
Abstract
The study measured the effect of stretch on passive mechanical properties in unexercised and eccentrically exercised plantarflexor muscles, to obtain insight into how stretch might serve athletes as a warm-up strategy. Passive torque, voluntary contraction strength and muscle soreness were measured before and after a large amplitude stretch given before and after a period of eccentric exercise and at 0, 1, 2 and 24 h later. Stretch of the unexercised muscle led to a 20% fall in passive torque which recovered within an hour. About 40% of the fall could be recovered immediately with a voluntary contraction. After eccentric exercise there was a rise in passive torque by 20% at 2 h post-exercise. This rise was postulated to result from an injury contracture in muscle fibres damaged by the exercise. It was accompanied by a fall in maximum voluntary torque and the development of muscle soreness at 24 h. Stretch of the exercised muscle led to a fall in passive torque and rise in pain threshold. It is proposed that in response to a stretch there is a fall in passive tension in the muscle due to stable cross-bridges in sarcomeres which could be recovered with a voluntary contraction and an additional component attributable to the elastic filament, titin. The size of the fall was not significantly different between exercised and unexercised muscle. These observations provide a physiological basis for the effects of passive stretches on skeletal muscle and help to explain why they are used as a popular warm-up strategy.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
A strategy commonly employed by athletes before a competitive event is to undergo a “warm-up” procedure. Taken literally, the term “warm-up” implies raising body temperature and, more specifically, the temperature of the muscles to be exercised. However warm-up is often referred to more loosely and can include any kind of activity that will prepare muscles for a period of exercise (Bishop 2003a, b). One technique commonly used is to repeatedly stretch the relaxed muscle out to long lengths. Clearly here there is little or no accompanying rise in muscle temperature. This paper is about mechanical changes in muscle after a large-amplitude stretch.
It is a common belief that a stretch will lower passive tension in muscle, thereby increasing joint flexibility and range-of-motion (Toft et al. 1989; Magnusson 1998; Harvey et al. 2002). The issue of lowering passive tension after stretch is potentially important since it may represent a means of reducing muscle injuries. However, at present there is no direct evidence in support of such a claim (Safran et al. 1989; Witvrouw et al. 2004). It has been shown that muscle stiffness levels are a predictor of the amount of muscle damage from eccentric exercise (McHugh et al. 1999). By reducing muscle stiffness, injury susceptibility might be reduced. So if stretches reduced stiffness, it would be an important outcome for eccentrically biased sporting activities. However, there are alternative views. For a review, see McHugh (2003).
We have been studying passive muscle properties from two points of view. The first is muscle thixotropy. We mean by this the dependence of passive mechanical properties on the previous history of contraction and length changes (Proske et al. 1993). We therefore considered that as well as any effects on non-contractile elements in the muscle, one possible consequence of a warm-up stretch was that it lowered muscle passive tension as a result of muscle’s thixotropic property.
Eccentric exercise is accompanied by a number of changes in muscle mechanical properties (Proske and Morgan 2001). We have previously confirmed earlier observations (Jones et al. 1989; Howell et al. 1993; Chleboun et al. 1998) that immediately after a period of eccentric exercise there is a rise in passive tension (Weerakkody et al. 2003a; Whitehead et al. 2001b, 2003). This rise was attributed to tension generated in damaged muscle fibres that had developed an injury contracture as a result of uncontrolled Ca2+ release (Proske and Morgan 2001). Furthermore, this additional passive tension could be partly collapsed by means of a large-amplitude passive stretch (Gregory et al. 2003; Whitehead et al. 2003). All of this raised the possibility that there might be two mechanisms responsible for the fall in passive tension after large amplitude stretches, one attributable to muscle thixotropy, the other acting on above-normal tension levels from exercise-induced muscle damage. In other words, we considered the possibility that some of the beneficial effects reported from the use of warm-up stretches were by individuals who were carrying mild levels of damage from previous eccentric exercises.
In a recent study (Reisman et al. 2005) we reported that after a period of eccentric exercise of elbow flexors, they showed a rise in passive tension, as indicated by a change in resting elbow angle. A series of passive stretches was able to collapse this tension back towards pre-exercise levels. Two important benefits of such stretches given after the exercise were a reduced sense of muscle stiffness and lower levels of delayed-onset muscle soreness (DOMS).
In the present study we have extended those observations. We chose to make measurements on leg muscles rather than arm muscles. Elbow flexors do not routinely undergo high force eccentric contractions whereas ankle plantar flexors and knee extensors do so during running, downhill walking and landing from a jump. It made it more likely that the chosen muscles would, from time to time, exhibit symptoms of damage from unaccustomed eccentric exercise. Furthermore, our earlier work had relied on an indirect estimate of passive tension, inferred from changes in resting elbow angle. Here we wanted to directly measure passive torque. It was hoped to link any changes in passive tension with perceived levels of DOMS. Finally we wanted to further assess the contributions from the above two mechanisms to the fall in tension and to determine whether there was any interaction between them.
Methods
Seventeen young, adult subjects (22.6 ± 3.5 years) 11 female, 6 male participated in the experiments. Each of the two experiment was carried out on ten subjects. It made for a total of 17 since some subjects participated in both experiments. All subjects were in good health, they did not have any previous history of leg injuries and none engaged in regular training programs that involved ankle muscles. Subjects gave their informed, written consent prior to undertaking the experiments, which were all approved by the Monash University Committee for Human Experimentation, and ethical aspects of the experiments conformed to the Declaration of Helsinki.
The equipment
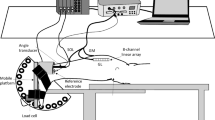
The testing equipment was similar to that described previously (Jones et al. 1997) and consisted of an adjustable chair clamped to a steel frame (Fig. 1). A pair of footplates rotated about an axle aligned with the ankle joint. Two strain gauges were cemented to the shaft of the axle on each side. The strain gauges, connected in a Wheatstone bridge configuration, provided a voltage output proportional to torque about the ankle. Ankle angle was defined as the included angle between foot plate and lower leg. Once a subject’s foot was firmly strapped to the footplate, ankle angle could be changed systematically by rotating the footplate and locking it in position (Fig. 1).

Apparatus used for measurement of passive torque about the ankle joint. Subjects sat in an adjustable chair which was clamped to a steel frame supporting two footplates. Each footplate rotated about an axle, to which were cemented strain gauges for torque measurements. The footplates could be locked in place in 5° increments (120° to 60°included ankle angle). The knee was held in position at ~ 140° included angle by means of a brace. Surface electrodes were placed on triceps surae and EMG was monitored throughout the experiment to ensure subjects remained relaxed during measurements
Measurement of passive tension
At the start of a set of measurements the angles subtended by the footplate, lower leg and upper leg were measured with a digital inclinometer. Position of the chair was adjusted to give an included knee angle of 140° (180° = straight leg). The position of the knee was fixed with a leg brace and kept constant for the remainder of the experiment. Passive torque was measured in 5° increments of ankle dorsiflexion starting from 120° (plantarflexed) up to 60° (dorsiflexed). Provided the subject remained relaxed, as indicated by the absence of any electromyographic activity (EMG), a 30-s segment of torque was recorded at each angle. Because of small fluctuations in torque, an average was calculated over 3 s of record, once stress relaxation had subsided after each change in angle (Fig. 2). Passive torque measurements were gravity corrected at each angle by subtracting the torque due to the weight of the foot and footplate.

Changes in passive torque at different ankle angles in an unexercised muscle. The ankle was rotated in 5° increments from 120° (plantarflexed) to 60° (dorsiflexed). At each angle a 30-s segment of record was recorded after the peak of stress relaxation. Upper pair of traces passive torque, lower trace ankle angle. Bold trace–torque responses under pre-stretch conditions; Faint trace—immediately after the muscle had been subjected to a large stretch over the range 120° to 50°
The electromyogram
Electromyography was recorded using bipolar adhesive surface electrodes on the belly of the triceps surae. The electrodes were silver/silver chloride (3 M Red Dot, Canada). Leads were placed on medial gastrocnemius and on soleus and an indifferent was placed on the tibial head. Voltage output from the EMG electrodes was bandpass filtered (10 Hz–1 kHz) and differentially amplified (Bioamp ML131, ADInstruments, Australia). Torque and EMG data was recorded at 100 Hz using a Maclab 8 s using Chart V3.66 software running on a Macintosh computer (AD Instuments, Australia). EMG biofeedback was provided to subjects using an audio amplifier and speaker.
Experiment 1: stretch of unexercised muscle
Ten subjects participated in this experiment (three male, seven female). Three sets of control measurements of passive torque were carried out on ankle plantar flexors of both legs with at least 6 h allowed to lapse between each set. This was necessary since the act of measurement of passive torque at the longer muscle lengths itself represented a stretch of the muscle which had to subside before another could be made. Subjects were provided with audio feedback of EMG during the measurements. Whenever the ankle was moved to a new position and before the measurement was made, the EMG was inspected to ensure that subjects’ triceps surae had remained relaxed. At each angle a 30-s segment of torque trace was recorded, starting after the peak of the stress relaxation (Fig. 2) The final value at 30 s, measured as the mean over 3 s, was used to determine the torque: angle relationship, and an exponential curve was fitted to values (Fig. 3).

Passive torque-angle curves in an unexercised muscle before and after a stretch. Pooled data (n = 10) showing mean values (±SEM) Exponential curves were fitted to the data. Filled circles with solid line pre-stretch, triangles and dashed line curve immediately after stretch, open circles with dotted line curve 1 h after stretch. The hatched bar at the bottom of the figure indicates the approximate range of ankle angles during walking
After the third control measurement, one leg was randomly designated as the experimental leg and its ankle muscles were stretched by dorsiflexing the ankle to 50°, holding it there for 30 s and then returning it to its resting position. Subjects tolerated a stretch to 50°, although it was close to the limit of the range for ankle extensors. No subject experienced any pain during the stretch. The amplitude, speed (~ 0.035 rad s−1) and duration of the stretch were arranged to approximate what athletes do as part of a routine stretching regime before exercise (Depino et al. 2000). These stretches should be seen as considerably less severe than, for example, those employed by Fowles et al. (2000). Measurements of passive tension were made immediately after the stretch, at 30 min, 1 and 3 h. For the measurements at 30 min the fall in passive tension may have been slightly overestimated since the effect from making the previous set of measurements had not yet fully subsided.
Voluntary contractions
In an attempt to obtain insight into mechanisms underlying the effects of a stretch on passive torque, additional supplementary observations were made on ten subjects. Seven were from the group used for the measurements of passive torque in unexercised muscle, plus a further three were newly recruited subjects. It was reasoned that the fall in torque from the stretch was due to the introduction of slack in muscle fibres. A contraction should take up at least some of the slack. Therefore the effect of a voluntary contraction on the recovery of passive torque was measured after a stretch. To do this, passive torque was measured at 70° before a stretch and immediately afterwards. Then the ankle was rotated back to 120° and a half-maximum contraction was carried out over 3 s. Immediately afterwards the ankle was returned to 70° and passive torque was remeasured. Here the rationale was that at a short muscle length (120°) any slack present was due to muscle thixotropy and it was known that a half-maximum contraction removed such slack (Gregory et al. 1998).
Experiment 2: stretch of eccentrically exercised muscle
The ten subjects from Experiment 1 also participated in this experiment (three male, seven female). Control passive torque and pain measurements were made on three separate occasions before the exercise, 2 on 1 day (separated by at least 2 h), and a third immediately before the exercise. The leg, randomly assigned the experimental leg, was subjected to eccentric exercise for 1 h. The subject’s other leg was used as a control.
The exercise
Subjects carried out the eccentric exercise by walking backwards, downhill, on a forward-moving treadmill (Biodex RTM, 17). The treadmill was inclined at 9° and moved at 2.3 km h−1. Subjects were asked to step backwards with one foot using a toe-to-heel action to ensure that calf muscles were actively stretched during the movement. The other leg was brought alongside and placed flat on the treadmill. It therefore did not undergo any eccentric contractions and could be used as a control for the exercised leg. By this time the subject had been carried back to the top of the treadmill ready for the next step. Subjects carried out 25–30 steps a minute and maintained this for 1 h (1,500–1,800 contractions).
Muscle damage indicators
The main indicators of muscle damage measured in this study were the post-exercise increase in passive torque and the soreness developed at 24 h onwards. An additional measurement carried out on four subjects who had undergone eccentric exercise was the fall in peak active torque immediately after the exercise, and 24 h later. A fall in torque measured shortly after the exercise was considered a less reliable damage indicator than the rise in passive torque because it included a component of metabolic fatigue generated independently of the damage.
Fall in active torque
Subjects were asked to carry out a maximum voluntary isometric contraction (MVC) while ankle angle was held at 70°. It had been shown previously that 70° corresponded to approximately the optimum angle for peak torque (Jones et al. 1997). Its value was checked here by asking subjects to carry out MVCs over a range of angles from 100° to 55°. Optimum angle was 66° ± 2°. Subjects were given verbal encouragement and visual feedback of torque levels during the genesis of each 3 s duration contraction. They were asked to generate three sucessive contractions, separated by 5 min rest periods and the highest torque level reached was designated the value for MVC. Values for MVC were measured immediately before and after the eccentric exercise and 24 h later, in both the exercised and unexercised leg.
Muscle pain
Perceived muscle pain was measured as both a soreness rating and a tenderness rating. Subjects were asked to rate muscle soreness during walking, using a visual analogue scale (0 = no soreness, 10 = worst possible soreness). Before making their assessment subjects were shown a series of descriptors for different pain levels and they were asked to rate their pain using the closest matching descriptor.Tenderness was measured by determining the pressure-induced pain threshold (Weerakkody et al. 2003b). This was done using a compression gauge (Attonic, Japan) with a 15-mm diameter metal disc attached to its shaft. The gauge was pressed into the the muscle and force was recorded when the subject first reported any pain. Pain thresholds of 30 N or more were considered as no pain. Three measurements were made, one in the middle of each of medial and lateral gastrocnemius and a third on soleus. The average of the three values was used as the reported threshold.
Rise in passive torque
Passive torque after eccentric exercise was measured in the same way as described above for the unexercised muscle. Measurements were made on both the exercised muscle and the contralateral control before the exercise, immediately afterwards and at 1, 2, 3 and 24 h post-exercise. At 2 h and at 24 h a stretch to 50° of 30 s duration was given and passive torque was measured immediately afterwards.
Statistical analysis
The data was analysed with a software package (Datadesk 5.0, Data Descriptions, Ithaca, NY, USA). In Experiment 1 repeated measures ANOVAs were used to test the effect of time on passive torque and to assess the effects of a stretch and voluntary contraction on passive torque. In Experiment 2 a repeated measures ANOVA was used to test the effect of time on changes in pain threshold, pain ratings and passive torque in both the experimental and control legs. Where significance was found, an LSD post hoc test was used to test for significance of individual time points. Paired t-tests were used to test for differences in the effects of stretch at 2 and 24 h in both the experimental and control legs. In all tests significance was assigned at P < 0.05. Values are given as mean ± SD (standard deviation) where the data were from individual subjects and mean ± SEM (standard error of the mean) for pooled data from a group of subjects.
Results
Experiment 1: stretch of unexercised muscle
Passive torque and the effect of a stretch on passive torque were measured on ten subjects. Passive torque at 0, 0.5, 1 and 3 h post-stretch decreased to 80.2% (±3.0%), 95.3% (±5.2%) 98.9% (±4.3%) and 102.5% (±4.1%) over these time points respectively (Fig. 4). Example torque traces are shown for one subject in Fig. 2. Immediately after the stretch passive torque values were lower at all angles, the differences becoming more prominent at longer lengths where torque levels were higher. At the longer lengths there was also a more marked stress-relaxation at the end of each stretch. Pooled data for the ten subjects is shown in Fig. 3. Notice passive torque had recovered to near pre-stretch levels by 1 h, post-stretch.

Time-course of recovery of passive torque after stretch (n = 10 subjects). Values shown are mean (±SEM) measured at an ankle angle of 70°. Percentage changes in passive torque after the stretch, expressed relative to the pre-stretch value, given as 100% (dotted line). Measurements were made before, immediately after the stretch (0 h), 0.5 h later and then at 1 and 3 h. Asterisk indicates significant difference from control value
To demonstrate the time course of torque changes more clearly, percentage changes measured at 70° have been shown over the 24 h period (Fig. 4). There was a significant fall in passive torque after stretch over the different time points (two-way repeated measures ANOVA F 4,4 = 4.89, P < 0.05). The LSD post hoc test showed that passive torque immediately after stretch was significantly (P < 0.05) different from the pre-stretch value. Full recovery of passive torque took at least 1 h (Fig. 4).
The effect of a half-MVC on recovery of passive torque at 70° was tested on ten subjects. In this group a stretch to 50° led to a significant reduction in passive torque (84 ± 3%). After a contraction there was a significant recovery of passive torque to 91% (±2%) of the pre-stretch value. That is, 42% of the fall after the stretch had been recovered. However, passive torque after the contraction was still significantly below the pre-stretch value. A repeated measures ANOVA showed that there was a significant difference between the three conditions, pre-stretch torque, post-stretch torque and post-stretch torque after a contraction (F 2,18 = 17.5; P < 0.05). A post hoc LSD test revealed a significant fall in torque after stretch as well as a significant recovery after the contraction.
Experiment 2: stretch of eccentrically exercised muscle
This experiment was carried out on ten subjects. After the exercise all subjects showed a rise in passive torque in ankle extensors of the exercised leg and at 24 h they had developed muscle soreness, indicating that the exercise had led to some muscle damage. In addition, for the four subjects for whom MVC torque measurements had been made post-exercise, torque had fallen significantly.
Fall in active torque
Immediately after the exercise mean peak torque had dropped to 82% (±6%) of the pre-exercise value and by 24 h it had recovered to 91% (±4%). There were no significant changes in peak torque for the contralateral muscle, immediately post-exercise or at 24 h.
Muscle pain
Immediately post-exercise subjects reported a small level of muscle soreness in the exercised leg during slow walking (mean 0.4), but this remained non-significant until the 24 h measurement (mean 4.4 ± 0.7, Fig. 5). There was no persisting soreness when subjects sat with leg muscles relaxed. There was no soreness at any time in the unexercised muscle of the other leg.

Delayed-onset muscle soreness after eccentric exercise. Upper panel reported levels of pain during walking before a period of eccentric exercise (Pre-ex), immediately afterwards (0 h) and at 1, 2, 3 and 24 h after the exercise. Pain was scored using a Visual Analogue Scale of 0–10 in which 0 was no soreness and 10 was worst possible soreness. All values are mean (±SEM) for ten subjects. Asterisks indicate a significant change in values from control or pre-stretch. Lower panel changes in pain threshold (N) in response to local compression of the muscle (mean ± SEM for ten subjects). A threshold of 30 N or more (pre-ex) was considered as no pain. Immediately after the exercise (0 h) and at 1 h there was no change in threshold. A small fall took place at 2 h and a larger one at 24 h. A large stretch at 24 h significantly raised threshold. Asterisks indicate a significant change in values from control or pre-stretch
A repeated measures ANOVA showed that there was a significant effect of time on soreness in the exercised leg (F 7,63 = 24.15, P < 0.05). Further analysis showed that soreness at 24 h was above pre-exercise levels (LSD post hoc test).
Measurements of pain threshold in response to muscle compression showed a slight, non-significant fall at 2 and 3 h after the exercise. There was, however, a further, significant fall in threshold at 24 h (Fig. 5). Compression threshold had fallen by an average of 6.7 N. A Repeated measures ANOVA showed that there was a significant effect of time on pain threshold (F 7, 63 = 156.8).
At 24 h post-exercise, pain measurements were made before and after a stretch (Fig. 5). After the stretch muscle soreness had fallen significantly (2.6 ± 0.7; LSD post hoc test). Similarly pain threshold was significantly higher (25.6 ± 1.8 N) than it had been before the stretch (23.2 ± 1.5 N; LSD post hoc test). However pain ratings remained significantly above pre-exercise levels, even after a stretch, while compression pain threshold after a stretch remained significantly below the pre-exercise value.
Passive torque
After the exercise passive torque in the experimental leg was found to have risen in all subjects. The changes in passive torque at 2 h post-exercise for the full range of ankle angles and the effect of a stretch are shown in Fig. 6. The percentage changes for the group at 70° are shown in Fig. 7. Passive torque immediately after the exercise rose to 109% (±6%, Fig. 7). It continued to rise to peak at 2 h post-exercise at 120% (±5%). A stretch at 2 h post-exercise reduced passive torque to 94% (±3%), of the control value. It then recovered and by 24 h was back to 116% (±4%). A stretch at 24 h lowered passive torque back to 96% (±4%, Fig. 7).

Passive angle-torque curves before and after exercise. Values are mean (±SEM) for ten subjects. Exponential curves have been fitted to each set of data. Filled circles with continuous line, curve measured before the exercise (pre-ex); open squares and dashed line curve measured at 2 h post-exercise (2 h); open circles, dotted line curve measured at 2 h immediately after applying a large passive stretch (2 h + stretch). Hatched bar approximate range of ankle angles during walking

Time course of passive torque changes after eccentric exercise, before and after stretch of the exercised muscle (filled circles, solid line) and the unexercised contralateral muscle (open circles, dashed line). Values are mean (±SEM) for ten subjects. Passive torque (measured at 70° ankle angle) is expressed as a percentage of the pre-exercise control value, assigned 100%. Measurements were made immediately before the exercise (pre-ex), immediately afterwards (0 h), at 1, 2, 3 and 24 h post-exercise. Asterisks indicate values significantly different from control or pre-stretch
Passive torque in ankle plantarflexors of the control leg did not rise significantly after the exercise. However a stretch reduced passive torque from 105 to 84% (±2.6%) of control values followed by recovery to 97% (±5.5%) at 24 h. A second stretch at this point lowered passive torque to 79% (±3.5%, Fig. 7).
Statistical analysis showed a significant main effect of time (F 7,63 = 4.9, P < 0.05) on passive torque in the exercised leg. Post hoc analysis showed passive torque at 2 h and at 24 h was significantly higher than pre-exercise torque. Despite the higher passive torque in the eccentrically exercised leg, the amount by which the stretch at 24 h reduced passive torque was not significantly different from that for the unexercised leg. Paired t-tests showed that for the exercised muscle, at 2 h post-exercise the fall in torque after the stretch (26%) was not significantly different from the fall seen in the control muscle (21%). Similarly at 24 h after the exercise the fall in the exercised muscle (20%) was not significantly different from that in the control muscle (18%).
Discussion
Experiment 1: stretch of unexercised muscle
In the first experiment we demonstrated that a large passive stretch of ankle extensors led to a 20% reduction in passive torque. Torque took an hour to recover back to control values (Fig. 4). However an MVC immediately after the stretch recovered 40% of the fall.
Based on animal experiments, we have considered passive tension in skeletal muscle to comprise three components, resting tension, short-range tension and elastic tension (Whitehead et al. 2001a). Resting tension, also called filament resting tension (Hill 1968) and short-range tension, which together comprise muscle thixotropy, are attributed to the presence of weakly-attached, stable cross-bridges between myofilaments of passive muscle fibres (Proske and Morgan 1999). Stretch of passive muscle leads to detachment of stable cross-bridges and their re-formation at longer lengths. The fibres, stiffened by the presence of bridges that have attached at the longer length, fall slack when the muscle is returned to its original length (Proske et al. 1993). We mean by slack that the straight-line distance between the attachment points at each end of the muscle is less than the actual length of muscle fibres. Given that triceps surae has such a long tendon it is probable that for this muscle group some of the slack will be in the tendon as well as in muscle fibres. A subsequent stretch or contraction must take up any slack before tension can begin to rise.
The third component of passive tension, the elastic tension, at short muscle lengths is not dependent on the previous history of length changes. However, at longer lengths, beyond the optimum length for peak active tension it, too, becomes history dependent (Whitehead et al. 2001a). A structural element within the sarcomere which may be responsible for elastic tension is the elastic filament titin. It connects the myosin molecule in each half-sarcomere to the z-line and is known to be responsible for a proportion of the passive tension in muscle (Horowits 1992; Linke et al. 1996). The history-dependence of passive tension attributed to the titin molecule may relate to its unfolding and re-folding property (Wang et al. 1993). In other words, at longer lengths passive tension after a stretch can fall, both due to development of slack from thixotropy and from a fall in elastic tension.
We propose that some of the fall in passive torque in ankle extensors after passive stretch is due to the development of slack in muscle fibres and tendon. Theory predicts that active cross-bridge cycling during a contraction will take up any pre-existing slack and therefore, once the muscle relaxes, the new level of passive tension will be higher. In the experiments reported here, although a voluntary contraction after a stretch increased passive tension, it did not fully return it to control levels. Our interpretation of this result is that the contraction removed the component of slack due to thixotropic properties but not that attributable to a fall in elastic tension. Full recovery took at least an hour (Fig. 4). Here our interpretation is that recovery of elastic tension, involving re-folding of the titin molecule would not be expected to be affected by an MVC and for the rabbit psoas muscle it took 20 min for fibres to recover their original sarcomere length (Wang et al. 1993).
It has recently been claimed that titin is Ca2+ sensitive and that following active stretches of muscle fibres on the descending limb of their length-tension relationship, passive tension is increased as a result of the contraction-generated rise in Ca2+ acting on titin (Rassier et al. 2005). Such a mechanism could potentially explain the recovery of passive tension after a stretch using a voluntary contraction. However, in the present study the voluntary contractions were generated at relatively short muscle lengths (120°) where it is generally agreed titin makes little or no contribution to passive tension.
In animal experiments we have shown that recovery of short-range tension occurs spontaneously and takes about 60 s (Whitehead et al. 2001b). Similar values have been reported for human ankle muscles (Hufschmidt and Schwaller 1987) and finger muscles (Lakie and Robson 1988). The fact that in the present experiments passive torque took nearly an hour to return to pre-stretch levels adds support to the view that additional factors were playing a role. It is interesting that in animal experiments the recovery of tension after a stretch following a series of eccentric contractions also took an hour (Whitehead et al. 2003). While it remains uncertain what might be responsible for such a slow time course, one possibility is the elastic tension attributed to titin (Wang et al. 1993).
Experiment 2: stretch of eccentrically exercised muscle
Evidence of damage
In this study three damage indicators were measured after the eccentric contractions. These were a fall in MVC, muscle soreness (DOMS) and rise in passive torque (see Proske and Morgan 2001; Proske and Allen 2005 for reviews). Fall in MVC as a damage indicator is complicated by the effects of fatigue. All forms of exercise lead to fatigue immediately afterwards, but not necessarily to damage. However recovery from fatigue would be expected to be complete at 1–2 h, so that any force deficits persisting at 24 h would be expected to be entirely due to damage (Smith and Newham 2007). For the four subjects where MVC was measured, this had fallen to 82% of control values immediately after the exercise and had recovered to 91% by 24 h. So the remaining shortfall of 9% suggested that some damage had occurred.
The second damage indicator was DOMS, which had developed by 24 h. It was measured as a muscle soreness rating and as pain threshold in response to local muscle compression (Fig. 5). DOMS typically persisted for 4 days post-exercise (Weerakkody et al. 2003b).
The third damage indicator was a rise in whole muscle passive tension post-exercise, measured as ankle torque in the absence of EMG. This had increased by 9% immediately after the exercise and continued to rise over the next 2 h to reach 20% above control levels. These observations are consistent with data from our own observations on human and animal muscles (Reisman et al. 2005; Whitehead et al. 2001b, 2003). Similar findings have been reported by other groups (Jones et al. 1987; Howell et al. 1993; Chleboun et al. 1998)
It has been hypothesised (Morgan 1990) that sarcomeres become disrupted during eccentric contractions. That, in turn, leads to membrane damage, uncontrolled release of Ca2+ and development of an injury contracture (Whitehead et al. 2003). The contracture is a dynamic process with sarcomeres in a damaged region shortening as a result of active cross-bridge cycling, thereby stretching adjacent sarcomeres and membranes, leading to a gradual spread of the contracture. As a result, the level of passive tension in the muscle increases with time, over a period of up to 2 h after the exercise (Fig. 7).
Effect of stretch
Inspection of Fig. 7 shows that passive torque in ankle extensors fell significantly after stretch of the exercised muscle, as measured at 70°. Interestingly the fall (26%) was only a little more than that observed in unexercised muscle (21%). Given that passive torque had risen by 20% post-exercise, it meant that at the end of the stretch passive torque was only a little below pre-exercise, pre-stretch levels. However it was well above levels measured in the muscle of the other leg after it had undergone a similar stretch. It is concluded that the presence of extra passive tension in the exercised muscle does not lead to a proportionately larger fall in tension in response to a stretch when compared with the fall in an unexercised muscle. A similar result had previously been obtained for elbow flexors (Reisman et al. 2005).
In animal experiments, in an extensively damaged muscle following a series of eccentric contractions (45% loss of force), one possible mechanism considered for the fall in passive tension after a large stretch was that the stretch increased the number of disrupted sarcomeres, sheared t-tubules and damaged muscle membranes leading to an increase in muscle compliance (Gregory et al. 2007). Given that in the present experiments the fall in active torque was only 18% immediately after the exercise, the amount of damage present was likely to be relatively small. It made it less likely that the fall in torque after a stretch was due to aggravation of the eccentric damage. It led us to postulate that some of the fall in passive torque produced by a stretch could have its origin in the uninjured portions of the muscle which would show thixotropic behaviour, much like the unexercised muscle. In fact the fall in torque after the stretch in the exercised muscle at 2 h and at 24 h was about 12% greater than in the unexercised muscle (Fig. 7). It suggested that mechanisms operating in both damaged and undamaged portions of the muscle were involved, although the contribution from the damaged parts of the muscle was likely to be small.
Both subjectively perceived soreness during walking and muscle tenderness, measured as pain threshold during muscle compression had reached significant levels by 24 h post-exercise (Fig. 5). Pain threshold in response to muscle compression had fallen by 6.7 N at 24 h post-exercise representing a 22% fall. Following a passive stretch, pain threshold increased by 2.2 N, that is, by about a third compared with its value before the stretch. Therefore a passive stretch does represent a useful strategy for reducing muscle pain. It should be remembered, though, that the act of carrying out the stretch will by itself be painful for the subject and if any eccentric damage is present, risks exacerbating the damage. In addition, pain threshold after a stretch will be higher only for as long as passive tension remains low. As soon as a voluntary contraction occurs almost half of the tension is recovered, leading to a lowering of pain threshold. A passive stretch as a means of reducing DOMS should therefore be seen as essentially a short-term strategy, but a useful one at the onset of exercise.
Wider considerations
The findings presented here confirm our earlier observations that passive tension in muscle can be reduced by a stretch (Reisman et al. 2005). The new data has shown that some of that tension can be rapidly recovered with a voluntary contraction. In addition, a stretch does not have a very much larger effect on an eccentrically exercised muscle in which passive tension levels are higher than normal, as a result of muscle damage. The most likely explanation for this finding is that the reduction in tension from the stretch in the exercised muscle is a result of the development of slack, which manifests itself predominantly in the passive parts of the muscle, not the regions undergoing an injury contracture. Stretch of a damaged muscle was, nevertheless, beneficial for subjects since it led to a significant rise in pain threshold and to a fall in passive tension to levels seen in undamaged muscle. Thus our observations confirm that passive stretches are likely to benefit all athletes who want to reduce sensations of stiffness in unexercised muscle or sensations of stiffness and soreness in muscle that has been damaged following a period of eccentric exercise. It remains an open question whether reducing passive tension in a muscle in this way also reduces the risk of injury.
References
Bishop D (2003a) Warm up I: potential mechanisms and the effects of passive warm up on exercise performance. Sports Med 33:439–454
Bishop D (2003b) Warm up II: performance changes following active warm up and how to structure the warm up. Sports Med 33:483–498
Chleboun GS, Howell JN, Conatser RR, Giesey JJ (1998) Relationship between muscle swelling and stiffness after eccentric exercise. Med Sci Sports Exerc 30:529–535
Depino GM, Webright WG, Arnold BL (2000) Duration of maintained hamstring flexibility after cessation of an acute static stretching protocol. J Athl Train 35:56–59
Fowles JR, Sale DG, MacDougall JD (2000) Reduced strength after passive stretch of the human plantarflexors. J Appl Physiol 89:1179–1188
Gregory JE, Wise AK, Wood SA, Prochazka A, Proske U (1998) Muscle history, fusimotor activity and the human stretch reflex. J Physiol 513:927–934
Gregory JE, Morgan DL, Proske U (2003) Tendon organs as monitors of muscle damage from eccentric contractions. Exp Brain Res 151:346–355
Gregory JE, Morgan DL, Allen TJ, Proske U (2007) The shift in muscle’s length-tension relation after exercise attributed to increased series compliance. Eur J Appl Physiol 99:431–441
Harvey L, Herbert RD, Crosbie J (2002) Does stretching induce lasting increases in joint ROM? A systematic review. Physiother Res Int 7:1–1310
Hill DK (1968) Tension due to interaction between the sliding filaments in resting striated muscle. The effect of stimulation. J Physiol 199:637–684
Horowits R (1992) Passive force generation and titin isoforms in mammalian skeletal muscle. Biophysical J 61:392–398
Howell JN, Chleboun G, Conatser R (1993) Muscle stiffness, strength loss, swelling and soreness following exercise-induced injury in humans. J Physiol 464:183–196
Hufschmidt A, Schwaller I (1987) Short-range elasticity and resting tension of relaxed human lower leg muscles. J Physiol 391:451–465
Jones DA, Newham DJ, Clarkson PM (1987) Skeletal muscle stiffness and pain following eccentric exercise of the elbow flexors. Pain 30:233–242
Jones DA, Newham DJ, Torgan C (1989) Mechanical influences on long-lasting human muscle fatigue and delayed-onset pain. J Physiol 412:415–427
Jones C, Allen T, Talbot J, Morgan DL, Proske U (1997) Changes in the mechanical properties of human and amphibian muscle after eccentric exercise. Eur J Appl Physiol Occup Physiol 76:21–31
Lakie M, Robson LG (1988) Thixotropic changes in human muscle stiffness and the effects of fatigue. Q J Exp Physiol 73:487–500
Linke WA, Ivemeyer M, Oliveiri N, Kolmerer B, Ruegg JC, Labeit S (1996) Towards a molecular understanding of the elasticity of titin. J Mol Biol 261:62–71
Magnusson SP (1998) Passive properties of human skeletal muscle during stretch manoeuvres. Scand J Med Sci Sports 8:65–77
McHugh MP (2003) Recent advances in the understanding of the repeated bout effect: the protective effect against muscle damage from a single bout of eccentric exercise. Scand J Med Sci Sports 13:88–97
McHugh MP, Connolly DA, Eston RG, Kremenic IJ, Nicholas SJ, Gleim GW (1999) The role of passive muscle stiffness in symptoms of exercise-induced muscle damage. Am J Sports Med 27:594–599
Morgan DL (1990) New insights into the behavior of muscle during active lengthening. Biophys J 57:209–221
Proske U, Allen TJ (2005) Damage to skeletal muscle from eccentric exercise. Exerc Sport Sci Rev 33:98–104
Proske U, Morgan DL (1999) Do cross-bridges contribute to the tension during stretch of passive muscle? J Muscle Res Cell Motil 20:433–442
Proske U, Morgan DL (2001) Muscle damage from eccentric exercise: mechanism, mechanical signs, adaptation and clinical applications. J Physiol 537:333–345
Proske U, Morgan DL, Gregory JE (1993) Thixotropy in skeletal muscle and in muscle spindles: a review. Prog Neurobiol 41:705–721
Rassier DE, Lee D-J, Herzog W (2005) Modulation of passive force in single skeletal muscle fibres. Biol Lett 1:342–345
Reisman S, Walsh LD, Proske U (2005) Warm-up stretches reduce sensation of stiffness and soreness after eccentric exercise. Med Sci Sports Exerc 37:929–936
Safran MR, Seaber AV, Garrett WE Jr (1989) Warm-up and muscular injury prevention. An update. Sports Med 8:239–249
Smith IC, Newham DJ (2007) Fatigue and functional performance of human biceps muscle following concentric or eccentric contractions. J Appl Physiol 102:207–213
Toft E, Espersen GT, Kalund S, Sinkjaer T, Hornemann BC (1989) Passive tension of the ankle before and after stretching. Am J Sports Med 17:489–494
Wang K, McCarter R, Wright J, Beverly J, Ramirez-Mitchell R (1993) Viscoelasticity of the sarcomere matrix of skeletal muscles. The titin-myosin composite filament is a dual-stage molecular spring. Biophys J 64:1161–1177
Weerakkody NS, Percival P, Morgan DL, Gregory JE, Proske U (2003a) Matching different levels of isometric torque in elbow flexor muscles after eccentric exercise. Exp Brain Res 149:141–150
Weerakkody NS, Percival P, Hickey MW, Morgan DL, Gregory JE, Canny BJ, Proske U (2003b) Effects of local pressure and vibration on muscle pain from eccentric exercise and hypertonic saline. Pain 105:425–435
Whitehead NP, Gregory JE, Morgan DL, Proske U (2001a) Passive mechanical properties of the medial gastrocnemius muscle of the cat. J Physiol 536:893–903
Whitehead NP, Weerakkody NS, Gregory JE, Morgan DL, Proske U (2001b) Changes in passive tension of muscle in humans and animals after eccentric exercise. J Physiol 533:593–604
Whitehead NP, Morgan DL, Gregory JE, Proske U (2003) Rises in whole muscle passive tension of mammalian muscle after eccentric contractions at different lengths. J Appl Physiol 95:1224–1234
Witvrouw E, Mahieu N, Danneels L, McNair P (2004) Stretching and injury prevention: an obscure relationship. Sports Med 34:443–449
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Reisman, S., Allen, T.J. & Proske, U. Changes in passive tension after stretch of unexercised and eccentrically exercised human plantarflexor muscles. Exp Brain Res 193, 545–554 (2009). https://doi.org/10.1007/s00221-008-1657-5
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00221-008-1657-5