
Abstract
We investigated the possible consequences of two consecutive postural tasks on adaptation. Four groups (total number of 46 healthy subjects) were perturbed on two consecutive days with vibration stimulus to tibialis anterior or posterior calf muscles, or both in different orders. Postural movements were recorded with a force platform. There were three major results: (1) tibialis anterior vibration instigated postural adaptation during exposure to the vibration, but did not induce long-term adaptation from day to day, contrary to posterior calf vibration. (2) The long-term postural adaptation from day to day when the posterior calf was vibrated was not affected by prior or subsequent tibialis anterior vibration, which contrasts to other studies on motor learning. (3) Exposure to posterior calf vibration prior tibialis anterior vibration, led to changes of postural strategies and larger amount of torque variance, implying that postural strategies initiated by the gastrocnemius vibration were re-employed during the subsequent tibialis anterior stimulation. This may represent the formation of an internal model, used as feed-forward control of posture, possibly consisting of sensory reweighting. Postural perturbations need to be sufficiently difficult to withstand, in order to induce long-term learning, and postural strategies may be transferred between different postural challenges if they post different demands. Clinically, this suggests that exercises designed to rehabilitate patients should be sufficiently challenging to instigate learning processes, and spaced in order to avoid development of inappropriate postural strategies.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
The ability of the postural control system to adapt and learn to resolve sensory conflicts is essential for the human capacity to undertake the wide range of activities that constitute normal daily life. It is also crucial in recuperating and rehabilitating from losses or decline of the sensory systems involved in maintaining postural control. Patients are generally rehabilitated by a set of exercises, designed to enhance the function of the remaining function in order to procure a balance close to normal. The adaptive mechanisms are generally held to be due to sensory reweighting, i.e. an increased reliance on the remaining functioning sensory systems (Oie et al. 2002; Fransson et al. 2003).
The ability to maintain upright stance is complex and uses both feedback (Johansson et al. 1988) and feed-forward mechanisms (Frank and Earl 1990). Feedback control depends on sensory input from the vestibular system (inner ear), vision and the somatosensory system (proprioception—sensors in joints and muscles, and pressor receptors of the soles of the feet). Information from these afferent sources are processed, integrated and weighted to their relative importance and context in the central nervous system (CNS) (Peterka 2002). Feed-forward mechanisms are based on experience of planning a movement or preparing to withstand a perturbation before it actually takes place (Wolpert et al. 1995).
The postural control system can be manipulated by solitary perturbations of the different sensory systems, and the method selected in this study, vibratory perturbation to postural muscles, increases the firing of the muscle spindles, thus signaling that the muscle is being stretched (Matthews 1986). The vibration induces a reflexive contraction (tonic vibratory reflex) of the affected muscle as well as an illusion of body movement that yields increased body sway (Goodwin et al. 1972). Postural adaptation to daily repeated vibratory perturbation to the calf muscles show two in time separated processes (Tjernstrom et al. 2002). There is an immediate reduction of evoked body sway during the active vibratory perturbation and further reductions of responses develop over the days. These fast and more slow processes of learning postural control mimic how motor memories generally are formed; through consolidation of short-lasting into longer-lasting memories (Shadmehr and Brashers-Krug 1997). The concept of a consolidation process in motor memory formation is that the learnt skill is processed and modified within the CNS, while not executing the skill itself (Shadmehr and Brashers-Krug 1997). Sleep is generally regarded as beneficial to the consolidation processes (Shen and McNaughton 1996), and it has been shown that neuronal activities from preceding experience (or learning) are re-expressed in the hippocampus during slow-wave sleep (Wilson and McNaughton 1994). The consolidation process that takes place between training sessions has been shown to be time-dependent (McGaugh 1966; Abel and Lattal 2001) and can be disrupted by learning a second task immediately before and up to 3 h after the first (Brashers-Krug et al. 1996; Shadmehr and Brashers-Krug 1997). The mechanism for consolidation is not fully understood, but is considered to involve both intrinsic and extrinsic synaptic function, such as synaptic connections and long-term potentiation and/or depression of synapses (Sanes 2003).
Present aim was to investigate how postural control is maintained when individuals are subjected to repeated perturbation caused by vibratory stimulus to different muscles of the lower leg in different orders, i.e. vibration to tibialis anterior and/or posterior calf muscles. The hypothesis being that there could be differences in adaptation and development of postural strategies in response to the different types of perturbations and that the development of learned responses could be influenced by other coinciding perturbations.
Materials and methods
Subjects
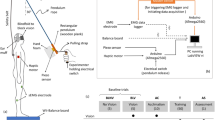
Forty-six healthy volunteers were recruited and divided into four groups (Fig. 1). Group ‘Calf only’ (n = 12, 7 male, 5 female, mean age 27.0, range 19–38 years) had posterior calf vibration only. Group ‘Tib only’ (n = 11, 5 male, 6 female, mean age 27.3, range 23–44 years) had tibialis anterior vibration only. Group ‘Calf-Tib’ labeled (n = 12, 5 male, 7 female, mean age 27.3, range 20–38) was exposed to posterior calf vibration prior to tibialis anterior vibration. Group ‘Tib-Calf’ (n = 11, 7 male, 4 female, mean age 23.3, range 20–27 years) was exposed to tibialis anterior vibration prior to posterior calf vibration. The subjects did not use any medication, except oral contraceptives or had experienced any otoneurological, neurological, psychiatric, orthopedic or hearing disorder. Informed written consent was obtained and the study was conducted in accordance with the Helsinki Declaration of 1975 and approved by the local ethical committee.

Table of group division and the order and application of the vibratory stimulation. Analyses were performed to study changes within each of the vibration tests, and between the same tests on the consecutive days. In addition a GLM ANOVA was used to investigate the order effects of where the vibration was applied
Method
Postural control was evaluated by a force platform while perturbing stance. Forces and torques actuated by the feet were recorded with six degrees of freedom by a custom-made force platform developed at the Department of Solid Mechanics, Lund Institute of Technology. Data were sampled at 50 Hz with a 12-bit AD converter. The vibrations were delivered by two custom-made cylindrical vibrators (0.06 m long and 0.01 m in diameters), and held in place with an elastic strap around each leg. Special care was taken to have the same strap tension for both legs and according to the standard in all tested subjects. The vibrators were placed either over the Gastrocnemius muscle or over the tibialis anterior muscle of both legs orientated with the long axis in the muscle-fiber direction. The vibration amplitude was 1.0 mm amplitude at a constant frequency of 85 Hz. The vibratory stimulation was executed according to a computer controlled pseudorandom binary sequence (PRBS) schedule (Johansson 1993) for 200 s by turning on/off the vibratory stimulation. The PRBS schedule was composed of stimulation shift periods limited to multiples of 0.8 s and the sequence was designed to contain a balanced variation of vibration pulses of duration from 0.8 up to 6.4 s long (yielding an effective bandwidth of 0.1–2.5 Hz). The spectral bandwidth of the stimulus was determined by FFT analysis. Thus, the designated PRBS stimuli covered a broad power spectrum and the randomized stimulation reduced the opportunity to make anticipative and pre-emptive adjustments.
Procedure
After a brief information the subjects were instructed to stand erect but not at attention, with their eyes closed and arms crossed over the chest and feet at an angle of about 30° open to the front and the heals approximately 3 cm apart. For group ‘Calf-Tib’ and ‘Tib-Calf’ two tests were performed after another with a 5-min rest period in between. The same test order was maintained on the second day (Fig. 1).
Data analysis
The variance of torque was calculated for five periods for each test: the quiet stance period (0–30 s) before stimulation was applied, and from four periods (1–4) during the stimulation (30–80, 80–130, 130–180, and 180–230 s, respectively). The data were normalized by squared mass and squared height since regression analysis showed dependence on those factors.
Postural stability while standing is commonly analyzed using force platforms and the movements of the center of pressure (CoP), i.e. the point of application of the ground reaction force. We measured torque and analyzed the variance of the torque values. Torque correspond to CoP; torque τ is calculated from the formula τ = CoP · Fz; where Fz ≈ m · g; where m the assessed subjects mass (in kg) and g gravitational constant 9.81 (in m/s2). Hence, changes in recorded torque are equivalent to changes in CoP (Patel et al. 2008). Changes in recorded torque from the force platform correspond well to the actual body movements and posture changes induced by vibratory stimulus (Fransson et al. 2007). The formula for variance is given by;
i sample, n number of samples recorded during an analyzed period.
One benefit with presenting torque variance values is that the calculated value corresponds directly with the energy used toward the support surface to maintain stability (Magnusson et al. 1990; Johansson and Magnusson 1991). As stimulation to calf muscles predominantly cause anteroposterior sway (Eklund 1973), only those responses were analyzed.
Non-parametric statistics were used since some values were not normally distributed and normal distribution could not be obtained after log-transformation. Statistics was calculated with non-parametric Wilcoxon for paired data (groups) and Mann–Whitney for unpaired data (between the groups). Comparisons according to Wilcoxon were made between the rest period and the stimulation period 1, and between stimulation period 1 and 4 of each test. The different periods were also analyzed between the consecutive days. Between the different groups were the differences in each period of either calf or tibialis anterior vibration analyzed according to Mann–Whitney. The statistics were corrected according to Bonferroni (Altman 1991), in the case of Wilcoxon with a factor of 3, and Mann–Whitney, a factor of 2, yielding P < 0.0167 and <0.025, respectively, that were considered to be statistically significant. This was performed since the same data in Wilcoxon were used three times and in Mann–Whitney two times.
In addition, the effects of order of where the vibration was applied was analyzed using a general linear model univariate analysis of variance (GLM univariate ANOVA) test on log-transformed values (Altman 1991). The accuracy of the GLM model was evaluated by testing whether the GLM model residuals, the model residuals provided by the statistical software (SPSS ver 17), were distributed normally. Analyses showed that normal GLM model residuals were better ensured if the torque variance data analyzed by GLM model data was log-transformed prior to the analysis. This procedure was subsequently used in all GLM analyses. In the GLM analysis, P < 0.05 were considered statistically significant.
Results
Posterior calf vibration (‘Calf only’, ‘Calf-Tib’ and ‘Tib-Calf’)
The induced torque variances for posterior calf vibration for the 2 days are demonstrated in Fig. 2 along with the paired statistical analysis. The vibratory stimulus resulted in increased torque variance in all groups (P < 0.001). During the stimulation the induced torque decreased significantly from period 1 to period 4 in all groups (P < 0.001). No difference in elicited torque could be found between the groups in any of the days. In Fig. 3 is normalized raw data from one representative subject from group ‘Calf only’ shown, demonstrating an initial backward sway in response to the stimulation and a shift of stance into a more forward position as the test progressed.

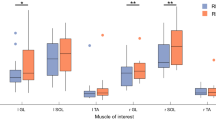
Torque variance in response to gastrocnemius vibration. Mean and standard error of mean (SEM) of elicited torque in anteroposterior direction on both test days. Only statistical significant are denoted in the figure. The annotations above the graph show the differences between the test days (Wilcoxon). The annotations below the graph demonstrate the significant differences in induced torque between periods of each test, and demonstrate a similar torque reduction during the test in all groups on both days (Wilcoxon). *P < 0.0167, **P < 0.01 and ***P < 0.001. The numerical P values in the above figure were not considered significant. The statistics were corrected according to Bonferroni, in the case of Wilcoxon with a factor of 3, and Mann–Whitney, a factor of 2, yielding P < 0.0167 and <0.025, respectively, that were considered to be statistically significant

Normalized torque elicited to vibration stimulation. Normalized ‘raw data’ from three subjects on the first day. At the top is data from a subject exposed to posterior calf vibration (‘Calf only’), in the middle a subject exposed to tibialis anterior vibration succeeding posterior calf vibration (‘Calf-Tib’) and at the bottom a subject exposed to posterior calf vibration (‘Tib only’). Please note the difference in response amplitude throughout the test between ‘Calf-Tib’ compared to ‘Tib only’. Please also note the different initial displacements between ‘Calf only’ and ‘Calf-Tib’ and the similar forward leaning of body position
Tibialis anterior vibration (‘Tib only’, ‘Calf-Tib’ and ‘Tib-Calf’)
The induced torque variances for tibialis anterior vibration for the 2 days are demonstrated in Fig. 4. The vibratory stimulus resulted in increased torque variance in all groups (for P values please see Fig. 4). During the stimulation, the induced torque decreased significantly from period 1 to period 4 in all groups on both days, except for group ‘Tib-Calf’ on the second day. There were no differences in torque variance between the 2 days in any group in any period.

Torque variance in response to tibialis anterior vibration. Mean and standard error of mean (SEM) of elicited torque in anteroposterior direction on both test days. Only significant differences are shown in the figure. The annotation above the graph demonstrates the differences between the groups (Mann–Whitney). Please note the difference between ‘Calf-Tib’ and the other two groups. The annotation below the graph demonstrate the significant differences in induced torque during each test, and demonstrate a similar torque reduction during the test in all groups both days (Wilcoxon). *P < 0.025, **P < 0.01 and ***P < 0.001. The numerical P values in the above figure were not considered significant. The statistics were corrected according to Bonferroni, in the case of Wilcoxon with a factor of 3, and Mann–Whitney, a factor of 2, yielding P < 0.0167 and <0.025, respectively, that were considered to be statistically significant
Group ‘Calf-Tib’ that received posterior calf vibration before tibialis anterior vibration had significantly more induced torque than the other two groups in the first stimulation period on both days (Fig. 4). In addition to yielding more torque, previous gastrocnemius vibration induced also a different positioning of the body, around which the forces that act are generated to withstand perturbations (Fig. 3).
Order dependency
The ANOVA analysis in Table 1 demonstrates that torque variance decreased when the gastrocnemius was stimulated (group ‘Calf only’) both during test and between the days. When tibialis anterior was stimulated (group ‘Tib only’), however, were there only reductions of torque variance during each test and not between the tests.
When both locations were stimulated though in different order, could the same results be extrapolated, since the interaction between ‘Stimulation × Period’ was not significant—indicating the same reduction of torque variance during the test regardless the vibrated muscle. However, the interaction between ‘Stimuli × Day’ was significant (P < 0.001) demonstrating that the reduction of torque variance between the days was related to vibration site.
The order of to which muscle vibration was applied had great significance (P < 0.001), and it is obvious from the Figs. 2 and 4 that this is due to a higher torque variance in group ‘Calf-Tib’ when tibialis anterior vibration succeeded posterior calf vibration.
Discussion
Measuring torque variance during vibratory perturbation gives an appreciation of the energy that is spent on maintaining postural control while resolving a sensory conflict (Magnusson et al. 1990). As energy conservation is a fundamental principle, it would seem plausible that a postural task is handled with more ease and is better performed, if less energy is consumed (Johansson and Magnusson 1991).
Effects of different response amplitude
The results indicate that there are differences in how the postural control system respond and adapt to vibration to different muscles. Both tibialis anterior and posterior calf vibration induced significant increases of torque variance, though with different amplitudes (posterior calf greater). Both stimuli yielded reduction of the torque variance over time (during the test), but only the posterior calf vibration generated a reduction from day to day, i.e. induced responses started on a lower level the second day. This suggests that the amplitude or strength of the induced perturbations is of importance whether the reduction of responses will be learned or not. Despite that tibialis anterior stimulation yielded a decrease or an adaptation within the test, the perturbation was not strong enough to induce a further adaptation of the learned strategy to reduce the body sway to the succeeding day. In other words, the experience from the day before was not advantageous in order to withstand perturbations when tibialis anterior was stimulated; the memory from day 1 was not consolidated to the second day. This is well in line with the existing theories on memory processing since it is generally believed that the stimulus (or learning) is required to be of sufficiently large amplitude to induce alterations at synapses held to be involved in consolidation processes (Bliss and Collingridge 1993). Furthermore it correlates well to what has been demonstrated in studies on learning ability, where more challenging tasks yields improved learning performances (Mahncke et al. 2006).
The reductions of torque variance when exposed to posterior calf vibration within a test were similar, irrespective if the subjects had been subjected to tibialis anterior vibration immediately before or afterwards. This implies that processes involved in consolidating postural experiences of sufficient difficulty are insensitive to similar but antagonistic exercises executed in close proximity to the task, contrary to other types of motor learning (Brashers-Krug et al. 1996; Shadmehr and Brashers-Krug 1997). One explanation could be that the adaptation involved in maintaining balance is so vital in coping with postural threats, that an unlearning would make no sense from an evolutional perspective (Dorris et al. 2000). In animal models it has been shown that a perceived life-threatening situation induces fast consolidation of motor memories (Bohbot et al. 1996). Long-term adaptation to repeated posterior calf vibratory stimulation has been demonstrated with test intervals of 20 min (Tjernstrom et al. 2005) although it must be emphasized that postural control seemed to be better learned with longer intervals than, providing ample time for cellular mechanisms for consolidation processes to develop (Karni et al. 1998). Another explanation to the lack of interference on learning or consolidation to the second day could lie in the nature of the ‘second task’. The action of tibialis anterior is not exactly antagonistic to gastrocnemius and soleus. However, Fig. 3 implies that early responses to tibialis anterior stimulation yielded forward sway and early responses to posterior calf stimulation yielded backward sway, thus being antagonistic in that respect. The postural responses to posterior calf vibration in groups ‘Calf only’, ‘Calf-Tib’ and ‘Tib-Calf’ generated a forward leaning over time, presumably a better position to generate forces to withstand perturbations. The postural responses to tibialis anterior vibration only or prior to posterior calf vibration (group ‘Tib only’ and ‘Tib-Calf’) did not lead to any adjustments of body position (Fig. 3).
Effects of stimuli order
The order in which the subjects were exposed to tibialis anterior or posterior calf vibration had impact on the amplitude of torque variance. Higher amplitudes of torque variance in response to tibialis anterior vibration were generated when that stimulus succeeded posterior calf vibration (group ‘Calf-Tib’). In Fig. 3 this is further demonstrated, in which another effect of the stimuli order is evident. The subject from group ‘Calf-Tib’ besides responding with greater torque also leaned forward when receiving vibration to tibialis anterior, as is common and previously observed with posterior calf vibration (Tjernstrom et al. 2002). This postural strategy to control the perturbation was not observed in group ‘Tib only’ or ‘Tib-Calf’, which suggests that it was induced by the previous experience from posterior calf vibration. The higher amplitude of torque variance corresponds to other studies when responses are scaled to the preceding amplitude (Diener and Dichgans 1988). These observations have led to the conclusion that humans employ internal models in a feed-forward manner to control upright posture, i.e. preformed ways to respond to a perturbation (Diener et al. 1988). The preformed responses have been reported to decrease as the test progress, related to learning from feedback information. However, in the present study the responses of greater magnitude and leaning in group ‘Calf-Tib’ did not decrease to the same levels as the other two groups, which would be the obvious way of dealing with tibialis anterior vibration. This could be due to the non-predictable pattern of the pseudo-randomized stimulation pulses used in the present study as opposed to other studies where the stimulation was constant (Diener and Dichgans 1988). Another plausible explanation could be that the internal model or strategy, that may have been developed by the control system during posterior calf vibration, consisted of a sensory reweighting process, where the control system rely more on vestibular and other somatosensory cues. The control system during tibialis anterior vibration seemed to rely less on sensory information from the lower legs, resulting in greater amount of torque variance than was necessary (Figs. 3, 4). This suggests that the feedback loop conveying sensory information from the vibrated muscle was ignored or at least postponed from the beginning of the stimulation. This corresponds to the hypothesis underlying sensory reweighting processes, in this case instigated by experiencing posterior calf vibration previously.
No effect on the responses to posterior calf vibration could be discerned if it succeeded tibialis anterior stimulation (group ‘Tib-Calf’). This could also be an effect of the lesser response amplitude to tibialis anterior vibration, not being powerful enough to induce a preferred strategy or that possible effect was negligible compared to the induced responses to posterior calf vibration.
The greater magnitude of the induced torque variance in group ‘Calf-Tib’ could also be due to a vibration after-effect (Wierzbicka et al. 1998). However, this seems more unlikely since quiet stance was unaffected by the previous posterior calf stimulation, and similar studies on posterior calf vibration during test conditions with open or closed eyes, which yields different torque-amplitudes, has not indicated any differences in torque variance regardless the order in which the tests were performed, eyes closed first or vice versa (Tjernstrom et al. 2002).
To conclude; a postural perturbation has to be sufficiently difficult to withstand in order to induce long-term learning, and experiences or postural strategies can be transferred between postural challenges if they post different demands. If these observations are generalized it follows that training exercises employed in rehabilitation programs for patients suffering from sensory loss or a postural disturbance, should be sufficiently challenging to instigate learning processes. Furthermore, exercises should be spaced and varied in order not to activate inappropriate internal models from previous training exercises.
References
Abel T, Lattal KM (2001) Molecular mechanisms of memory acquisition, consolidation and retrieval. Curr Opin Neurobiol 11(2):180–187
Altman D (1991) Practical statistics for medical research. Chapman & Hall, New York
Bliss TV, Collingridge GL (1993) A synaptic model of memory: long-term potentiation in the hippocampus. Nature 361(6407):31–39
Bohbot V, Otahal P et al (1996) Electroconvulsive shock and lidocaine reveal rapid consolidation of spatial working memory in the water maze. Proc Natl Acad Sci USA 93(9):4016–4019
Brashers-Krug T, Shadmehr R et al (1996) Consolidation in human motor memory. Nature 382(6588):252–255
Diener HC, Dichgans J (1988) On the role of vestibular, visual and somatosensory information for dynamic postural control in humans. Prog Brain Res 76:253–262
Diener HC, Horak FB et al (1988) Influence of stimulus parameters on human postural responses. J Neurophysiol 59(6):1888–1905
Dorris MC, Pare M et al (2000) Immediate neural plasticity shapes motor performance. J Neurosci 20(1):RC52
Eklund G (1973) Further studies of vibration-induced effects on balance. Ups J Med Sci 78(1):65–72
Frank JS, Earl M (1990) Coordination of posture and movement. Phys Ther 70(12):855–863
Fransson PA, Johansson R et al (2003) Adaptation to vibratory perturbations in postural control. IEEE Eng Med Biol Mag 22(2):53–57
Fransson PA, Hjerpe M et al (2007) Adaptation of multi-segmented body movements during vibratory proprioceptive and galvanic vestibular stimulation. J Vestib Res 17(1):47–62
Goodwin GM, McCloskey DI et al (1972) Proprioceptive illusions induced by muscle vibration: contribution by muscle spindles to perception. Science 75(28):1382–1384
Johansson R (1993) System modeling and identification. Prentice-Hall, Englewood Cliffs
Johansson R, Magnusson M (1991) Optimal coordination and control of posture and locomotion. Math Biosci 103(2):203–244
Johansson R, Magnusson M et al (1988) Identification of human postural dynamics. IEEE Trans Biomed Eng 35(10):858–869
Karni A, Meyer G et al (1998) The acquisition of skilled motor performance: fast and slow experience-driven changes in primary motor cortex. Proc Natl Acad Sci USA 95(3):861–868
Magnusson M, Johansson R et al (1990) Galvanically induced body sway in the anterior-posterior plane. Acta Otolaryngol 110(1–2):11–17
Mahncke HW, Bronstone A et al (2006) Brain plasticity and functional losses in the aged: scientific bases for a novel intervention. Prog Brain Res 157:81–109
Matthews P (1986) What are the afferents of origin of the human stretch reflex, and is it a purely spinal reaction? In: Freund HJ, Büttner U, Cohen B, Noth J (eds) Progress in brain research, vol 64. Elsevier (Biomechanical Division), Amsterdam, pp 55–66
McGaugh JL (1966) Time-dependent processes in memory storage. Science 153(742):1351–1358
Oie KS, Kiemel T et al (2002) Multisensory fusion: simultaneous re-weighting of vision and touch for the control of human posture. Brain Res Cogn Brain Res 14(1):164–176
Patel M, Gomez S et al (2008) Effects of 24-h and 36-h sleep deprivation on human postural control and adaptation. Exp Brain Res 185(2):165–173
Peterka RJ (2002) Sensorimotor integration in human postural control. J Neurophysiol 88(3):1097–1118
Sanes JN (2003) Neocortical mechanisms in motor learning. Curr Opin Neurobiol 13(2):225–231
Shadmehr R, Brashers-Krug T (1997) Functional stages in the formation of human long-term motor memory. J Neurosci 17(1):409–419
Shen B, McNaughton BL (1996) Modeling the spontaneous reactivation of experience-specific hippocampal cell assembles during sleep. Hippocampus 6(6):685–692
Tjernstrom F, Fransson PA et al (2002) Adaptation of postural control to perturbations—a process that initiates long-term motor memory. Gait Posture 15(1):75–82
Tjernstrom F, Fransson PA et al (2005) Improved postural control through repetition and consolidation. J Vestib Res 15(1):31–39
Wierzbicka MM, Gilhodes JC et al (1998) Vibration-induced postural posteffects. J Neurophysiol 79(1):143–150
Wilson MA, McNaughton BL (1994) Reactivation of hippocampal ensemble memories during sleep. Science 265(5172):676–679
Wolpert DM, Ghahramani Z et al (1995) An internal model for sensorimotor integration. Science 269(5232):1880–1882
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Tjernström, F., Fransson, PA., Patel, M. et al. Postural control and adaptation are influenced by preceding postural challenges. Exp Brain Res 202, 613–621 (2010). https://doi.org/10.1007/s00221-010-2166-x
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00221-010-2166-x