body representation contributing to the sense of ownership of body parts; this frame is centered on one’s own dominant hand and extends to the surrounding space in which others’ body is represented in allocentric perspective. Such a hypothesis would explain the advantage for the recognition of one’s own dominant hand and of others’ hand that lies on the side facing one’s own dominant hand.
The existence of specialized brain systems that represent the sector of space closely surrounding the hand (i.e., peri-hand space representation) has been demonstrated in both monkeys (Graziano 1999) and humans (Ladavas et al. 2000; Farnè et al. 2000). For instance, in a study on patients with lesions to the right parietal cortex, Farnè et al. (2000) showed that tactile stimuli on the dorsal surface of the left hand could be extinguished by a visual stimulus above patients’ own right hand, or above a rubber hand presented in egocentric perspective; when the rubber hand was rotated to make it visually inconsistent with the subject’s own perspective (allocentric perspective), no cross-modal extinction could be observed. These results indicate that the fronto-parietal visuo-tactile peri-hand representation system codes one’s own body parts according to an egocentric perspective, in line with neuroimaging evidence (Chan et al. 2004; Saxe et al. 2006).
It has been claimed that the visuo-tactile hand-centered coordinate system allows the brain to attribute body parts to the self (Makin et al. 2008). We found here that the egocentric representation of one’s own “peri-dominant-hand space” is the reference frame according to which one can distinguish between one’s own and others’ body parts. Such behavioral findings would imply that visual perception is body-specific, namely that body-specific patterns of motor experience shape not only how we act upon the world, but also how we perceive it. In the context of the relationships between action and perception, a substantial amount of evidence has demonstrated that motor-related processes feed visual perception, but these data mainly derived from studies investigating action observation (for a review see Wilson and Knoblich 2005). Our study, instead, used static body parts not implying motion, and thus our results extended the role of the body-related specificity of the action system (e.g., hand dominance) into the domain of visual discrimination between self and not-self body parts. This may reflect the neurofunctional interplay between (fronto-parietal) areas representing one’s own hand and those (EBA) involved in others’ body representation, but neuroimaging studies employing the present behavioral paradigm are necessary to define the neural mechanisms subserving the modulation of body-specificity on the sense of ownership.
References
Alaerts K, Heremans E, Swinnen SP, Wenderoth N (2009) How are observed actions mapped to the observer’s motor system? Influence of posture and perspective. Neuropsychologia 47:415–422
Bekkering H, Wohlschlager A, Gattis M (2000) Imitation of gestures in children is goal-directed. Q J Exp Psychol A 53:153–164
Casasanto D (2009) Embodiment of abstract concepts: good and bad in right- and left-handers. J Exp Psychol Gen 138:351–367
Chan AW, Peelen MV, Downing PE (2004) The effect of viewpoint on body representation in the extrastriate body area. Neuroreport 15:2407–2410
Conson M, Mazzarella E, Trojano L (2009) Numbers are represented in egocentric space: effects of numerical cues and spatial reference frames on hand laterality judgements. Neurosci Lett 452:176–180
Farnè A, Pavani F, Meneghello F, Ladavas E (2000) Left tactile extinction following visual stimulation of a rubber hand. Brain 123:2350–2360
Gillihan SJ, Farah MJ (2005) Is self special? A critical review of evidence from experimental psychology and cognitive neuroscience. Psychol Bull 131:76–97
Gleissner B, Meltzoff AN, Bekkering H (2000) Children’s coding of human action: cognitive factors influencing imitation in 3-year-olds. Develop Sci 3:405–414
Graziano MS (1999) Where is my arm? The relative role of vision and proprioception in the neuronal representation of limb position. Proc Natl Acad Sci USA 96:10418–10421
Hodzic A, Muckli L, Singer W, Stirn A (2009a) Cortical responses to self and others. Hum Brain Mapp 30:951–962
Hodzic A, Kaas A, Muckli L, Stirn A, Singer W (2009b) Distinct cortical networks for the detection and identification of human body. Neuroimage 45:1264–1271
Koski L, Iacoboni M, Dubeau MC, Woods RP, Mazziotta JC (2003) Modulation of cortical activity during different imitative behaviors. J Neurophysiol 89:460–471
Ladavas E, Farnè A, Zeloni G, di Pellgrino G (2000) Seeing or not seeing where your hands are. Exp Brain Res 131:458–467
Makin TR, Holmes NP, Ehrsson HH (2008) On the other hand: dummy hands and peripersonal space. Behav Brain Res 191:1–10
Oldfield RC (1971) The assessment and analysis of handedness: the Edinburgh inventory. Neuropsychologia 9:97–113
Ottoboni G, Tessari A, Cubelli R, Umiltà C (2005) Is handedness recognition automatic? A study using a Simon-like paradigm. J Exp Psychol Hum Percept Perform 31:778–789
Peelen MV, Downing PE (2007) The neural basis of visual body perception. Nat Rev Neurosci 8:636–648
Saxe R, Jamal N, Powell L (2006) My body or yours? The effect of visual perspective on cortical body representations. Cereb Cortex 16:178–182
Willems RM, Hagoort P (2009) Hand preference influences neural correlates of action observation. Brain Res 1269:90–104
Willems RM, Toni I, Hagoort P, Casasanto D (2009) Body-specific motor imagery of hand actions: neural evidence from right- and left-handers. Front Hum Neurosci 3:39
Willems RM, Hagoort P, Casasanto D (2010a) Body-specific representations of action verbs: neural evidence from right- and left-handers. Psychol Sci 21:67–74
Willems RM, Peelen MV, Hagoort P (2010b) Cerebral lateralization of face-selective and body-selective visual areas depends on handedness. Cereb Cortex 20:1719–1725
Wilson M, Knoblich G (2005) The case for motor involvement in perceiving conspecifics. Psychol Bull 131:460–473
Acknowledgments
We are grateful to Ida De Simone for her help in collecting data.
Author information
Authors and Affiliations
Corresponding authors
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
About this article
Cite this article
Conson, M., Aromino, A.R. & Trojano, L. Whose hand is this? Handedness and visual perspective modulate self/other discrimination. Exp Brain Res 206, 449–453 (2010). https://doi.org/10.1007/s00221-010-2418-9
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00221-010-2418-9