
Abstract
Perception of tilt (somatogravic illusion) in response to sustained linear acceleration is generally attributed to the otolithic system which reflects either a translation of the head or a reorientation of the head with respect to gravity (tilt/translation ambiguity). The main aim of this study was to compare the tilt perception during prolonged static tilt and translation between 8 and 20° of tilt relative to the gravitoinertial forces (i.e., G and GIF, respectively) when the semicircular cues were no more available. An indirect measure of tilt perception was estimated by means of a visual and kinesthetic judgment of the gravitational horizon. The main results contrast with the interpretation regarding the tilt/translation ambiguity as the same orientation relative to the shear forces G for the true tilt or GIF in the centrifuge did not induce the same horizon perception. Visual adjustment and arm pointing in the centrifuge were always above the ones observed in a G environment. Part of the lowering of the judgment in the centrifuge may be related to the mechanical effect of GIF on the effectors as shown by the shift of the egocentric coordinates in the direction of GIF. The role of the extravestibular graviceptors in the judgment of the degree of tilt of one’s own body relative to G or GIF was discussed.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
Perception of the orientation of the body and its movements is achieved through the integration of information provided by several sensory systems. Body movements stimulate the sense organs, yielding sensory signals (e.g., signals from the semicircular canals, otolithic organs, somesthetic and visual systems) that encode motion and orientation. Problems arise when humans use assisted means of locomotion, such as aircraft’s, space vehicles and ships and are exposed to motion having angular and linear accelerations which can differ substantially in direction, intensity or frequency from those normally experienced on the ground. Different kinds of erroneous perception of position and movement have been described, which are embraced within the broad definition of spatial disorientation (Benson 1990). One of these erroneous perceptions is the somatogravic illusion (Benson 1990; Clark and Graybiel 1949b; Clement et al. 2001; Cohen 1977): Submitted to a linear acceleration and in the absence of relevant visual cues, subjects feel tilted in the direction opposite to the acceleration. This illusion is usually accompanied by visual effects, the so-called oculogravic illusion (Clark and Graybiel 1949a; Graybiel 1952). Visually perceived objects appear to be above their true physical position (Cohen 1973; Cohen et al. 2001; Graybiel 1952; Schöne 1964; Whiteside et al. 1965). Experimentally, the gravitoinertial modifications are usually engendered by means of a centrifuge. The visual mislocalization of a target set on an horizontal plane passing through the eyes, that is, perceived to be perpendicular to the direction of gravity (visually perceived eye level: VPEL) generally defines the magnitude of the illusion (DiZio et al. 1997; Li and Matin 1993, 1995; Matin and Li 1992, 1994, 1995).
This tilt illusion, which has been shown for very low radial accelerations (0.01 m s−2; Raphel and Barraud 1994; Raphel et al. 1996), is generally attributed to the otolith system, which responds to the gravitational forces relative to the head (Fernandez and Goldberg 1976; Loe et al. 1973) and to the inertial forces produced during linear motion (Fernandez and Goldberg 1976). When accelerated, the effective stimulus sent to the otoliths is proportional to the vector sum of these two forces. Both the angle and amplitude of the resultant (GIF) are altered relatively to gravity. Unfortunately, the otoliths which are inertial sensors cannot differentiate between the vertical linear acceleration of gravity and other linear accelerations acting on the body. Then, tilt illusion may arise because the angle of GIF with respect to gravity (G) is similar to the one generated by a head tilt (Paige and Tomko 1991).
The ability of the central nervous system (CNS) to solve this tilt/translation ambiguity is critical to provide an accurate representation of spatial orientation and to elicit the appropriate sensorimotor reflexes (Seidman et al. 1998). It has been suggested that the perception of tilt or translation is governed by the implementation of internal models. GIF resolution based the implementation of the internal model on the canal–otolith interaction (Angelaki and McHenry 1999; Merfeld et al. 2001; Merfeld and Zupan 2002). During head tilt, information issued from the semicircular canal is coplanar and synergistic with otolith stimulation, while during head translation canal and otolith stimuli are orthogonal (Seidman et al. 1998). However, under usual conditions the ambiguity is mainly resolved by the use of the semicircular canal cues (Angelaki et al. 1999); it implies that the CNS cannot make a perceptual difference between static tilt and prolonged centrifugation. Indeed, the vestibular system and its related perceptions have the remarkable ability to “forget” start-up cues after a sufficient time has elapsed (Holly and McCollum 1996). The canal response decays gradually after the initial angular acceleration ends. For example, subjects estimate angular velocity as zero when seated and spinning on-axis for approximately 50 s after a 10-s acceleration at 24° s-² (Brown 1966).
In the absence of a canal–otolith interaction, it has been stated that a separation of the GIF may occur based on the phasic irregular otolithic afferent signal, interpreted primarily as a linear acceleration and the tonic regular otolithic afferent signal representing gravity (Droulez and Darlot 1989; Mayne 1974). However, it has been shown that primary otolith afferent neurons provide identical responses during head tilts relative to G and during translational movements (Angelaki and Dickman 2000; Dickman et al. 1991; Fernandez and Goldberg 1976; Fernandez et al. 1972; Loe and Werner 1973; Si et al. 1997). Another factor which may contribute to solve the apparent ambiguity is the difference in the saccular and utricular magnitude shear force stimulation between true and simulated tilt (Curthoys et al. 1999; MacDougall et al. 1999). Finally, assuming a neural representation of the gravity vector ( \( \ifmmode\expandafter\vec\else\expandafter\vec\fi{\ifmmode\expandafter\hat\else\expandafter\^\fi{g}}\); Angelaki et al. 1999; McIntyre et al. 2000; Merfeld et al. 2001, 2005a, b; Merfeld and Zupan 2002; Zupan et al. 2002), \( \ifmmode\expandafter\vec\else\expandafter\vec\fi{\ifmmode\expandafter\hat\else\expandafter\^\fi{g}} \) could be viewed as a gauge developed and enhanced with spatial experience (Carriot et al. 2004, 2005; Long et al. 1975). The tilt/translation resolution would then rely on the comparison of the magnitude of the resultant force and of \( \ifmmode\expandafter\vec\else\expandafter\vec\fi{\ifmmode\expandafter\hat\else\expandafter\^\fi{g}} \) and on the evaluation of the angle between the force vector and \( \ifmmode\expandafter\vec\else\expandafter\vec\fi{\ifmmode\expandafter\hat\else\expandafter\^\fi{g}}\). However, the way observers judge the orientation of the objects relative to G also depends on their body tilt estimation relative to \( \ifmmode\expandafter\vec\else\expandafter\vec\fi{\ifmmode\expandafter\hat\else\expandafter\^\fi{g}} \). The perception of the body z-axis has been shown to be influenced by the GIF (Prieur et al. 2005). This misperception of the egocentric coordinates would induce errors in the perceptual judgment of an object relative to the geocentric information.
The main goal of this study was to compare the tilt perception during prolonged static tilt and translation between 8 and 20° of tilt relative to the gravitoinertial forces (i.e., G and GIF, respectively). An indirect measure of tilt perception was estimated by means of a visual judgment (VPEL). The VPEL requires to adjusting the perceived gravitational horizon direction to the eye level. The perception of the body z-axis was also investigated. The transverse plane of the eyes (TPE; Howard and Templeton 1966) was used as an indirect measure of the body z-axis perception. The TPE is a purely egocentric referenced task. It needs to set a target on the plane passing through the socket of the eye which is subjectively perceived as perpendicular to the body z-axis.
It has been shown that the subjects were less influenced by an illusion when the perceptual judgment of the egocentric horizon involved the use of whole limb movements (Tremblay and Elliott 2003). Arm-lifting movements provide information about orientation in space by generating additional cues about the direction of gravity (Guedry et al. 1978). For instance, the dynamic gravitational torque generated by arm movements may be involved in limb position sense in space (Bock 1994; Gooey et al. 2000; Worringham and Stelmach 1985). It may improve a more general exocentric perception about the direction of gravity (Fitger 1976; Gentaz and Hatwell 1996; Luyat et al. 2001). In parallel, a recent study demonstrated that during a body tilt, horizon perception assessed visually and motor-kinesthetically might depend on the same cognitive process consisting in a more or less pronounced shift from an exocentric frame of reference to a more egocentric one (Bringoux et al. 2004). This interpretation is supported by a work suggesting that the ego- and exocentric frames of reference are preexisting neurophysiological structures between which subjects could switch easily, depending on the task demand (Ghafouri et al. 2002). Our last objective was to investigate whether the ratio between the visual (VPEL) and the motor-kinesthetic judgments (subjective proprioceptive horizon: SPH) which exists during true tilt was maintained during simulated tilt.
Methods
Subjects
Fifteen right-handed subjects (8 males and 7 females; age ranged from 19 to 40 years) participated in this study. They had normal vision and were free of any apparent vestibular disorder. All of them were naive about the purpose of the experiment and gave informed consent in compliance with the Huriet Law (i.e., Helsinki Convention), which governs and regulates human experimentation in France.
Apparatus
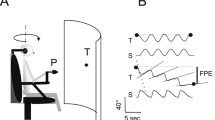
Two basic apparatus was used. The first one was a centrifuge that could rotate at a constant velocity between 0 and 170° s−1. Subjects sat in a cabin in order to sense no airflow during motion. They faced the yaw axis of rotation; the external auditory meatus was distant 50 cm from the axis of rotation (Fig. 1, upper panels). When the centrifuge rotated, the gravitational–inertial resultant force GIF applied on the subjects was the vectorial sum of the radial horizontal acceleration γr combined with gravity G. As a result, GIF was greater than G and tilted in pitch with respect to the vertical. The second apparatus was a “tilt chair”, capable of rotating to any angle in the sagittal plane. As in the centrifuge, subjects sat upright facing the axis of rotation; the external auditory meatus was distant 50 cm from the axis of rotation. (Fig. 1, lower panels). In both the apparatuses, subjects were restrained in the chair by means of two straps fixed at the level of the abdomen and shanks. The head position was held constant with respect to the body axis by the use of a helmet and a neck rest fixed on the chair. Whatever the motion (centrifugation or tilt), the feel of the apparatus was almost absent because of the high level technical characteristics of the motors. In addition, the noise of the motion was maintained constant by use of ear phones providing a white noise signal, such that subjects were unaware of the experimental manipulation which took place in the centrifuge or in the tilt chair.

The apparatus consisted of a centrifuge (upper panels a and b) and a tilting chair (lower panels c and d) in which subjects took place. The illustrations show the distance between the axis of rotation (A1) and the axis of the subjects’ ear (A2). The physical forces acting onto the subjects were represented by F x (dorso-ventral shear force), F z (force along the body z axis) and R (resultant gravitoinertial force); α was the angle between body z-axis and the resultant force. In the centrifuge, F x is the radial acceleration γr, F z the gravity vector and R represents GIF. In the tilting chair, R represents the gravity vector. The theoretical egocentric and horizon localizations for the two experimental situations are also indicated. For the visual judgment tasks (left panels a and c), the electronic plotter board, the luminous target as a light-emitting diode (T) and the joystick (J) are illustrated. The arm outstretched and guided by an aluminium pipe inserted in the rail parallel to the body z-axis which was used in the kinesthetic judgment tasks is also represented (right panels b and d)
In the visual tasks (Fig. 1, right panels), a luminous target, which was a yellow light-emitting diode (LED) fixed on the pencil holder of an electronic plotting board, was positioned 50 cm in front of the subjects in a vertical position. Its accuracy of displacement was ±0.05 mm. The brightness of the LED (2.82 cd m-² measured by a spectrascam 650) was adjusted to be perceptible without dark accommodation. All the elements of the basic apparatus and the electronic plotting board were painted black and installed in a dark room so that, when the LED was switched on, subjects did not perceive any visual reference. Subjects could move the LED “up” or “down” using a joystick.
In the kinesthetic tasks (Fig. 1, left panels), subjects’ right arm was maintained outstretched and guided by means of a very thin and light aluminum black pipe. The pipe was tapped on the arm and on the forefinger. The pipe tip overhung the forefinger of about 20 cm and was inserted in a rail parallel to the body z-axis. Subjects could only move the arm in the sagittal plane. In the starting position, the arm was laid on a starting push button. In the left hand, subjects held a validation push button. Three infrared-emitting markers were placed at different levels of the body: right hand (first phalanx of the index), right shoulder (acromion), right hip (iliac crest). An acquisition system (Optotrak System Northern Digital Inc.) recorded the pointing movements at a sampling frequency of 100 Hz. Data were filtered with a Butterworth filter (second order, 10 Hz cutoff frequency). All the elements were painted black.
The LED motion on the electronic plotting board, the Optotrak system, the centrifuge rotation and the “tilt chair” were computer controlled and monitored.
Experimental design
Each subject performed eight separate sessions presented randomly: VPEL, TPE, SPH and TPA in the two experimental apparatus (centrifuge and tilt chair). Three days, at least, separated each session. Each session included four angular tilts in pitching: 0 (upright position), 8.83, 13.64 and 19.26°, presented in a growing order of angular tilt. With respect to G, these angles corresponded to the orientation of GIF in the centrifuge or to a real body tilt in the “tilt chair”. Data for the various rotation velocities are summarized in Table 1.
Procedure
In the horizon perception task, subjects were instructed to set the luminous target (visual task, VPEL) or the arm (kinesthetic task, SPH) to the location perceived as the line passing through the eye lenses (VPEL) or through the shoulder (SPH) and parallel to the floor of the laboratory. In the egocentric task, subjects were instructed to set the luminous target (visual task, TPE) or the arm (kinesthetic task, TPA) to the location perceived as a line passing through the socket of the eyes (TPE) or through the shoulders (TPA) and perpendicular to the head–body axis representative line.
Just before the session, subjects were instructed to use the joystick (visual task) or the pushes buttons (kinesthetic task). During the visual task, once subjects were installed in the experimental apparatus (centrifuge or tilt chair), the middle of the plotter (zero on the xy axis) was brought into alignment with the horizontal, median plane through the bridge of the nose (between the eyes). Then, room lights were switched off and 2 min after subjects started by making ten settings, while remaining upright (tilt chair) or motionless (centrifuge). In the visual task, for each setting, the target was switched off and randomly displaced on the y-axis in the periphery of the electronic plotting board. The LED was then switched on and a pip sound gave the signal to set the target and to validate the setting with a trigger on the joystick. Then, the LED was switched off again and displaced to the next preset position. In the kinesthetic task, subjects were eyes closed during all the experiment. A pip sound gave the signal to set the arm and to validate the setting with the left-hand push button. Once the settings in motionless or upright position were realized, one of the eight experimental tasks was performed. In the centrifuge, the motor was engaged and the centrifuge was started up slowly (acceleration 2° s−2) until it rotated at 100° s−1 which corresponded to an 8.82° angle. After 2 min of rotation at constant velocity, subjects were asked to make the ten settings under the given gravitational–inertial condition. Then, the centrifuge was accelerated to the next velocity. At the end of the 150° s−1 (19.26°) velocity the centrifuge was stopped gradually. In the tilt chair, subjects were tilted backward slowly with a 3° s−1 constant velocity and a starting acceleration and final deceleration of 0.001° s−2, until 8.93°. After 2 min of stable tilt, subjects were asked to make the ten settings. Then, the chair was tilted to the next position. At the end of the 19.26° of tilt, the chair returned to the initial upright position.
Data collection
The adjustment was expressed either in degrees of visual angle relative to the center of the plotter or in degrees of arm–trunk angle. In accordance with experiments showing a motor adaptation to the coriolis and gravitoinertial forces (Lackner and DiZio 2003; after four settings for the study of Coello et al. 1996), the five first settings were removed from the analysis for all tasks. For each tilt condition, the adjustment was averaged over the five remaining trials. Data analyses were carried out on the angular difference (error) between the dot or arm setting and the reference (true horizon or egocentric position). When ε was negative, the adjustment was below the reference value and when it was positive, the adjustment was above the reference value. To remove any inaccuracy that might be introduced as subjects were being installed on the apparatus (e.g., aligning with great precision the center of the plotter onto the gaze axis), a relative error was calculated. It was the algebraic difference ε between the mean error value under a tilt condition (δ2) and the mean error value measured while subjects were motionless (centrifuge) or upright (tilt chair) (δ1): ε=δ2−δ1.
Results
Horizon and egocentric perception during centrifugation
To investigate whether gravitoinertial modifications affected perception, a three GIF conditions × two references (horizon vs. egocentric) × two tasks (visual vs. kinesthetic) ANOVA with repeated measures on all factors were applied to “ε” for the centrifuge condition. A post hoc analysis (Newman–Keuls) was performed when P<0.05.
As shown in Fig. 2, there was a shift downward to the lower part of the body as indicated by the negative values for all the perceptual tasks. Whatever the perceptual task, the horizon settings were lower than the egocentric ones [mean deviation 10.42 vs. −6.16°, respectively; F(1,14)=15.9; P<0.05]. The visual settings were always lower than the kinesthetic ones [mean deviation 12.35 vs. −4.22°, respectively; F(1,14)=37.1; P<0.05] but no significant interaction of the two factors was found (P>0.05). There was also an increased lowering of the setting errors with GIF [F(2,28)=189.7; P<0.05] whose importance depended on the perceptual task [F(2,28)=42.8; P<0.05] and reference [F(2,28)=13.2; P<0.05]. Increasing GIF lowered more consistently (1) the horizon settings than the egocentric ones (P<0.05) and (2) the visual settings than the kinesthetic ones (P<0.05). In accordance with the literature, the centripetal acceleration induced a displacement of the horizon localization in the direction of GIF suggesting an illusion of tilt in pitching. More surprisingly, increasing the simulated pitch angle also lowered the egocentric perception.

Mean horizon and egocentric settings as a function of the psychophysical task (visual and kinesthetic) when subjects were centrifuged (simulated tilt). Increasing the angle between G and GIF induced a displacement of the horizon and egocentic settings in the direction of GIF
To further evaluate the relationship between the horizon and egocentric perception, a three GIF conditions × two tasks (visual vs. kinesthetic) analysis of covariance (ANCOVA) with repeated measures on the two factors was applied to the horizon settings, with the egocentric estimation as a covariant factor. Results showed that while the effect of GIF remained significant [F(2,16)=5.3; P<0.05], the main effect of task and the interaction of the two factors were no more significant (P>0.05). Thus, the egocentric estimation could account for the horizon setting errors.
Horizon and egocentric perception during true tilt
To investigate whether body tilt influenced perception, a three pitch angles × two references (Horizon vs. egocentric) × two tasks (visual vs. kinesthetic) ANOVA with repeated measures on all factors were applied to “ε” for the true tilt. Results showed a significant effect of task [F(1,14)=28.5; P<0.05] but no effect of reference and pitch angle conditions (P>0.05). An interaction of reference × task and a three-way interaction of reference × task × pitch angle were found [F(1,14)=7.6; F(2,28)=5.4; P<0.05, respectively]. As shown in Fig. 3, the egocentric settings did not vary according to the pitch angle or perceptual modality. Moreover, no systematic bias from the reference was found [reference comparison t test; P>0.05]. A systematic bias under the true horizon for the VPEL settings and above that for the SPH settings was found [mean deviation 4.8 vs. 3.29°, respectively; reference comparison 5.4<t<21.4; P<0.05]. However, while there was no influence of the angle of body tilt on the VPEL, SPH estimates were elevated with backward tilt. Moreover, the egocentric estimation did not account for the difference between the visual and kinesthetic horizon setting errors in true tilt as the ANCOVA showed that the main effect of task [F(1,8)=9.9; P<0.05] and the interaction of task × pitch angle [F(2,16)=14.73; P<0.05] remained significant.

Mean horizon and egocentric settings as a function of the psychophysical task (visual and kinesthetic) when subjects were in the tilting chair (true tilt). The egocentric settings did not vary according to the pitch angle or perceptual modality, whereas a systematic bias under the true horizon for the VPEL settings and above that for the SPH settings was found
Comparison of true and simulated tilt effects on perceptual tasks
Results observed in the centrifuge suggested that the perception is related to the vectorial sum of G and the linear acceleration. The following analyses investigated whether the same orientation relative to the gravitoinertial forces, G for the true tilt or GIF in the centrifuge, induced the same errors in horizon perception. For this purpose, data were expressed in terms of errors with respect to G in the true tilt condition and to GIF in the simulated tilt condition. Negative values indicated settings under the reference.
In order to compare the relationship between perceived horizon and body orientation for true and simulated tilt, a linear regression analysis was applied to the mean individual data recorded in the three body orientations for each of the four experimental conditions (two tilt conditions × two perceptual tasks). The results, summarized in Table 2, showed a significant linear influence of the angle of tilt for the kinesthetic judgments in true and simulated tilt [F(1,43)=4.25; F(1,43)=19.78; P<0.05, respectively]. All kinesthetic estimates seemed to be elevated with backward tilt (positive slope, F(1,84)=19.99, P<0.05). The two straight lines were parallel [deviation from parallelism: F(1,84)=0.99; P>0.05], but the kinesthetic estimates in simulated tilt were always above the ones observed in true tilt [mean error 7.61 vs. 3.29°, respectively; F(1,84)=14.96; P<0.05]. The visual judgments were not influenced by the pitch angles (flat slope, [F(1,84)=3.16, P>0.05]). However, the visual lines were parallel [deviation from parallelism: F(1,84)=0.02; P>0.05] and separated by an offset. Visual estimates in simulated tilt were always above the ones in true tilt [mean error −0.63 vs. −4.81°, respectively; F(1,84)=5.40; P<0.05].
To investigate if the shift between true and simulated tilt was similar for visual and kinesthetic tasks two tasks × three pitch angles ANOVA with repeated measures on the two factors were applied to the difference between errors in simulated tilt and errors in true tilt. Results showed no effect of the task, suggesting that the centrifuge induced a similar offset for the two perceptual tasks.
In order to determine whether response variability could account for the difference between true and simulated tilt, an ANOVA was performed on the mean intrasubjects’ standard deviation. Main effects of tilt [F(1, 14)=7.26; P<0.05] and task [F(1, 14)=20.36; P<0.05] were found. The simulated tilt yielded a smaller intrasubjects’ variability than the true tilt (1.38 vs. 1.61°, respectively). A higher intrasubjects’ variability for the visual judgment tasks was observed whatever the tilt condition (1.79 vs. 1.21° for the visual and kinesthetic tasks, respectively)
Discussion
The aim of the present study was to reassess the tilt/translation ambiguity for common linear accelerations of the usual life—that is, between 1.5 and 3.5 m s−2 of linear acceleration (e.g., an acceleration of 1.52 and 3.43 m s−2 is the acceleration of a car which accelerates from 0 to 100 km h−1 in 18 and 8 s, respectively). Regarding the tilt/translation ambiguity, subjects felt tilted in pitching relative to GIF. However, the same orientation relative to the shear forces G for the true tilt or GIF in the centrifuge did not induce the same horizon perception. Visual adjustment and arm pointing in the centrifuge were always above the ones observed in a G environment. In addition, this offset was similar for both the visual and kinesthetic estimations.
Considering hypotheses on the tilt/translation resolution in the absence of canal/otolith interaction, it can be argued that the utricular/saccular shear force differentiation (Curthoys and Halmagyi 1999; MacDougall et al. 1999) cannot contribute to this difference in perception. For tilt angles under 20°, the difference between the true and simulated tilts was less than 0.55 m s−2 for the saccular shear force stimulation and less than 0.20 m s−2 for the utricular shear force stimulation, that is, close to the otolith vertical and horizontal perceptual thresholds. The lowering of the horizon settings may be related to a misperception of the body z-axis. In other words, a shift of the egocentric coordinates in the direction of GIF may be responsible for errors in the evaluation of the angle between the body axis and the existing forces. In the centrifuge, the mechanical action of GIF on eyes and arm proprioception may be responsible for the lowering of the egocentric coordinates. However, the arm was pulled downward less than the eyes. The body musculature straws under the increased load but this increased load may become conscious when lifting the arm, whereas it was not perceived with a simple eye movement. Moreover, the same horizon setting error was observed whether the response was visual or kinesthetic when the egocentric estimation was taken into account. However, the VPEL and the SPH were lower than the TPE and TPA, respectively. Thus, the lowering of the judgment would not be completely imputable to the mechanical effect of GIF on the effectors. Whether the tilt/translation ambiguity was solved to perform perceptual judgments of an object relative to gravity remains unclear. Subjects seemed to perceive themselves as less tilted in the centrifuge than on the tilt chair. This suggested that the new vertical corresponding to GIF was misperceived.
On the one hand, a sensory adaptation in response to prolonged centrifugation has been reported (Coello et al. 1996; Lackner and DiZio 2003). A different adaptation profile of simulated and true tilt responses can perhaps explain the offset found here. However, analysis of the first five versus the last five judgments showed no difference whatever the perceptual task and tilt conditions. In addition, a kinematics analysis of the kinesthetic judgments showed no difference between the first five and the last five trials whatever the parameter (i.e., peak magnitude, time-to-peak). Moreover, the kinematics profiles were similar for the true and simulated tilts.
On the other hand, this offset may be imputable to a difference in the somesthetic stimulation. The somesthetic system is assumed to provide information about body orientation notably in response to antigravitational forces. It has been shown that labyrinthine defective and normal subjects exhibited a similar estimation of postural tilt whether they were tilted relative to G (Bringoux et al. 2002; Ito and Gresty 1997) or GIF in a centrifuge (Clark and Graybiel 1966) suggesting the contribution of extravestibular graviceptors in body tilt perception. In order to investigate a presumable difference in the alteration of gravity-based tactile stimulation induced by true and simulated tilt, pressure variations were recorded under the bottom—ischiatic tuberosity, in the back, on the scapulas and behind the head. Results from three subjects with different morphological characteristics showed that the pressure values increased both with true and simulated body tilt. More importantly, the similar variations of pressure with respect to body orientation observed for the two conditions suggested that the cutaneous cues indicated a similar body tilt. However, it is simplifying to resume the somesthetic system to cutaneous receptors and internal organs. The cardiovascular system (fluid distribution) is also involved in providing gravity information (Vaitl et al. 1997, 2002). When subjects are seated in the centrifuge, the different somesthetic receptors are at a variable distance from the axis of rotation. As a result, forces acting on vascular pressure receptors at the level of the buttocks and at the level of the feet were quite different. This was not the case in the true tilt situation. The question may be raised if in such a situation of decorrelation subjects could feel less tilted in the centrifuge than in the tilted chair.
Overall, results contradicted the interpretation regarding the tilt/translation ambiguity as the same orientation relative to the shear forces G for the true tilt or GIF in the centrifuge would induce the same horizon perception. Difference in horizon perception could not be attributed to a possible problem of “spatial disorientation” in the centrifuge which interferes with thinking processes in perceptual tasks (Ito and Gresty 1997). The greater precision of the settings in the centrifuge, as reflected by the smaller standard deviation, corresponded to a greater subjects’ confidence regarding the direction of the horizon. However, cognitive information about the environment and its properties may influence spatial orientation (Guedry 1974; Lackner and DiZio 2005; Wertheim et al. 2001). It can be assumed that subjects’ expectations related to centrifugation become important for the self-motion perception system. Whether the tilt/translation ambiguity was solved, none of the subjects reported a translation sensation but rather a tilting sensation. This suggested that even if subjects knew that they were rotated, they did not interpret this movement as a linear horizontal displacement when rotated at a constant velocity. The existence of a tilt response during centrifugation could be due to a misperception of the body z-axis relative to the internal model of G notably in response to the backward direction of somesthetic stimulation which should not exist when the body-axis and gravity are aligned. Whatever the hypothesis for explaining the difference between perception in true and simulated tilt, extravestibular graviceptors may play a crucial role in the judgment of the degree of tilt of one’s own body.
References
Angelaki DE, Dickman JD (2000) Spatiotemporal processing of linear acceleration: primary afferent and central vestibular neuron responses. J Neurophysiol 84:2113–2132
Angelaki DE, McHenry MQ (1999) Short-latency primate vestibuloocular responses during translation. J Neurophysiol 82:1651–1654
Angelaki DE, McHenry MQ, Dickman JD, Newlands SD, Hess BJ (1999) Computation of inertial motion: neural strategies to resolve ambiguous otolith information. J Neurosci 19:316–327
Benson AJ (1990) Sensory functions and limitations of the vestibular system. In: Warren R, Wertheim AH (eds) Perception and control of self-motion. Lawrence Erlbaum, Hillsdale pp 145–170
Bock O (1994) Scaling of joint torque during planar arm movements. Exp Brain Res 101:346–352
Bringoux L, Schmerber S, Nougier V, Dumas G, Barraud PA, Raphel C (2002) Perception of slow pitch body tilts in bilateral labyrinthine-defective subjects. Neuropsychologia 40:367–372
Bringoux L, Tamura K, Faldon M, Gresty MA, Bronstein AM (2004) Influence of whole-body pitch tilt and kinesthetic cues on the perceived gravity-referenced eye level. Exp Brain Res 155:385–392
Brown JH (1966) Interacting vestibular stimuli and nystagmic habituation. Acta Otolaryngol 62:341–350
Carriot J, Bringoux L, Charles C, Mars F, Nougier V, Cian C (2004) Perceived body orientation in microgravity: effects of prior experience and pressure under the feet. Aviat Space Environ Med 75:795–799
Carriot J, Charles C, Huffenus AF, Nougier V, Raphel C, Cian C (2005) Effect of low gravitational stimulation on the perception of target elevation: role of spatial expertise. Percept Psychophys 67:1044–1051
Clark B, Graybiel A (1949a) Apparent rotation of a fixed target associated with linear acceleration in flight. Am J Opthalm 32:549–557
Clark B, Graybiel A (1949b) Linear acceleration and deceleration as factor influencing nonvisual orientation during flight. J Aviat Med 20:92–101
Clark B, Graybiel A (1966) Perception of the visual horizontal in normal and labyrinthine defective observers during prolonged rotation. Am J Psychol 79:608–612
Clement G, Deguine O, Parant M, Costes-Salon MC, Vasseur-Clausen P, Pavy-LeTraon A (2001) Effects of cosmonaut vestibular training on vestibular function prior to spaceflight. Eur J Appl Physiol 85:539–545
Coello Y, Orliaguet JP, Prablanc C (1996) Pointing movement in an artificial perturbing inertial field: a prospective paradigm for motor control study. Neuropsychologia 34:879–892
Cohen MM (1973) Visual feedback, distribution of practice, and intermanual transfer of prism aftereffects. Percept Mot Skills 37:599–609
Cohen MM (1977) Disorienting effects of aircraft catapult launchings: III. Cockpit displays and piloting performance. Aviat Space Environ Med 48:797–804
Cohen M, Stopper A, Welch R, DeRochia C (2001) Effects of gravitational and optical stimulation on the perception of target elevation. Percept Psychophys 63:29–35
Curthoys IS, Halmagyi GM (1999) Vestibular compensation. Adv Otorhinolaryngol 55:82–110
Curthoys IS, Betts GA, Burgess AM, MacDougall HG, Cartwright AD, Halmagyi GM (1999) The planes of the utricular and saccular maculae of the guinea pig. Ann NY Acad Sci 871:27–34
DiZio P, Li W, Lackner JR, Matin L (1997) Combined influences of gravitoinertial force level and visual field pitch on visually perceived eye level. J Vestib Res 7:381–392
Dickman JD, Angelaki DE, Correia MJ (1991) Response properties of gerbil otolith afferents to small angle pitch and roll tilts. Brain Res 556:303–310
Droulez J, Darlot C (1989) The geometric and dynamic implications of the coherence constraints in three-dimensional sensorimotor interactions. In: Jeannerod M (ed) Attention and performance. Erlbaum, New York pp 495–526
Fernandez C, Goldberg JM (1976) Physiology of peripheral neurons innervating otolith organs of the squirrel monkey. III. Response dynamics. J Neurophysiol 39:996–1008
Fernandez C, Goldberg JM, Abend WK (1972) Response to static tilts of peripheral neurons innervating otolith organs of the squirrel monkey. J Neurophysiol 35:978–987
Fitger C (1976) Tactile-kinesthetic space estimation: the influence of gravity. Psychol Res 39:113–135
Gentaz E, Hatwell Y (1996) Role of gravitational cues in the haptic perception of orientation. Percept Psychophys 58:1278–1292
Ghafouri M, Archambault PS, Adamovich SV, Feldman AG (2002) Pointing movements may be produced in different frames of reference depending on the task demand. Brain Res 929:117–128
Gooey K, Bradfield O, Talbot J, Morgan DL, Proske U (2000) Effects of body orientation, load and vibration on sensing position and movement at the human elbow joint. Exp Brain Res 133:340–348
Graybiel A (1952) Oculogravic illusion. AMA Arch Ophthalmol 48:605–615
Guedry FE (1974) Psychophysics of vestibular sensation. Handbook of sensory physiology. Kornhuber HH, New York, pp 1–154
Guedry FE, Mortensen CE, Nelson JB, Correia MJ (1978) A comparison of nystagmus and turning sensations generated by active and passive turning. In: Hood JD (ed) Vestibular mechanisms in health and disease. Plenum, New York, pp 317–325
Holly JE, McCollum G (1996) The shape of self-motion perception-I. Equivalence classification for sustained motions. Neuroscience 70:461–486
Howard IP, Templeton WB (1966) Human spatial orientation. Wiley, London
Ito Y, Gresty MA (1997) Subjective postural orientation and visual vertical during slow pitch tilt for the seated human subject. Aviat Space Environ Med 68:3–12
Lackner JR, DiZio PA (2003) Adaptation to rotating artificial gravity environments. J Vestib Res 13:321–330
Lackner JR, DiZio P (2005) Vestibular, proprioceptive, and haptic contributions to spatial orientation. Annu Rev Psychol 56:115–147
Li W, Matin L (1993) Eye & head position, visual pitch, and perceived eye level. Invest Ophthalmol Vis Sci 34:1311
Li W, Matin L (1995) Differences in influence between pitched-from-vertical lines and slanted-from-frontal horizontal lines on egocentric localization. Percept Psychophys 57:71–83
Loe PR, Tomko DL, Werner G (1973) The neural signal of angular head position in primary afferent vestibular nerve axons. J Physiol 230:29–50
Long GM, Ambler RK, Guedry FE Jr (1975) Relationship between perceptual style and reactivity to motion. J Appl Psychol 60:599–605
Luyat M, Gentaz E, Corte TR, Guerraz M (2001) Reference frames and haptic perception of orientation: body and head tilt effects on the oblique effect. Percept Psychophys 63:541–554
MacDougall HG, Curthoys IS, Betts GA, Burgess AM, Halmagyi GM (1999) Human ocular counterrolling during roll-tilt and centrifugation. Ann N Y Acad Sci 871:173–180
Matin L, Li W (1992) Mislocalizations of visual elevation and visual vertical induced by visual pitch: the great circle model. Ann N Y Acad Sci 656:242–265
Matin L, Li W (1994) The influence of a stationary single line in darkness on the visual perception of eye level. Vision Res 34:311–330
Matin L, Li W (1995) Multimodal basis for egocentric spatial localization and orientation. J Vestib Res 5:499–518
Mayne R (1974) A systems concept of vestibular organs. In: Handbook of sensory physiology. Springer, Berlin Heidelberg New York, pp 493–580
McIntyre J, Stratta F, Droulez J, Lacquaniti F (2000) Analysis of pointing errors reveals properties of data representations and coordinate transformations within the central nervous system. Neural Comput 12:2823–2855
Merfeld DM, Zupan LH (2002) Neural processing of gravitoinertial cues in humans. III. Modeling tilt and translation responses. J Neurophysiol 87:819–833
Merfeld DM, Zupan LH, Gifford CA (2001) Neural processing of gravito-inertial cues in humans. II. Influence of the semicircular canals during eccentric rotation. J Neurophysiol 85:1648–1660
Merfeld DM, Park S, Gianna-Poulin C, Black FO, Wood S (2005a) Vestibular perception and action employ qualitatively different mechanisms: II. VOR and perceptual responses during combined tilt & translation. J Neurophysiol 94(1):199–205
Merfeld DM, Park S, Gianna-Poulin C, Black FO, Wood S (2005b) Vestibular perception and action employ qualitatively different mechanisms: I. Frequency response of VOR and perceptual responses during translation and tilt. J Neurophysiol 94(1):186-98
Paige GD, Tomko DL (1991) Eye movement responses to linear head motion in the squirrel monkey. II. Visual-vestibular interactions and kinematic considerations. J Neurophysiol 65:1183–1196
Prieur JM, Bourdin C, Vercher JL, Sares F, Blouin J, Gauthier GM (2005) Accuracy of spatial localization depending on head posture in a perturbed gravitoinertial force field. Exp Brain Res 161:432–440
Raphel C, Barraud PA (1994) Perceptual thresholds of radial accelerations as indicated by visually perceived eye level. Aviat Space Environ Med 65:204–208
Raphel C, Barraud PA, Koessler C, Cian C (1996) Effects of a visual frame and of low radial accelerations on the visually perceived eye level. Percept Psychophys 58:906–914
Schöne H (1964) On the role of gravity in human spatial orientation. Aerosp Med 35:764–772
Seidman SH, Telford L, Paige GD (1998) Tilt perception during dynamic linear acceleration. Exp Brain Res 119:307–314
Si X, Angelaki DE, Dickman JD (1997) Response properties of pigeon otolith afferents to linear acceleration. Exp Brain Res 117:242–250
Tremblay L, Elliott D (2003) Contribution of action to perception of self-orientation in humans. Neurosci Lett 349:99–102
Vailt D, Mittelstaedt H, Baisch F (1997) Shifts in blood volume alter the perception of posture. Int J Psychophysiol 2:99–105
Vailt D, Mittelstaedt H, Saborowski R, Stark R, Baisch F (2002) Shifts in blood volume alter the perception of posture: further evidence for somatic graviception. Int J Psychophysiol 44:1–11
Wertheim A, Mesland B, Bles W (2001) Cognitive suppression of tilt sensations during linear horizontal self-motion in the dark. Perception 30:733–741
Whiteside TC, Graybiel A, Niven JI (1965) Visual illusions of movement. Brain 88:193–210
Worringham CJ, Stelmach GE (1985) The contribution of gravitational torques to limb position sense. Exp Brain Res 65:38–42
Zupan LH, Merfeld DM, Darlot C (2002) Using sensory weighting to model the influence of canal, otolith and visual cues on spatial orientation and eye movements. Biol Cybern 86:209–230
Acknowledgement
The authors are grateful to Mr Alain ROUX of the Centre de Recherches du Service de Santé des Armées for the technical expertise and helpful assistance.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Carriot, J., Barraud, PA., Nougier, V. et al. Difference in the perception of the horizon during true and simulated tilt in the absence of semicircular canal cues. Exp Brain Res 174, 158–166 (2006). https://doi.org/10.1007/s00221-006-0434-6
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00221-006-0434-6