
Abstract
It has been proposed that visually guided reaching movements performed in the lower visual field (LVF) of peripersonal space are more effective and efficient than their upper visual field (UVF) counterparts (Danckert and Goodale 2001). In the present investigation we sought to determine whether this purported visual field asymmetry reflects advantaged processing of online visual feedback. To accomplish that objective, participants performed discrete reaching movements to each of three target locations in the LVF and UVF. In addition, reaches were completed under conditions wherein target location remained constant throughout a reaching response (i.e., control trials) and a separate condition wherein target location unexpectedly perturbed at movement onset (i.e., experimental trials). We reasoned that the target perturbation paradigm would provide a novel means to assess a possible superior–inferior visual field asymmetry for online reaching control. In terms of the impact of a target perturbation, both visual fields demonstrated equal proficiency integrating visual feedback for online limb adjustments. Interestingly, however, the spatial distribution of movement endpoints in the LVF was less than UVF counterparts (cf. Binsted and Heath 2005). Taken together, the present findings suggest that although LVF and UVF reaches readily use visual feedback to accommodate an unexpected target perturbation, reaches in the LVF elicit advantaged spatial benefits influencing the effectiveness of online limb corrections.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Online adjustments to visually guided reaching movements are thought to be mediated by dedicated visuomotor mechanisms that reside in the posterior parietal cortex (PPC) of the dorsal visual pathway. In support of that position, single neuron recording in the monkey has shown preferential reach-related activity in area 7a for online error corrections (MacKay 1992; see also Mountcastle et al. 1975). In humans, selective transcranial magnetic stimulation of the PPC (Desmurget et al. 1999) as well as naturally occurring PPC lesions (Pisella et al. 2000) disrupt the normally “automatic” limb adjustments characterizing unexpected changes in target position (e.g., Goodale et al. 1986; see also Gréa et al. 2002).
Interestingly, it has been proposed that an anatomical disparity in the superior – inferior retinal axis (Curcio and Allen 1990; Dichtl et al. 1999 Footnote 1) leads to a lower visual field (LVF) advantage for attentional resolution (Handy et al. 2003), motion segmentation (Lakha and Humphreys 2005) and the control of goal-directed actions (see Previc 1990 or Previc 1998 for review). With specific regard to goal-directed reaching movements, an over-representation of the LVF within a number of the constituent regions of the dorsal visual pathway (Maunsell and Van Essen 1987; Galati et al. 2000) is thought to underpin a functional bias for the processing of visual cues for online control in lower working space (see Danckert and Goodale 2003 for a review). In support of that position, Danckert and Goodale (2001) found that continuous reaches (i.e., a Fitts reciprocal aiming task) to targets of various sizes were generally more accurate when performed in the LVF as opposed to the upper visual field (UVF).Footnote 2 Moreover, LVF, but not UVF, reaches produced a robust speed-accuracy trade-off as defined by Fitts Law (Fitts 1954). Similarly, Khan and Lawrence (2005) reported that LVF reaches elicit reduced spatial variations late in the reaching trajectory and enhanced endpoint accuracy: a finding they attributed to improved feedback processing in the lower region of working space. It is, however, important to note that Binsted and Heath (2005) did not observe a functional asymmetry for reaches performed in the superior – inferior retinal axis across a number of movement parameters. For example, Binsted and Heath (2005) reported the magnitude and timing of initial movement impulses and overall durations of LVF and UVF reaches elicited similar speed versus accuracy relations and comparable endpoint accuracy values. That study did, however, report a small but statistically reliable advantage for LVF reaches in terms of the spatial variability of movement endpoints (cf. Khan and Lawrence 2005). Thus, examination of the three published studies contrasting visual field asymmetries in goal-directed aiming highlights a degree of controversy regarding the extent to which a LVF advantage for visuomotor control is systematic and generalizable.
In the present investigation we sought to determine if the discrepant findings just described might relate to between-experiment differences in the extent reaching movements were controlled primarily online or primarily offline (see Heath 2005). Indeed, the instruction set provided to participants in the Binsted and Heath (2005) investigation emphasized the optimization of movement speed and produced average movement times of 360 ms. That instruction set has been shown to minimize participants’ use of visual feedback for online error reduction (e.g., Vince 1948; Keele 1968). In contrast, Danckert and Goodale’s (2001) instruction set emphasizing “both speed and accuracy equally” (p. 304) as well as Khan and Lawrence’s (2005) instruction set to move “as smoothly as possible” (p. 396) in a criterion time of 400 ms may have engendered participants adoption of a more online mode of reaching control. Notably, a LVF advantage for reaching control might be restricted to situations wherein visual information is used for online error nullification.
The present study employed a variant of the discrete reaching task used by Khan and Lawrence (2005) and Binsted and Heath (2005). In particular, our investigation included a condition wherein the position of a to-be-touched target object was unexpectedly perturbed at the onset of a reaching movement, thus creating a requirement for online and visually based limb adjustments.Footnote 3 Previous studies employing the target perturbation paradigm used here have shown that the visuomotor system is readily able to modify an initially inappropriate movement trajectory via the evocation of discrete (Carnahan et al. 1993; Heath et al. 1998, 1999; Paulignan et al. 1997) and/or continuous corrections to the movement trajectory (Elliott et al. 1999a; Heath 2005; Pelisson et al. 1986). Importantly, the present investigation required participants to foveate on a position above or below target space prior to and during reaching movements so that responses were completed in LVF and UVF, respectively. We reasoned that such a manipulation provided a novel means to test whether preferred connections of the LVF to the dorsal visual pathway elicit more efficient and effective online movement corrections than reaches performed in the UVF of peripersonal space.
Methods
Participants
Twenty-five participants from the Indiana University community volunteered for this experiment (18–34 years: 14 males and 11 females). All participants reported normal vision and were right-handed as determined by a modified version of the University of Waterloo Handedness questionnaire (Bryden 1977). Participants provided informed consent approved by the Office of Human Research, Indiana University, and this study was conducted in accordance with the ethical standards laid down in the 1964 Declaration of Helsinki.
Apparatus
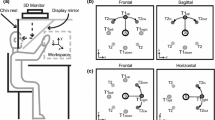
An aiming apparatus similar to Held and Gottlieb (1958) was used in this experiment. The apparatus was placed on a normal table-top and consisted of a two-sided rectangular box (74 cm high, 96 cm wide, 60 cm deep) divided in half by a partially transparent mirror. A 17-in. computer monitor (NEC Multisync 1765: 16 ms response rate) was placed upside down on the superior surface of the apparatus in order to project stimuli onto the partially transparent mirror. The difference in height between the computer monitor and mirror, and the mirror and the lower surface of the aiming apparatus was constant (i.e., 37 cm), thus an image projected onto the mirror appeared on the surface of the aiming apparatus (i.e., below the mirror). Participants sat at an open end of the apparatus and completed reaching movements to virtual targets (see below) that appeared on the surface of the apparatus. The distance between the eyes and virtual targets was ~42 cm and this optical geometry was maintained via a head–chin rest (Lafayette Instruments, Model 14302).
Reaching movements were initiated from a common home position (i.e., a microswitch located 16 cm to the left of the participants’ midline and 31 cm from the front edge of the table surface) to “touch” a virtual target 35 cm (near), 38 cm (middle), and 41 cm (far) to the right of the home position (and 31 cm from the front edge of the table). Targets were white circles (0.5 cm in diameter) presented against a high-contrast black background. Additionally, a fixation cross (2×2 cm) was projected 8 cm above or below target location. The visual angle between the fixation cross and target was 12°. Fixating on the cross above the target resulted in reaching movements conducted in the LVF, whereas fixating on the cross below the target resulted in reaching movements conducted in UVF.
The lights in the experimental suite were darkened to prevent direct viewing of the aiming limb. In place of the veridical limb, two light emitting diodes (red LEDs) attached to a splint complex were secured to the index finger of the right hand (i.e., the pointing finger) to provide visual feedback about limb position. Eprime (ver 1.0) was used to present visual stimuli and to control visual and auditory events.
Procedure
Two trial blocks (control and experimental) were used in this investigation. The presentation of trial blocks was counterbalanced across participants. Participants were instructed to point to a target using the index finger of their right hand emphasizing “the accuracy and speed” of their movement. As stated in the Introduction, we emphasized both the accuracy and speed of reaching movements to ensure participants reaching strategy would allow for the integration of visual feedback for online error corrections. A trial began once participants depressed the home position. That action illuminated the LEDs attached to the splint complex, and the LEDs remained illuminated for the duration of a trial. Additionally, depressing the home position resulted in projection of the fixation cross and participants were required to foveate this position for the duration of a trial. After a 1,000 ms interval, the fixation cross was concurrently presented with a target object for a 1,500 ms preview period.
During control trials, one of the three target locations (e.g., near, middle, far) was presented during the preview phase, after which participants received an auditory tone instructing them to initiate their reaching movement. The target and the fixation cross remained visible throughout the reaching movement. Participants completed an equal number of reaches to each target location in each visual field for a total of 90 control trials. Reaches in the UVF and LVF, as well as the order of target location, were ordered pseudo-randomly. Importantly, participants were made aware that the location of the target would not change during control trials.
In the experimental trials, the same procedures as just described were employed with two exceptions. First, the middle target was always presented during preview. Secondly, on a small percentage of trials an unpredictable target perturbation occurred at movement onset. Specifically, during 70% of the experimental trials the middle target also served as the target location during the reaching response, whereas the remaining 30% of experimental trials entailed a target perturbation once participant’s finger released pressure from the home position. For one half of the perturbation trials, the middle target was replaced with the near target, and on the other half, the middle target was replaced with the far target. Participants completed a total of 200 experimental trials; hence, the number of perturbation trials associated with the near and the far target corresponds to the number of near and far trials performed in the block of control trials. The ordering of target perturbations as well as the ordering of the visual fixation was pseudo-randomized. Participants were told a priori that a small percentage of experimental trials would entail an unexpected change in target location.
Data collection and reduction
In addition to containing dual LEDs, the splint complex attached to the pointing finger contained an infra-red emitting diode (IRED). IRED position data were sampled at 200 Hz for 2 s following the auditory initiation cue via an OPTOTRAK 3020 (NDI, Waterloo, ON, Canada). Off-line, displacement data were filtered via a second-order dual-pass Butterworth filter employing a low-pass cut-off frequency of 15 Hz. Instantaneous velocities were calculated by differentiating the displacement data using a three-point central finite difference algorithm. Velocity data were differentiated again to obtain acceleration information.
To make certain visual fixation was maintained during reaching movements, a SONY TR42 8-mm camcorder filming in high-speed mode (60 Hz) was used to provide up-close images of participants’ eyes. The video image was monitored online via a 13-in. Toshiba color CRT. Trials in which the experimenter detected an eye movement during the response accounted for no more than 1% of the trials for any participant and were not included in subsequent data analyses.
Dependent variables and statistical analyses
Movement onset was the frame at which the instantaneous velocity rose above 50 mm/s for ten consecutive frames (50 ms). Movement offset was the frame at which instantaneous velocity fell below a value of 50 mm/s for ten connective frames. Dependent variables included: movement time (MT: time between movement onset and offset), peak velocity (PV: maximum resultant velocity between movement onset and offset), time to peak velocity (TPV: time between movement onset and maximum resultant velocity), time after peak velocity (TAPV: time between maximum resultant velocity and movement offset), constant error (CE) in the primary movement direction (overshoot = +CE, undershoot = -CE) and its associated variable error (VE) value. We also computed the spatial position of the limb in the primary movement direction at discrete points in the reaching trajectory to determine the extent to which our experimental manipulations influenced unfolding reaching responses. Specifically, we measured limb displacement at four kinematic markers: peak acceleration (PA: maximum resultant acceleration between movement onset and offset), PV, peak deceleration (PD: maximum resultant negative acceleration between movement onset and offset) and the ultimate endpoint of the response (END: i.e., position of limb at movement offset). Additionally, we computed the proportion of endpoint variability (R 2) explained by limb displacement at PV and PD separately for each participant.Footnote 4 The logic behind this analysis is that reaches controlled more online should produce lower R 2 values as the unfolding trajectory is modified to attenuate early planning errors (see Heath 2005 for complete details).
Corrective submovements (i.e., discrete movement corrections) were identified using the algorithms developed by Chua and Elliott (1993). Corrective submovements were defined as significant deviations in acceleration over and above the primary acceleration and deceleration, and included deviations before and after PV, and secondary zero-crossings. A significant deviation was defined as a “fluctuation” that did not lead to a change in sign lasting at least 70 ms (i.e., 14 samples) and meeting an amplitude criterion of 10% of the greatest absolute acceleration value. Zero-crossing were negative to positive transitions in acceleration. As well, reversals in the direction of movement were determined from the velocity profiles. Reversals were positive to negative transitions in velocity reflecting a change from forward to backward movement. These corrections were tallied and divided by the number of trials in each condition to determine the average number of corrections per trial.
An alpha level of 0.05 was used to interpret all omnibus tests and only significant effects are reported. Unless otherwise stated (see exceptions below), dependent variables were subjected to 2 (block: control, experimental) by 2 (visual field: UVF, LVF) by 3 (target: near, middle, far) repeated-measures ANOVA. Where appropriate, F-statistics were corrected for violations of sphericity using the appropriate Huynh-Feldt correction (corrected degrees of freedom reported to one decimal place). Significant effects/interactions were decomposed using simple effects or power polynomials. For comparison with other studies examining visual field asymmetries in goal-directed aiming, Table 1 provides ANOVA summary of the impact of visual field across common variables used in this and other research.
Results
Performance measures
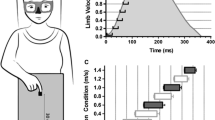
The analysis of MT exhibited an effect for target, F(2, 48)=23.86, P<0.001, and a block by target interaction, F(2, 48)=4.98, P<0.02. Reaches to the near target during experimental trials elicited longer MTs than their control condition counterparts (t(24)=2.25, P<0.04). MTs for reaches to the middle and far target did not vary as a function of experimental condition (ts(24)=1.03 and 0.77, respectively Ps>0.05) (Fig. 1). We also computed slopes of the relationship between target distance and MT separately for each participant and subjected those data to 2 (block: experimental, control) by 2 (visual field: LVF, UVF) repeated-measures ANOVA. Analysis of slope values produced an effect for block, F(1, 24)=7.49, P<0.02: slopes for experimental trials (4 ms) were shallower than control trials (10 ms). Slopes for the LVF (6 ms) and UVF (7 ms) did not differ.

The top panel depicts peak velocity (PV: mm/s), and the lower panel depicts movement time (MT: ms) as function of block (experimental, control), visual field [lower visual field (LVF), upper visual field (UVF)] and target location (near, middle, far)
Our analysis of CE showed that control trials undershot target location less than experimental trials, F(1, 24)=4.29, P<0.05. The results for VE showed that LVF reaches were less variable than UVF reaches, F(1, 24)=4.51, P<0.05. In addition, VE elicited an effect for block, F(1, 24)=7.20, P<0.02, and an interaction involving block by target, F(2, 48)=6.29, P<0.01. Reaches to the near and far target were more variable during experimental than control trials (ts(24)=2.60 and 3.11, respectively, Ps<0.02). Reaches to the middle target did not vary across experimental conditions (t(24)=1.03, P>0.05) (see Table 2 for CE and VE experimental means).
Kinematic measures
The results for PV yielded an effect for target, F(2, 48)=283.17, P<0.001, and a block by target interaction, F(2, 48)=53.01, P<0.001. PVs for reaches to the near target during experimental trials were larger than their control condition counterparts (t(24)=3.06, P<0.01), whereas PVs did not differ for the middle target (t(24)=−0.44, P>0.05). For the far target, PVs during experimental trials were less than control trials (t(24)=−3.26, P<0.01) (Fig. 1). In line with our MT analysis, we computed slopes for the relation between PV and target distance. It was found that slopes for control trials (129 mm/s) were steeper than experimental trials (38 mm/s), F(1, 24)=26.50, P<0.001. The positive slope for experimental trials indicates a degree of scaling with target eccentricity, albeit to a lesser degree than control trials. Slopes for the LVF (83 mm/s) and UVF (84 mm/s) did not vary.
The results for TPV revealed an effect for target, F(2, 48)=40.93, P<0.001, such that TPV increased with target eccentricity (only linear effect significant: F(1, 24)=49.53, P<0.001). The analysis of TAPV yielded a block by target interaction, F(2, 48)=6.69, P<0.01. Reaches to the near target during experimental trials spent more time after PV than reaches to the same target during control trials (t(24)=3.00, P<0.01). TAPV for reaches to middle and far targets did not vary across control and experimental trials (ts(24)=0.60 and −0.54, respectively, Ps>0.05).
Spatial displacement of reaching trajectories
Limb displacement in the primary movement direction was examined via 4 (marker: PA, PV, PD, END) by 2 (block: control, experimental) by 2 (visual field: UVF, LVF) by 3 (target: near, middle, far) repeated-measures ANOVA. This analysis produced effects for marker, F(1.9, 45.7)=857.15, P<0.001, block, F(1, 24)=14.88, P<0.02, and target, F(2, 48)=954.84, P<0.001, as well as interactions involving marker by target, F(4.6, 111.6)=187.25, P<0.001, and marker by block by target, F(2.7, 66.8)=9.82, P<0.001. We elected to decompose the three-way interaction by examining the effect for target separately at each kinematic maker and block. At PA, target eccentricity did not influence experimental or control trials (Fs(2,48)=1.95 and 2.07, respectively, Ps>0.05). At PV (F(2, 48)=7.41 and 84.24, Ps<0.01), PD (Fs(2, 48)=184.60 and 677.51, Ps<0.001) and END (Fs(2, 48)=476.67 and 1774.57, Ps<0.001), however, limb displacement scaled to target eccentricity for both experimental and control trials, respectively. Although these post hoc contrasts did not uncover the differential impact of block on marker and target, examination of Table 3 indicates the influence of block stemmed from a more robust scaling between limb displacement and target location for control trials—particularly at PV.
Proportion of endpoint variance (R 2) explained at PV and PD and corrective submovements
R2 values were submitted to 2 (marker: PV, PD) by 2 (block: control, experimental) by 2 (visual field: UVF, LVF) by 3 (target: near, middle, far) repeated-measures ANOVA. This analysis produced effects for marker, F(1, 24)=33.28, P<0.001, and target, F(2, 48)=4.92, P<0.02. R2 values increased from PV (0.13±0.01) to PD (0.37±0.04) and R2 values for near (0.25±0.02) and far (0.28±0.03) targets were less than the middle target (0.21±0.02) (significant quadratic polynomial: F(1, 24)=19.30, P<0.001).
Analysis of corrective submovements revealed effects for block, F(1, 24)=11.01, P<0.01, target, F(2, 48)=5.27, P<0.01, and an interaction involving block by target, F(2, 48)=21.94, P<0.001. Reaches to the near (0.82) and far (0.65) targets during experimental trials elicited more corrective submovements than their control condition counterparts (near=0.53; far=0.53) (ts(24)=4.45 and 3.89, respectively, Ps<0.01). The mean number of corrective submovements for the middle target did not vary across experimental (0.63) and control (0.59) trials (t(24)=1.53, P>0.05).
Discussion
The current study examined whether online movement corrections are more effective and efficient when implemented in the LVF of peripersonal space. To accomplish that objective, we examined the trajectories of discrete reaches performed in the LVF and UVF under conditions wherein target location remained constant (i.e., control trials) or was unexpectedly perturbed at movement onset (i.e., experimental trials). We reasoned that the latter situation would create a requirement to modify an originally planned movement trajectory, thus providing a novel opportunity to examine the hypothesized LVF advantage for online limb adjustments.
The impact of an unexpected target perturbation: evidence for online control
The magnitude and timing of peak velocity scaled in relation to final target properties during both control and experimental trials. Interestingly, however, the slopes relating PV to target eccentricity were much shallower for experimental as compared to control trials (cf. Heath et al. 1998). Taken together, those findings indicate that although some trajectory reorganization occurred during the early stages of a perturbation trial (e.g., Bédard and Proteau 2004), initial reaching kinematics were influenced by target characteristics available to the visuomotor system at the time of response cuing (the so-called real-time control hypothesis: Westwood and Goodale 2003). Such a finding is in keeping with a two-component model of reaching control (i.e., Woodworth 1899) and the notion that central planning mechanisms governing the initial stages of visually guided actions are largely, but not entirely, refractory to the integration of afferent resources for online error reduction (e.g., Beggs and Howarth 1972; Carlton 1981).
In spite of the fact that the initial kinematics of some experimental trials (i.e., the perturbation trials) were inappropriate for the optimization of speed and accuracy, participants’ trajectories were reorganized based on final target properties (cf. Heath et al. 1998, 1999). Indeed, Table 3 shows that in response to a target perturbation participants amended the spatial characteristics of their reaching trajectory en route to the target. In terms of the overall duration of reaching movements, MTs for the middle and far target did not vary as a function of control and experimental trials. Thus, participants efficiently modified their reaching trajectory when a target perturbation occurred in a direction consistent with the primary movement impulse. For a perturbation to the near target, however, MTs were significantly longer than their control condition counterparts. That pattern of results has been previously linked to the increased mechanical and psychophysical difficulty associated with applying corrective deceleration forces in a direction opposite the primary movement impulse (Elliott et al. 1999b, 2001; Heath and Westwood 2003; Oliveira et al. 2005). In line with that position, our results show the overall lengthening of MT for a near target perturbation to be attributed to increased TAPV, that is, the time when the limb is decelerating and the operator is devoting significant resources to the evocation of online adjustments required to “hit the target” (e.g., Langolf et al. 1976; MacKenzie et al. 1987; Krigolson and Heath 2004).
Concerning the nature of movement corrections, trials containing a target perturbation elicited more corrective submovements than trials wherein target position remained stationary (cf. Heath et al. 1998, 1999). Moreover, the majority of corrective submovements (i.e., 75%) occurred after peak velocity. These results in combination with the findings for TAPV described just above appear to link visually based movement corrections to discrete adjustments during the deceleration phase of the reaching trajectory (e.g., Beggs and Howarth 1972; Meyer et al. 1988). That explanation, however, is tempered by the fact that the timing, magnitude and displacement of the limb at peak velocity during perturbation trials scaled in relation to final target characteristics (albeit peak velocity scaled to a lesser degree for experimental than control trials). Moreover, experimental and control trials elicited similar R 2 values at PV and PD. Thus, participants’ ability to correct their reaching trajectory online does not appear to have been entirely limited to discrete adjustments during the latter stage of the reaching trajectory; rather, evidence suggests that a combination of discrete and continuous (e.g., Georgopoulos et al. 1983; Heath et al. 2004; Pelisson et al. 1986; Stubbs 1976) corrections enabled the visuomotor system to effect temporally efficient and effective online movement corrections throughout the movement trajectory (see Elliott et al. 1999a for review).Footnote 5 Importantly, the fact that experimental and control trials exhibited broadly comparable movements durations and endpoint accuracy values indicates that visual feedback was used to accommodate an unexpected target perturbation (cf. Heath et al. 1998).
Online movement control in the LVF and UVF
As just described, the target perturbation paradigm used here mandated online limb adjustments and thus provided a means to explore the hypothesized LVF advantage for online reaching control (e.g., Khan and Lawrence 2005). In terms of early reaching kinematics, neither experimental nor control trials elicited a visual field asymmetry. As such, the timing and magnitude of PV, as well as the slopes associated with the linear equations for PV, did not differ for LVF and UVF reaches. Moreover, the spatial position of the limb during early reaching kinematics (i.e., PA and PV) did not differentiate between LVF and UVF reaches. Those results suggest that the LVF does not access preferential visual inputs for the initial kinematic parameterization of action. As indicated above, however, it is possible that the expression of a LVF advantage might be limited to situations wherein visual feedback is used later in the reaching trajectory for movement corrections. Thus, a LVF advantage might be preferentially asserted during the later stages of reaching, particularly when the performer modifies their trajectory to accommodate a target perturbation. In examination of that hypothesis, we found that MT and the slopes associated with the linear equations for MT, as well as endpoint accuracy values (i.e., CE) did not elicit a visual field asymmetry. Moreover, participants devoted similar epochs in time (i.e., TAPV), a similar number of corrective submovements and demonstrated equivalent R 2 values for LVF and UVF reaches—evidencing a comparable level of online control. Interestingly, however, we did observe a statistically reliable advantage for LVF reaches in terms of the spatial distribution of movement endpoints (i.e., VE). We will return to this important metric following redress of the temporal and endpoint accuracy symmetry of LVF and UVF reaching movements.
Recall Danckert and Goodale’s (2001) observation that reaches in the LVF, but not UVF, produce robust speed versus accuracy relations. In contrast, the control trials used in the present study as well as Binsted and Heath’s (2005) results demonstrate that early and late reaching kinematics elicit robust speed versus accuracy relations for both visual fields. In terms of reconciling this discrepancy, it is entirely possible that Danckert and Goodale’s use of exceptionally low index of difficulties (IDs: 0.3–1.5 bits) contributed to the production of essentially ballistic actions (see Gan and Hoffman 1988) selectively restricting the expression of speed versus accuracy relations to the LVF. The range of IDs used presently and in Binsted and Heath (IDs: 1.5–7 bits), however, was selected on the basis of values that were originally demonstrated by Fitts (1954) to show a robust log-linear relation between movement difficulty and movement time. Hence, we assert that speed versus accuracy relations can be expected for both LVF and UVF reaches when movements are completed within a range of IDs specified in Fitts’ classic research.
Concerning the issue of response accuracy, the three published studies examining the endpoint accuracy of LVF and UVF reaching movements have produced mixed results. One study reported null differences (Binsted and Heath 2005) whereas the other two studies (Danckert and Goodale 2001; Khan and Lawrence 2005) noted improved accuracy for the LVF: a finding Khan and Lawrence linked to superior feedback-based processing in the LVF. Of course, in the present investigation we employed a target perturbation paradigm as a direct test of the reported visual field asymmetry for visual feedback processing and did not find evidence of a LVF advantage for endpoint accuracy. Moreover, results for movement duration and movement characteristics during the deceleration phase of reaching trajectories did not show evidence of a visual field asymmetry.
As one might expect, it is entirely possible that the discrepant findings just mentioned might be best explained by means of between-experiment methodological differences. For example, in the Introduction we forwarded the notion that an instruction set emphasizing both movement accuracy and speed (Danckert and Goodale 2001) or the “smoothness” of a reaching response (Khan and Lawrence 2005) relative to instructions emphasizing movement speed alone (Binsted and Heath 2005) might impact reaching control and account for the equivocal expression of a visual field asymmetry. We, however, are able to rule-out that possibility because the instruction set used here (i.e., emphasizing both movement speed and accuracy) is parallel to that used in a previous study reporting a reliable LVF advantage for endpoint accuracy (i.e., Danckert and Goodale 2001). Thus, the present findings add importantly to the literature because they highlight the existence of a controversy surrounding the extent a LVF advantage can be attributed to a broad range of parameters underlying visually guided actions.
Notwithstanding the symmetrical temporal and accuracy characteristics of our LVF and UVF reaching movements, we did observe a LVF advantage in terms of endpoint variability. Similar findings were reported by Khan and Lawrence (2005) and Binsted and Heath (2005).Footnote 6 Moreover, Brown et al. (2005) showed that maximum grip aperture is more stable when reaching to grasp an object in the LVF. Thus, converging behavioural evidence suggests that the greater density of retinal ganglion cells in the superior hemiretina (Curcio and Allen 1990) persisting to the visuomotor networks of the dorsal visual pathway (e.g., Previc 1990) may enhance the fidelity of visuomotor processing and improve the stability of actions performed in the LVF of peripersonal space.
Notes
Interestingly, Dichtl et al. (1999) reported that retinal nerve fiber thickness is greater at the inferior disc border than the superior disc border.
Danckert and Goodale (2001) report a LVF endpoint accuracy advantage for target “lengths” of 3.7, 7.5, 14.9 and 30.0 mm. A target “length” of 1.9 mm, however, did not produce a visual field asymmetry.
Research employing the double-step paradigm has shown that online movement corrections occur involuntarily (Goodale et al. 1986) via an ‘automatic pilot’ (Pisella et al. 2000) operating without awareness from the participant. It is important to note that the target perturbation used in the present investigation was explicit (e.g., Heath et al. 1998) and participants were aware of the need for online limb adjustments.
Previous research has shown that the proportion of endpoint variance explained by the spatial position of the limb at peak acceleration does not provide sufficient predictive power to explain unfolding reaching kinematics (see Heath 2005; Heath et al. 2004). Hence, in the present research R2 values for movement endpoints were restricted to the spatial location of the limb at peak velocity and peak deceleration.
Endpoint variability for trials involving a perturbation was greater than for trials involving a stationary target position. As stated elsewhere, increased endpoint variability is thought to represent inherent neuromuscular instability in the evocation of online limb adjustments (e.g., Khan et al. 2002).
References
Bédard P, Proteau L (2004) On-line vs. off-line utilization of peripheral visual afferent information to ensure spatial accuracy of goal-directed movements. Exp Brain Res 158:75–85
Beggs WD, Howarth CI (1972) The accuracy of aiming at a target: some further evidence for a theory of intermittent control. Acta Psychol 30:171–177
Binsted G, Heath M (2005) No evidence of a lower visual field specialization for visuomotor control. Exp Brain Res 162:89–94
Brown LE, Halpert BA, Goodale MA (2005) Peripheral vision for perception and action. Exp Brain Res 165:97–106
Bryden MP (1977) Measuring handedness with questionnaires. Neuropsychologia 15:617–624
Carlton LG (1981) Visual information: The control of aiming movements. Q J Exp Psychol 33A:87–93
Carnahan H, Goodale MA, Marteniuk RG (1993) Reaching versus grasping and the differential use of visual feedback. Hum Mov Sci 12:219–234
Chua R, Elliott D (1993) Visual regulation of manual aiming. Hum Mov Sci 12:365–401
Curcio CA, Allen KA (1990) Topography of ganglion cells in human retina. J Comp Neurol 300:5–25
Danckert J, Goodale MA (2001) Superior performance for visually guided pointing in the lower visual field. Exp Brain Res 137:303–308
Danckert J, Goodale MA (2003) The ups and downs of visual perception. In: Johnson SH (eds) Taking action: cognitive neuroscience perspectives on intentional acts. MIT, Cambridge, pp 29–64
Desmurget M, Epstein CM, Turner RS, Prablanc C, Alexander GE, Grafton ST (1999) Role of the posterior parietal cortex in updating reaching movements to a visual target. Nat Neurosci 2:563–567
Dichtl A, Jonas JB, Naumann GOH (1999) Retinal nerve fiber layer thickness in human eyes. Graefe’s Arch Clin Exp Opthalmol 237:474–479
Elliott D, Binsted G, Heath M (1999a) The control of goal-directed limb movements: correcting errors in the trajectory. Hum Mov Sci 18:121–136
Elliott D, Heath M, Binsted G, Ricker KL, Roy EA, Chua R (1999b) Goal-directed aiming: Correcting a force specification error with the right and left hands. J Mot Behav 31:309–324
Elliott D, Helsen WF, Chua R (2001) A century later: Woodworth’s (1899) two-component model of goal directed action. Psychol Bull 127:342–357
Fitts PM (1954) The information capacity of the human motor system in controlling the amplitude of movement. J Exp Psychol 48:303–312
Gan KC, Hoffman ER (1988) Geometrical condtions for ballistic and visually controlled movements. Ergonomics 31:829–839
Galati G, Lobel E, Giuseppe V, Berthoz A, Pizzamiglio L, Le Bihan D (2000) The neural basis of egocentric and allocentric coding of space in humans: a functional magnetic resonance study. Exp Brain Res 133:156–164
Georgopolos AP, Kalaska JF, Caminiti R, Massey JT (1983) Interruption of motor cortical discharge subserving aimed arm movements. Exp Brain Res 49:327–340
Goodale MA, Pelisson D, Prablanc C (1986) Large adjustments in visually guided reaching do not depend on vision of the hand and perception of the target displacement. Nature 320:748–750
Gréa H, Pisella L, Rossetti Y, Desmurget M, Tilikete C, Grafton S, Prablanc C, Vighetto A (2002) A lesion of the posterior parietal cortex disrupts on-line adjustments during aiming movements. Neuropsychologia 40:2471–2480
Handy TC, Grafton ST, Shroff NM, Ketay S, Gazzaniga MS (2003) Graspable objects grab attention when the potential for action is recognized. Nat Neurosci 6:421–427
Heath M (2005) Role of limb and target vision in the online control of memory-guided reaches. Motor Control (in press)
Heath M, Hodges NJ, Chua R, Elliott D (1998) On-line control of rapid aiming movements: unexpected target perturbations and movement kinematics. Can J Exp Psychol 52:163–173
Heath M, Weir PL, Roy EA (1999) Visual-motor integration of unexpected sensory events in young and older participants: a kinematic analysis. Dev Neuropsychol 16:197–211
Heath M, Westwood DA (2003) Does a visual representation support the online control of memory-dependent reaching? Evidence from a variable spatial mapping paradigm. Motor Control 7:346–361
Heath M, Westwood DA, Binsted G (2004) The control of memory-guided reaching movements in peripersonal space. Mot Control 8:76–106
Held R. Gottlieb N (1958) Technique for studying adaptation to disarranged hand-eye coordination. Percept Mot Skills 8:83–86
Keele SW (1968) Movement control in skilled motor performance. Psychol Bull 70:387–403
Khan MA, Elliott D, Coull J, Chua R, Lyons J (2002) Optimal control strategies under different feedback schedules: Kinematic evidence. J Mot Behav 32:45–57
Khan MA, Lawrence GP (2005) Differences in visuomotor control between the upper and lower visual fields. Exp Brain Res 164: 395–398
Krigolson O, Heath M (2004) Background visual cues and memory-guided reaching. Hum Mov Sci 23:861–877
Lakha L, Humphreys G (2005) Lower visual field advantage for motion segmentation during high competition for selection. Spat Vis 18:447–460
Langolf GD, Chaffin DB, Foulke JA (1976) An investigation of Fitts’ law using a wide range of movement amplitudes. J Mot Behav 8:113–128
MacKay WA (1992) Properties of reach-related neuronal activity in cortical area 7A. J Neurophysiol 67:1335–1345
MacKenzie CL, Marteniuk RG, Dugas C, Liske D Eickmeir B (1987) Three dimensional movement trajectory in a Fitts’ task: Implication for control. Q J Exp Psychol 18:629–647
Maunsell JH, Van Essen DC (1987) Topographic organization of the middle temporal visual area in the macaque monkey: representational biases and the relationship to callosal connections and myeloarchitectonic boundaries. J Comp Neurol 266:535–555
Meyer DA, Abrams RA, Kornblum S, Wright CE, Smith JEK (1988) Optimality in human motor performance: ideal control of rapid aimed movements. Psychol Rev 95:340–370
Mountcastle VB, Lynch JCGA, Georgopolous A, Sakata H, Acuna C (1975) Posterior parietal association cortex of monkey: command function for operations within extrapersonal space. J Neurophysiol 38:871–908
Oliveira FT, Elliott D, Goodman D (2005) Energy-minimization bias: compensating for intrinsic influence of energy-minimization mechanisms. Mot Control 9:101–114
Paulignan Y, Frak VG, Toni I, Jeannerod M (1997) Influence of object position and size on human prehension movements. Exp Brain Res 144: 226–234
Pelisson D, Prablanc C, Goodale MA, Jeannerod M (1986) Visual control of reaching movements without vision of the limb. II. Evidence of fast unconscious processes correcting the trajectory of the hand to the final position of a double-step stimulus. Exp Brain Res 62:303–311
Pisella L, Grea H, Tilikete C, Vighetto A, Desmurget M, Rode G, Boisson D, Rossetti Y (2000) An ‘automatic pilot’ for the hand in human posterior parietal cortex: toward reinterpreting optic ataxia. Nat Neurosci 3:729–736
Previc FH (1990) Functional specialization in the lower and upper visual fields in humans: its ecological origins and neurophysiological implications. Behav Brain Sci 13:519–575
Previc FH (1998) The neuropsychology of 3-D space. Psychol Bull 124:123–164
Stubbs DF (1976) What the eye tells the hand. J Mot Behav 8:43–48
Vince MA (1948) Corrective movements in a pursuit task. Q J Exp Psychol 1:85–103
Westwood DA, Goodale MA (2003) A haptic size-contrast illusion affects size perception but not grasping. Exp Brain Res 153:253–259
Woodworth RD (1899) The accuracy of voluntary movement. Psychol Rev, whole no. 13 (Monograph Supplement)
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Krigolson, O., Heath, M. A lower visual field advantage for endpoint stability but no advantage for online movement precision. Exp Brain Res 170, 127–135 (2006). https://doi.org/10.1007/s00221-006-0386-x
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00221-006-0386-x