
Abstract
This article investigated both the ability of naive human subjects to learn interval production, as well as the properties of learning generalization across modalities and interval durations that varied systematically from the over-trained interval. Human subjects trained on a 450-, 650-, or 850-ms single-interval production task, using auditory stimuli to define the intervals, showed a significant decrease in performance variability with intensive training. This learning generalized to the visual modality and to non-trained durations following a Gaussian transfer pattern. However, the learning carryover followed different rules, depending on the duration of the trained interval as follows: (1) the dispersion of the generalization curve increased as a function of the trained interval, (2) the generalization pattern was tilted to the right in the visual condition, and (3) the transfer magnitude for 650 ms was less prominent than for the other two intervals. These findings suggest the existence of neural circuits that are tuned to specific time lengths and that show different temporal processing properties depending on their preferred interval duration.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
The ability to quantify time in the range of hundreds of milliseconds is a fundamental process in a wide range of sensory and motor tasks (Ivry 1996; Gibbon et al. 1997; Buonomano and Karmarkar 2002; Merchant et al. 2008a). Time in this range is essential for perception of speed, for producing and understanding speech and music, and for dancing and performing sports (Mauk and Buonomano 2004; Merchant and Georgopoulos 2006; Merchant et al. 2009). Hence, such complex behaviors depend on temporal information processing at perceptual, motor, and cognitive levels. Two opposite mechanisms have been proposed as the neural substrate of interval timing in the hundreds of milliseconds (Ivry and Schlerf 2008; Mauk and Buonomano 2004): a centralized mechanism that processes temporal information in a multimodal fashion, and across perceptual and motor-timing tasks; and a distributed mechanism that involves a specific neural circuit for different timing behaviors. Recent neuroimaging and psychophysical studies have led to an intermediate hypothesis, namely, that interval timing depends on a partially overlapping distributed mechanism, where main-core cortical and subcortical timing structures, such as SMA, prefrontal, and posterior parietal cortex, as well as the basal ganglia and the cerebellum, can be influenced differently by context-dependent information that is processed by the corresponding brain areas (Lewis and Miall 2003; Merchant et al. 2008a, b; Grondin 2001).
In this context, the study of perceptual interval learning and the generalization properties of such learning have provided important insights into the neural underpinnings of temporal information processing. For example, using interval discrimination it has been shown that intensive learning can generalize across untrained auditory frequencies (Wright et al. 1997; Karmarkar and Buonomano 2003), sensory modalities (Nagarajan et al. 1998; Westheimer 1999), stimulus locations (Nagarajan et al. 1998), and even from sensory to motor-timing tasks (Meegan et al. 2000). However, all these studies found no generalization toward untrained interval durations. It has been suggested that the learning transfer depends on the improvement of temporal processing and not on more efficient memory or decision processes, at least for auditory interval discrimination (Karmarkar and Buonomano 2003). Therefore, these findings not only support the notion of a centralized or a partially overlapping distributed timing mechanism, but they also introduce the concept of duration-specific circuits. Of course, these two features are not mutually exclusive when a large timing neural network is considered (Karmarkar and Buonomano 2007). Nevertheless, these studies have only tested large differences between the trained and untrained testing intervals, precluding the investigation of a potential generalization curve across nearby durations, which could have different shapes and widths depending on the length of the trained interval. Furthermore, timing generalization studies have focused on perceptual rather than motor timing, which could have different learning transfer properties if the timing mechanism is not totally centralized in these two temporal processing conditions (Merchant et al. 2008a).
Thus, in the present study, we investigated the generalization properties of intensive, single-interval production learning over a wide range of non-trained intervals (starting with small differences from the trained interval), in human subjects performing the task for one of three different training intervals. In addition, we tested whether the generalization functions were modulated by the modality of the stimuli used to define the interval. The results support the existence of interval-tuned neural populations that show different temporal processing properties according to specific generalization rules, which depend on the duration and modality of the trained interval.
Methods
Subjects
Forty volunteers (20M, 20F) between the ages of 18 and 41 years (mean 25.47) with normal hearing, and normal or corrected vision participated in this study. All subjects volunteered and gave written consent for this study before commencement of experiments. The study complied with the Declaration of Helsinki and was approved by the National University of Mexico Institutional Review Board.
Apparatus
Participants were comfortably seated on a chair facing a computer screen in a quiet room, and they tapped on the spacebar of a computer keyboard. All participants performed the task using a finger of their preferred hand (one subject was left-handed). The subjects could not see their own hand during tapping; the auditory stimuli were presented through noise-canceling headphones (Sony, MDR-NC50), and, in addition, subjects listened to white noise while performing the task. Hence, the participants did not have auditory or visual feedback from the tapping movement. For data collection, a custom-made Visual Basic program was used (Microsoft Visual Basic 6.0). This program controlled the timing of the stimuli and collected the execution data. Computers with high-speed processors (>2.0 GHz) were used for data collection.
Task
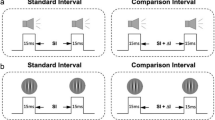
Subjects performed a single time intervals production task. For a given interval duration, this task had instruction and internally driven trials presented in blocks (see Fig. 1a). At the beginning of each trial in the instruction period, a target interval (two stimuli separated by a particular duration or inter-stimulus interval [ISI]) was presented, and then the subject tapped twice on the push-button to reproduce the same interval. This was repeated for n 1 instruction trials. Immediately after, the subject entered the internally driven period that consisted of n 2 trials in which he/she reproduced a single interval after a go signal appeared on the screen. Thus, the instruction period involved the reproduction of a time interval using an external guide that was renewed from trial to trial, whereas the internally driven period required the production of a time interval using an internal temporal representation formed during the instruction. Subjects were requested to respond 200 ms after presentation of the target interval or the go signal, with no other restriction than a maximum reaction time of 2,000 ms. This reaction time was 689.8 ± 242 ms (mean ± SD). The inter-trial interval was 1.5 s. We tested intervals delimited by auditory (pure tone, 32 ms, 1,000 Hz, 80 dB) or visual (green square, 16 ms, 10-cm side) stimulus markers.

a Illustration of the single interval production task showing one trial of each period (instruction and internally driven). ISI inter-stimuli interval, IRI inter-tap interval. b The three phases of the experiment (pre-test, training phase, and post-test) are shown with the corresponding number of days. During the pre- and post- test, the subjects performed both periods of the single-interval production task for a set of seven durations, each duration presented in a block of 5 instruction trials plus 20 internally driven trials. The seven duration blocks were first delimited by auditory stimuli, and then were delimited by visual stimuli in independent blocks. During the training phase, the subjects completed 60 blocks of trials per day. Each block consisted of 5 instruction trials plus 15 internally driven trials, using only the standard interval delimited always by auditory stimuli
Procedure
The generalization of interval production was investigated with respect to the abilities of human subjects to transfer learning from three standard intervals (450, 650, or 850 ms) across their contiguous untrained intervals and across the modality of the stimuli used to delimit the interval. Subjects were assigned pseudorandomly to the 450- (n = 14), 650- (n = 12), or the 850-ms (n = 14) standard intervals.
For each standard interval, the experiment consisted of a pre-test, a training phase, and a post-test for a total of ten consecutive sessions, one session per day (see Fig. 1b). In the pre- and post-test sessions, the performance of the subjects on the single-interval production task was measured using a set of seven target intervals that varied in duration systematically around the standard interval (see Table 1). Of course, this set included the standard interval. In order to determine the generalization process for as many target intervals as possible, the subjects were assigned pseudorandomly to one of two sets of target intervals (protocol A or B in Table 1). Therefore, for a particular standard interval, we tested 12 target intervals in two subgroups of subjects. In addition, the order of presentation of these seven target intervals was random. During the pre- and post-test, each block consisted of 5 trials in the instruction period followed by 20 trials of the internally driven period. The task was initially performed using auditory markers for delimiting the ISI for all the target intervals, and then it was repeated using visual markers. We limited the number of trials in these two conditions with the purpose of avoiding a learning process during the pre- and post-test. Nevertheless, it is important to mention that before the pre-test data collection, practice trials were given for a set of intervals that was different from the target intervals used during the pre- and post-test periods. The practice session continued until the subjects acknowledged that they understood the task and were comfortable with their performance. Feedback was given after each trial of the instruction period, showing the subject his/her produced inter-tap interval as a number on the screen. However, no feedback was given during the internally driven period, in order to be able to measure the reliability of the interval representation acquired during the training phase.
In the training phase, the subjects completed 60 task blocks (900 trials, ~60 min) each day, using the standard interval delimited only by auditory markers. In this case, a block consisted of 5 trials in the instruction period followed by 15 trials of the internally driven period, and a 5-min pause was introduced every 20 blocks. The training phase lasted for eight consecutive days. All trials (instruction and internally driven) during training were followed by feedback, since pilot studies showed that feedback was critical for appropriate learning (data not shown).
Data analysis
General
The standard deviation (SD) of the produced intervals was used as a measure of performance and was calculated for each block during the instruction and internally driven periods. Produced intervals were considered outliers and discarded when their duration was 1.5 times below or above the 20th and 80th percentile, respectively.
Standard statistical techniques were used for data analysis including the t-test, repeated-measures analysis of variance (ANOVA), and linear regression (Snedecor and Cochran 1989). The reported P-values in the repeated-measures ANOVAs correspond to the Greenhouse–Geisser test, which corrects for possible deviations in sphericity. The level of statistical significance to reject the null hypothesis was α = 0.05. Subroutines written in Matlab (version 7.3.0.267, R2006b) and the SPSS statistical package (version 12, SPSS Inc., Chicago, IL, 2003) were used for the statistical analyses.
Transfer of learning
Transfer curves were constructed by subtracting the SD of the post-test from the pre-test for each target interval. We called this measure PrePost-SDDiff. Hence, positive values of PrePost-SDDiff indicate an improvement in task performance.
Results
The single-interval production task required the execution of two tapping movements separated by a particular duration, in trials separated in two different conditions or periods. During the instruction period, subjects reproduced a time interval that was previously presented as two brief sensory stimuli, whereas in the internally driven period subjects produced the same interval after a go signal, which implied the use of an internal interval representation that was previously obtained during the instruction period (see “Methods” section, Fig. 1a). These two task periods were presented in blocks of trials for a particular interval duration and marker modality. In order to determine the learning abilities of interval reproduction and the learning generalization properties across modalities and non-trained interval durations, the subjects performed both periods of the single-interval production task during three experimental conditions: a pre-test, a training phase, and a post-test (Fig. 1b). The difference in performance variability between the post- and pre-test is a measure that not only reflects the overall learning achieved during the training phase for the over-trained interval duration, but also reveals the learning generalization abilities across non-trained conditions. Accordingly, during the pre- and post-tests of the present study, the subjects executed the single-interval production task for different interval durations defined by auditory or visual markers, whereas in the training phase they performed the task for only one of three standard intervals in the auditory marker condition for eight consecutive days (Fig. 1b). Hence, the initial question in this experiment was whether the intensive practice during the training phase could decrease the variability of interval production in human subjects.
Interval-production learning
Figure 2 shows the averaged standard deviation (±SEM across subjects) of the produced intervals plotted as a function of the training day for the instruction (left) and internally driven (right) periods of the task, and for the three standard intervals (450, 650, and 850 ms). It is clear that the performance improved as a result of the training in all conditions, following a typical learning curve across standard intervals and task periods. However, subjects were pooled for this and the subsequent analyses only if they showed: (1) a significant change in performance across the training days according to a repeated-measures ANOVA (data not shown), and (2) a negative slope in their power regression learning curve (see Eq. 1 below). In fact, five subjects did not meet these criteria and were eliminated from the analyses: one for 450-, two for 650-, and two for the 850-ms standard intervals.

Averaged learning curves. SD (mean ± SEM) of the produced intervals during the training phase plotted as a function of the training day. Filled circles, crosses, and open circles are data for 450, 650, and 850 ms, respectively. The lines are the predicted learning curves calculated from the fitting of data to a power function, solid lines for 450 ms (instruction: R 2 = 0.8938, P = 0.0004; test: R 2 = 0.9421, P = 0.0001), dotted lines for 650 ms (instruction: R 2 = 0.7739, P = 0.004; test: R 2 = 0.8996, P = 0.0003), and dashed lines for 850 ms (instruction: R 2 = 0.6498, P = 0.0157; test: R 2 = 0.8518, P = 0.0011)
We carried out a repeated-measures ANOVA for each task period using the SD as dependent variable, the days of training as the within-subject factor, and the standard interval as the between-subject factor. For the internally driven period, the results showed significant main effects on days of training (F 7,224 = 21.536, P < 0.0001) and standard interval (F 2,32 = 17.607, P < 0.0001), as well as on the day × interval interaction (F 14,224 = 2.116, P = 0.05). The analysis of the instruction period showed similar results, with significant main effects on days of training (F 7,224 = 10.169, P < 0.0001) and standard interval (F 2,32 = 20.2, P < 0.0001), and on the day × interval interaction (F 14,224 = 2.107, P = 0.045). Therefore, these findings support the idea that training improves the production of time intervals across standard intervals and task periods. In addition, this analysis indicates that the learning dynamics and/or the initial timing variability was different across standard intervals. Therefore, in order to characterize further the learning process, we fitted the following power function to the learning curves for each subject:
where SD is the standard deviation of the produced intervals, T the day of training, b the intercept, and m the time constant (slope) of the power function. Since the fittings of this function were appropriate (see Fig. 2 legend), the next step was to test whether the slope and intercept of the learning curves were different across standard intervals and task periods. Table 2 shows the means (±SEM across subjects) of the power function coefficients for the different standard intervals and task periods, as well as the mean (±SEM) of the SD difference between the last and first training days (SDdiff). ANOVAs were performed using each of these descriptive parameters (m, b, and SDdiff) as dependent variables, and the standard interval and task period (instruction vs. internally driven) as factors. The results showed no significant main effects of the standard interval on the time constant of the learning curves (m), suggesting that the learning time course was independent of the trained interval. In contrast, the standard interval showed significant main effects when the intercept b (F 2,64 = 42.99, P < 0.0001) and SDdiff (F 2,64 = 3.782, P = 0.028) were used as dependent variables. These last results indicate that the initial temporal variability followed the scalar property of temporal processing (Gibbon et al. 1997; Merchant et al. 2008a, b, c, d).
In addition, the results of the ANOVAs showed that during the internally driven period, m (F 1,64 = 5.336, P = 0.024), b (F 1,64 = 4.808, P = 0.032), and SDDiff (F 1,64 = 3.209, P = 0.078, marginal effect) were significantly larger than during the instruction period (see Table 2), suggesting that the learning process based on an internal representation of time was larger than during the instruction period, where subjects used their working memory to reproduce the standard interval.
Overall, these results confirm that extensive training in the single-interval production task induces a learning process characterized by a decrease in the temporal processing variability as a function of the training day. This learning curve was an asymptotic function with a negative slope that was not influenced by the trained interval.
Learning generalization of interval production
Changes in performance variability from the pre- to the post-test determined the magnitude of learning in the trained standard interval using auditory markers, as well as the magnitude of generalization in the untrained conditions due to learning. Consequently, as a first step, we compared the SD of the standard interval measured before and after the training phase for each subject during the internally driven period (Suppl. Fig. S1). We found that most subjects showed a reduction of the interval-production variability as a result of extensive training, except for those that were eliminated from the analysis due to their lack of learning (see above).
The main objective of the present study was to determine the transfer rules for interval-production learning. Specifically, we were interested in studying the transfer properties of single-interval production learning over non-trained (target) intervals and across the modalities used for delimiting the interval duration. Thus, an ANOVA was carried out using a repeated measures design, in which the SD was the dependent variable, the experimental phase (pre- or post-test) was the within-subject factor, and the standard interval, target interval, and modality were the between-subject factors. The results showed a significant main effect for experimental phase (F 1,391 = 197.577, P < 0.0001), as well as a significant effect for the phase × standard interval interaction (F 2,391 = 6.055, P = 0.003), but not a significant effect for the phase × target interval interaction (F 27,391 = 1.32, P = 0.132). Additionally, significant main effects were found for standard interval (F 2,391 = 4.658, P = 0.01) and target interval (F 27,391 = 12.346, P < 0.0001). In contrast, no significant main effects were found for the modality or its interaction with the other two factors. Therefore, these results indicate that after training there was a generalization process to some of the non-trained intervals, characterized by a significant decrease in performance variability in target intervals. This after-training improvement across target intervals was modulated by the standard interval, but not by the modality of the marker.
With the aim of studying further the properties of the interval-production learning generalization, transfer curves were constructed by subtracting the SD of the post- from the pre-test for each target interval. We called this measure PrePost - SDDiff. Figure 3 corresponds to the interquartile box-plots of the PrePost - SDDiff as a function of the target interval for all the standard intervals (450, 650, and 850 ms). t-Tests were performed for each target interval to determine whether PrePost - SDDiff was statistically different from zero. Asterisks above the interquartile box in these figures indicate the statistical significance of the generalization.

Pre-Post SD difference during the internally driven period. Inter-quartile ranges (white bars) and medians (black line within bars) of Pre-Post difference are plotted as a function of the target interval. Auditory (top) and visual marker condition (bottom). One asterisk indicates marginal effects (P < 0.1), two asterisks indicate significant (P < 0.05) differences from zero of the Pre–Post SD difference as measured by one sample t-test
Figure 3 (top) shows a specific generalization pattern for the 450-ms standard interval, with a smooth decrease in the PrePost - SDDiff as the target interval was farther away from the standard interval, describing a generalization curve that was similar to a neural-tuning function. In addition, the results showed some degree of cross-modal generalization for the 450-ms standard interval. For visual markers, the generalization curve was tilted to the right with respect to the auditory curve.
For the 650-ms standard interval, there was a generalization pattern that was less structured across target intervals or marker modalities (Fig. 3 middle). In fact, fewer intervals showed a significant change in the PrePost - SDDiff. Hence, we concluded that a less prominent transfer accompanied the extensive training for the 650-ms interval.
The generalization curves for the 850-ms standard, depicted in Fig. 3 (bottom), show an extensive transfer of learning across target intervals and marker modalities. Indeed, the generalization was wide with respect to the number of target intervals around the standard with a significant PrePost - SDDiff. Remarkably, the transfer was very similar for the non-trained modality, showing a generalization for visual markers that was as large as in the auditory condition.
A Gaussian function was fitted to the PrePost - SDDiff as a function of the target interval to determine the mean and half-height dispersion of the generalization curves above. The resulting significant fittings are shown in Fig. 4a, where it is evident that the proportion of variance accounted for (R 2) by the Gaussian models was lower for the 650-ms standard interval. This is in accordance with the previous results showing that, for this standard interval, the generalization across non-trained intervals was less evident than for the other two standard intervals (see above). Nevertheless, it is important to emphasize that four distinctive generalization rules could be extracted from these Gaussian fittings: (1) the half-height dispersion of the generalization curves increased as a function of the standard interval, (2) the generalization curves for the visual marker condition were tilted to the right with respect to the auditory marker condition, (3) the bias of the distribution mean with respect to the actual standard interval increased as a function of the magnitude of this interval, and (4) both marker modalities showed similar height and dispersion in their Gaussian distribution. Indeed, Fig. 4b shows that there was a linear increase in the half-height dispersion as a function of the standard interval for both marker modalities. In addition, the results showed that there was a linear decrease in the constant error (standard interval − distribution mean) as a function of the standard interval (Fig. 4c). Finally, this linear decrease in constant error was shifted upwards in the visual marker condition across the standard intervals (Fig. 4c).

a Gaussian fittings of the Pre-Post SD difference for the different standard intervals and the two interval-marker modalities. b Half-height dispersion as a function of standard interval and c constant error as a function of standard interval. In b and c, the auditory and visual interval markers are represented by filled circles—solid lines, and open circles—dashed lines, respectively
Timing and sensorimotor transfer
A question that emerged from the results was whether the observed generalization could be due to sensorimotor and/or procedural learning instead of interval-timing learning, given that an improvement in sensorimotor processing could lead to similar findings. To address this issue, we took advantage of the structure of the task that was composed of two periods: the instruction period that required the reproduction of a time interval using an external guide that was renewed from trial to trial, and the internally driven period that involved the production of a time interval using an internal temporal representation. Thus, if the transfer of learning was mainly due to the sensorimotor component of the task, we would expect a similar generalization pattern in both task periods. To test this hypothesis, first we normalized the Pre-Post_SDDiff to be able to compare the transfer magnitude between the instruction and internally driven periods, since each period had a different number of trials. Thus, the Pre-Post_SDDiff was divided by the Pre-test SD, and this ratio was multiplied by 100, and expressed as a percentage of the Pre-test SD. This measure, called percentage of gain, was used as the dependent variable in an ANOVA in which the task period, the standard interval, and the modality were used as factors. Interestingly, we found a significant main effect for task period (F 1,636 = 12.799, P < 0.0001), but non-significant effects on the two- and three-way interactions between the task period and the other independent variables. In fact, Table 3 shows that the mean values of the percentage of gain were larger in the internally driven than in the instruction period across standard intervals and modalities. Therefore, these results suggest that the reported learning generalization in the internally driven period was mainly due to an increase in precision of an internal temporal processing mechanism.
Discussion
The purpose of this study was to determine the capability of interval-production learning in naive subjects and to determine the generalization properties of this learning across a wide range of untrained intervals and between two sensory modalities used as interval markers. Regarding the first point, the results revealed that a learning process occurred during intensive training for single-interval production. In fact, as far as we know our results represent the first demonstration of interval-production learning for different trained intervals in human subjects. Interestingly, the time course of learning observed across trained intervals in this study is consistent with that observed for auditory- and somatosensory-interval discrimination (Kristofferson 1980; Wright et al. 1997; Nagarajan et al. 1998; Karmarkar and Buonomano 2003; Westheimer 1999). All these experiments consisted of an intensive daily training session repeated for more than 7 days. Under these circumstances, the learning occurred mainly during an initial rapid-improvement stage that lasted for 2 or 3 days, followed by a slower-improvement phase that spanned the remaining sessions. Therefore, our data, together with the findings reported in the literature, suggest that besides the initial procedural learning involved in each particular task, there is a learning process that may depend on the common or overlapping timing mechanism.
Not all subjects showed a learning process according to our criteria (negative slope for the learning curves and significant change in SD across training days). Since the learning transfer found in non-learners may be interpreted as noise, these subjects were excluded from further analyses, as in previous timing generalization studies (Wright et al. 1997; Karmarkar and Buonomano 2003).
An important finding of this study was the generalization pattern across untrained intervals that were contiguous to the trained standard interval, which at first glance seems to be at odds with the previous reports of interval-specific perceptual generalization (Kristofferson 1980; Wright et al. 1997; Nagarajan et al. 1998; Karmarkar and Buonomano 2003). However, it is important to consider, first, that most previous studies used shorter intervals than those used here, and second, that we tested the generalization of interval production instead of interval discrimination. More relevant is the fact that the earlier experiments used untrained intervals differing by more than 50% from the trained intervals, decreasing their sensitivity for detecting a cross-interval generalization process. Consequently, the present results suggest the existence of plastic neural networks that are duration specific but that follow these rules: (1) their interval specificity decreases as a function of the trained interval, showing the scalar property at the neurophysiological level; (2) the circuits have access to bimodal information, but the intervals defined by visual stimuli are produced shorter than the same intervals defined by auditory markers.
Concerning the first generalization rule, it has been shown that in both a self-paced and during the continuation phase of a synchronization-continuation tapping task, there is a clear execution preference toward particular intervals, following a categorical process. Interestingly, this preference follows a multimodal distribution that converges to zero around 250, 450, and 788 ms, with a large bias around 650 ms (Collyer et al. 1992, 1994). Accordingly, in interval discrimination studies, it was found that after extensive practice the variability in temporal performance reveals temporal categories or steps (Kristofferson 1980). In addition, Collyer et al. (1994) found that the dispersion of the self-paced intervals also increased as a function of the preferred duration. Therefore, these experiments not only support our hypothesis of duration-specific neural circuits that follow the scalar property, but they also emphasize the concept of under-representation of intervals around 650 ms in these circuits. The less prominent generalization pattern for the 650-ms standard interval could be the result of uneven requirements in the temporal-motor processing across behaviors of every-day life (i.e., Fant and Kruckenberg 1996; Moelants 2002), and/or the result of the neurophysiological properties of the timing circuits.
At least two non-mutually exclusive neural mechanisms could explain the duration specificity, the scalar property, and the 650-ms under-representation. It has been suggested that temporal processing depends on a set of neural oscillators that work at distinctive frequencies (Treisman et al. 1992). The preferred frequencies could define the duration specificity for a particular set of intervals (i.e., 450 and 850 ms), and excluding others (i.e., 650 ms), and the period of the oscillator could reproduce the scalar property based on its resolution (Collyer et al. 1994). An alternative is the interval-based model that assumes that different intervals are represented by duration-specific elements (Ivry 1996). This idea was supported by recent neurophysiological experiments from our laboratory, where the neural activity of the supplementary motor cortex was recorded while monkeys performed interval-production tasks. The results not only showed that neurons are tuned to particular interval durations but also indicated that, at the population level, the distribution of preferred intervals was bimodal, with peaks around 450 and 850 ms (Merchant et al. 2008d). In addition, a Gaussian fitting analysis showed that neural tuning increased its dispersion as a function of the preferred interval, following the scalar property of interval timing (Merchant et al. 2008d). Overall, these findings support the idea that the pattern of generalization observed in the present study could depend on an interval-tuning mechanism that can be trained to produce more precise temporal processing. Nevertheless, a differential change in neural network states (Buonomano 2000; Karmarkar and Buonomano 2007) or in the slope of neural tuning curves (Schoups et al. 2001) could produce similar results. Needless to say, more empirical data are needed to validate or reject these ideas.
Many studies have found that an auditory stimulus of a given duration tends to be judged as longer than a visual stimulus of the same duration (Goldstone and Lhamon 1974; Wearden et al. 1998; Grondin 2001). This effect has been attributed to an oscillator that decreases its rate during the visual condition (Wearden et al. 1998). In the present study we found that the visual generalization curve was tilted to the right with respect to the auditory curve. From the perspective of the interval-tuning model, the intensive training for one interval defined by auditory markers increased the quality of temporal information processing in a population of cells tuned to such interval. Thus, the right tilt in the generalization distribution during the visual condition implies that the comparison between the actual visual interval and the memory trace of the auditory interval by the enhanced tuned cells produces a bias in the transfer pattern.
Finally, the comparison between the instruction and internally driven periods during the learning and generalization suggests that the latter task period not only involves procedural and/or sensorimotor learning but also depends on an improvement in the temporal processing resolution. Similar findings were found in an auditory interval-discrimination task that showed that the generalization learning was independent of an overall enhancement in the ability to store and compare stimuli (Karmarkar and Buonomano 2003).
References
Buonomano DV (2000) Decoding temporal information: a model based on short-term synaptic plasticity. J Neurosci 20(3):1129–1141
Buonomano DV, Karmarkar UR (2002) How do we tell time? Neuroscientist 8(1):42–51
Collyer CE, Broadbent HA, Church RM (1992) Categorical time production: evidence for discrete timing in motor control. Percept Psychophys 51(2):134–144
Collyer CE, Broadbent HA, Church RM (1994) Preferred rates of repetitive tapping and categorical time production. Percept Psychophys 55(4):443–453
Fant G, Kruckenberg A (1996) On the quantal nature of speech timing. In: Proceedings of ICSLP 1996, Philadelphia, pp. 2044–2047
Gibbon J, Malapani C, Corby LD, Gallistel CR (1997) Toward a neurobiology of temporal cognition: advances and challenges. Curr Opin Neurobiol 7:170–184
Goldstone S, Lhamon WT (1974) Studies of auditory-visual differences in human time judgement. 1. Sounds are judged longer than lights. Percept Mot Skills 39(1):63–82
Grondin S (2001) From physical time to the first and second moments of psychological time. Psychol Bull 127(1):22–44
Ivry R (1996) The representation of temporal information in perception and motor control. Curr Opin Neurobiol 6:851–857
Ivry RB, Schlerf JE (2008) Dedicated and intrinsic models of time perception. Trends Cogn Sci 12(7):273–280
Karmarkar UR, Buonomano DV (2003) Temporal specificity of perceptual learning in an auditory discrimination task. Learn Mem 10:141–147
Karmarkar UR, Buonomano DV (2007) Timing in the absence of clocks: encoding time in neural network states. Neuron 53:427–438
Kristofferson AB (1980) A quantal step function in duration discrimination. Percept Psychophys 27(4):300–306
Lewis PA, Miall RC (2003) Distinct systems for automatic and cognitively controlled time measurement: evidence from neuroimaging. Curr Opin Neurobiol 13:250–255
Mauk MD, Buonomano DV (2004) The neural basis of temporal processing. Annu Rev Neurosci 27:307–340
Meegan DV, Aslin RN, Jacobs RA (2000) Motor timing learned without motor training. Nat Neurosci 3(9):860–862
Merchant H, Georgopoulos AP (2006) Neurophysiology of perceptual and motor aspects of interception. J Neurophysiol 95:1–13
Merchant H, Zarco W, Prado L (2008a) Do we have a common mechanism for measuring time in the hundreds of milliseconds range? Evidence from multiple-interval timing tasks. J Neurophysiol 99:939–949
Merchant H, Zarco W, Bartolo R, Prado L (2008b) The context of temporal processing is represented in the multidimensional relationships between timing tasks. PLoS ONE 3(9):e3169
Merchant H, Luciana M, Hooper C, Majestic S, Tuite P (2008c) Interval timing and Parkinson’s disease: heterogeneity in temporal performance. Exp Brain Res 184:233–248
Merchant H, Zarco W, Prado L (2008d) Neurophysiology of temporal information processing in the medial premotor areas of the primate. Neuroscience 2008 Annual Meeting. The Society for Neuroscience. Abstract number: 877.3
Merchant H, Zarco W, Prado L, Perez O (2009) Behavioral and neurophysiological aspects of target interception. Adv Exp Med Biol 629:201–220
Moelants D (2002) Preferred tempo reconsidered. In: Stevens C, Burnham D, McPherson G, Schubert E, Renwick J (eds) Proceedings of the 7th international conference on music perception and cognition. AMPS—Causal Productions, Sydney, pp. 580–583
Nagarajan SS, Blake DT, Wright BA, Byl N, Merzenich M (1998) Practice related improvements in somatosensory interval discrimination are temporally specific but generalize across skin location, hemisphere, and modality. J Neurosci 18(4):1559–1570
Schoups A, Vogels R, Qian N, Orban G (2001) Practising orientation identification improves orientation coding in V1 neurons. Nature 412:549–553
Snedecor GW, Cochran WG (1989) Statistical methods. Iowa State University Press, Ames, IA
Treisman M, Faulkner A, Naish PLN (1992) On the relation between time perception and motor action: evidence for a temporal oscillator controlling the timing of movement. Q J Exp Psychol 45A(2):235–263
Wearden JH, Edwards H, Fakhri M, Percival A (1998) Why “sounds are judged longer than lights”: application of a model of the internal clock in humans. Q J Exp Psychol 51B(2):97–120
Westheimer G (1999) Discrimination of short time intervals by the human observer. Exp Brain Res 129:121–126
Wright BA, Buonomano DV, Mahncke HW, Merzenich MM (1997) Learning and generalization of auditory temporal-interval discrimination in humans. J Neurosci 17(10):3956–3963
Acknowledgments
We are grateful to Dean Buonomano, Roberto Prado, and Dagmar Sternad for their fruitful comments on the manuscript. We also thank Dorothy Pless for proofreading the paper and Luis Prado and Raúl Paulín for their technical assistance. Supported in part by PAPIIT: IN206508-19, FIRCA: TW007224-01A1, and CONACYT grant 47170.
Author information
Authors and Affiliations
Corresponding author
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
About this article
Cite this article
Bartolo, R., Merchant, H. Learning and generalization of time production in humans: rules of transfer across modalities and interval durations. Exp Brain Res 197, 91–100 (2009). https://doi.org/10.1007/s00221-009-1895-1
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00221-009-1895-1