
Abstract
In the present study participants searched for an onset target or a color singleton target and were required to execute a saccade toward (prosaccade) or away (antisaccade) from the search target. The results showed that participants often made erroneous saccades toward the onset or color singleton when they were the search target in the antisaccade condition, but not when they were presented as task-irrelevant distractors. This suggests that task-relevance plays a critical role in the production of erroneous prosaccades and provides evidence that these saccades are not completely reflexive. Furthermore, it was found that the antisaccade cost (latency difference between prosaccades and antisaccades) was greater for color singleton search targets than for onset search targets. The present findings have implications for our understanding of the processes involved in the programming of antisaccades and the causes of erroneous prosaccades.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
During our everyday lives we are continuously confronted with a complex environment containing a great deal of information. In order to interact with our environment according to our goals we need to visually select information that is relevant, while ignoring information that is irrelevant. A key issue in oculomotor research is the degree to which our eye movements (saccades) are governed by the goals and intentions of the observer or by the properties of the visual environment. Saccades that are controlled by the goals and intentions of the observer are called endogenous saccades (voluntary, goal-directed), while saccades that are controlled by the salient properties of the visual scene are called exogenous (reflexive, stimulus-driven) saccades.
A common task that is often used to examine exogenous and endogenous saccades is the anti-saccade task, developed by Hallet (1978; also see Hallet and Adams 1980). In this task a single onset stimulus is presented left or right of a central fixation point and observers are required to execute a saccade (an antisaccade) in the opposite direction from the onset. Correct antisaccades are considered endogenous, because they are directed to a specific location based on the task-instructions. Although participants are able to make correct antisaccades on the majority of trials, on about 5–30% of the trials (depending on the specific characterstics of the task; e.g. Fischer and Weber 1992, 1996; Mokler and Fischer 1999) they execute an erroneous prosaccade toward the onset. These erroneous prosaccades are generally considered exogenous (or reflexive; e.g. Guitton et al. 1985; Everling and Munoz 2000; Olk and Kingstone 2003; Pratt and Trottier 2005) since they are directed to a salient onset stimulus despite the instruction to move the eyes away from the onset.
In order to account for the findings in the antisaccade task Hallet and Adams (1980) proposed a model, in which a saccade is automatically programmed toward the onset. A goal redefinition process determines the correct antisaccade location. If this goal redefinition process starts before the programming of the prosaccade is completed a saccade to the antisaccade location is executed, otherwise an erroneous prosaccade occurs. Guitton et al. (1985) applied this model to antisaccade performance of frontal lesion patients. Their results showed that frontal lesion patients executed a higher proportion of erroneous prosaccades relative to a control group. They suggested that the frontal lobe damage delays the cancellation signal (or the goal redefinition process) such that it occurs too late to prevent the erroneous prosaccades on a large proportion of trials. In a more recent single-case study Walker et al. (1998) found that although a frontal lobe patient produced a high proportion of erroneous prosaccades in the antisaccade task his performance was only minimally impaired in a fixation task, in which the patient was required to maintain fixation and to ignore a peripheral onset. The delayed cancellation account of Guitton et al. appears unable to account for these results. Walker et al. suggested that the high error rate of the frontal patient in the antisaccade task is in part due to the higher working memory demands in the antisaccade task relative to the fixation task. Given the involvement of the frontal lobe in working memory (e.g. Goldman-Rakic 1987; Asaad et al. 1998; Rao et al. 1997) it is possible that higher task demands of the antisaccade task make this task more susceptible to working memory failure than the simple task of maintaining fixation.
Another task that has been developed to examine endogenous and exogenous control of saccades is the oculomotor capture task (e.g. Theeuwes et al. 1998, 1999; Kramer et al. 1999; Godijn and Theeuwes 2002). In their original study Theeuwes et al. (1999) presented participants with displays containing six gray circles spaced equally on an imaginary circle around a central fixation point. Centered within each circle was a small figure-eight premask. After 1 s all of the circles except one (the color singleton) changed their color to red and the premasks changed into small letters by removing some of their line segments. On half of the trials an additional irrelevant red circle (an abrupt onset) was added to the display simultaneously with the color change of the distractors. Participants were required to move their eyes to the uniquely colored gray circle and to determine whether the letter inside it was a ‘C’ or a ‘reversed-C’. Even though the onset was completely irrelevant for the task (and the same color as the other, non-onset distractors) the first saccade was directed to the onset on about a third of the trials. These erroneous saccades to salient task-irrelevant distractors have been referred to as oculomotor capture (e.g. Irwin et al. 2000). In a follow-up study Godijn and Theeuwes (2002) used a modified version of the oculomotor capture task by removing the manual response task, thereby turning the task into a pure oculomotor task. The finding of Theeuwes et al. (1998, 1999) that onsets captured the eyes on a large proportion of trials was replicated; on 29% of the trials the eyes initially went to onset before moving on to the color singleton target.
There appear to be some striking similarities between the oculomotor behavior in the antisaccade task and the oculomotor capture task. In both tasks participants often execute erroneous saccades to a salient onset, despite the instruction to execute a saccade to a different location. It is typically assumed that in order to execute a saccade (endogenously) to the goal location participants need to inhibit the execution of an exogenous saccade to the onset in both tasks (e.g. Godijn and Theeuwes 2002; Olk and Kingstone 2003). Furthermore, in both tasks saccades to the onset have shorter latencies than saccades to the goal location, saccades to the onset often undershoot their target (land somewhere between the central fixation point and the onset location) and the inter-saccade-interval (ISI) between the initial saccade to the onset and the following saccade to the goal location is typically relatively short (around 100 ms on average; Godijn and Theeuwes 2002; Everling and Fischer 1998).
In addition to the similarities between the antisaccade task and the oculomotor capture task there are also some differences. One important difference between the oculomotor capture task and the antisaccade task is the fact that in the oculomotor capture task the onset is completely irrelevant for the task, while in the antisaccade task the onset is task-relevant. In the oculomotor capture task there is no motivation for participants to pay attention to the onset, since the location at which the onset is presented is completely independent of the location of the color singleton target. However, in the antisaccade task the onset location determines the antisaccade location. That is, without determining the location of the onset an antisaccade cannot be programmed. Indeed, it is often reported that participants are sometimes unaware of the presence of an onset in the oculomotor capture paradigm (e.g. Theeuwes et al. 1999), but in the antisaccade task participants are necessarily aware of the onset. One recent study by Kramer et al. (2005) provides evidence that different processes may play a role in erroneous prosaccades and oculomotor capture. Kramer et al. (2005) found that the proportion of oculomotor capture trials is relatively stable from age 8 to 25, while the proportion of erroneous prosaccades in the antisaccade task decreases with age from age 8 to 16. Kramer et al. (2005) suggested that performance of the oculomotor capture task primarily involves the ability to exert top–down (or endogenous) control in opposition to attentional capture, whereas the antisaccade task involves the ability to maintain multiple task-sets. Specifically, in the antisaccade task the task-set of participants presumably includes attending to the onset, inhibiting a saccade to the onset and executing a saccade in the opposite direction. Kramer et al. (2005) interpreted their findings as evidence that the ability to exert top-down control over attentional capture already exists at age 8, but the ability to maintain multiple task-sets takes substantially longer to develop.
The differences between the oculomotor capture task and the antisaccade task suggest the possibility that prosaccade errors in the antisaccade task are of a somewhat different nature than oculomotor capture errors. Specifically, it is possible that prosaccade errors may have an endogenous component which is lacking in the oculomotor capture task. Specifically, participants performing the antisaccade task need to process the onset location due to its task-relevance. It may in fact be the case that the task-relevance of the onset in the antisaccade task plays a role in the production of erroneous prosaccades. From this perspective erroneous prosaccades are not genuinely exogenous or reflexive. The suggestion here is not that erroneous prosaccades are endogenous, but merely that endogenous factors may play a role in the production of erroneous prosaccades. In fact, the exogenous effect of onsets is not in dispute, since it has been well-established that onsets can have a strong exogenous effect on attention (Yantis 1996; Theeuwes 1995) and saccades (e.g. Theeuwes et al. 1998; Godijn and Theeuwes 2002; Kramer et al. 1999).
The goal of the present study is to examine the effect of task-relevance in the production of erroneous saccades and to examine the degree to which antisaccade performance is affected by whether participants are searching for an onset or a color singleton. In Experiment 1 participants viewed displays containing both a gray color singleton circle and a red onset circle presented together with two red non-onset circles (see Fig. 1 for an example of the display sequence). In separate blocks participants were either required to execute a prosaccade to the onset (pro-onset), an antisaccade away from the onset (anti-onset), a prosaccade to the color singleton (pro-color), or an antisaccade away from the color singleton (anti-color). Thus, the display configurations were the same in all conditions. In two conditions the onset was task-relevant and the color singleton was task-irrelevant, in the other two conditions the color singleton was task-relevant and the onset was task-irrelevant. This novel task enables a comparison between oculomotor capture and erroneous prosaccades. For example, in the pro-color block a saccade to the onset distractor is referred to as oculomotor capture, whereas in the anti-onset block a saccade to the onset is referred to as an erroneous prosaccade. Similarly, in the pro-onset block a saccade to the color singleton distractor is oculomotor capture, whereas, in the anti-color block a saccade to the color singleton is an erroneous prosaccade. If erroneous prosaccades are completely exogenous and the task-relevance of the search target plays no role in erroneous prosaccades, no difference in the frequency of erroneous prosaccades and oculomotor capture is expected. On the other hand, if task-relevance does play a role in the production of erroneous prosaccades, the frequency of erroneous prosaccades is expected to be higher than the frequency of oculomotor capture.

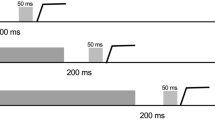
An example of the display sequence in Experiment 1. In the illustrated search display the saccade goal was the bottom center location in the pro-onset condition, the top center location in the anti-onset condition, the top left location in the pro-color condition and the bottom right location in the anti-color condition. See the text for further details
Previous studies examining oculomotor behavior with both onsets and color singletons have shown that onsets elicit more erroneous saccades than color singletons and saccade latencies to onsets are typically shorter than saccade latencies to color singletons. Therefore, in Experiment 1 it may be expected that more erroneous saccades will be elicited by the onset than by the color singleton. However, these previous studies have examined erroneous saccades to onsets and color singletons under conditions in which they were task-irrelevant. To the best of our knowledge there has been no direct comparison of antisaccade performance with onset targets versus color singleton targets.
Experiment 1 also allows a comparison between the antisaccade cost of an onset search target and that of a color singleton search target. The antisaccade cost is defined as the difference in mean saccade latency between correct antisaccades and correct prosaccades. It has been claimed that part of the antisaccade cost reflects the time needed to inhibit an erroneous prosaccade in the antisaccade task, resulting in a longer latency of antisaccades relative to prosaccades (e.g. Olk and Kingstone 2003; Pratt and Trottier 2005). Another factor that plays a role in the antisaccade cost is the endogenous requirement to compute the antisaccade goal based on the location of the peripheral onset (e.g. Olk and Kingstone 2003; Walker et al. 2000). It is also generally believed that onsets are especially effective at eliciting reflexive saccades (e.g. Pratt and Trottier 2005; Wu and Remington 2003). From this perspective it may be expected that the antisaccade cost is greater for onsets than for color singletons.
Experiment 2 is similar to Experiment 1, but with two important differences. First, participants only performed the pro-color and anti-color blocks. Second, an onset distractor was only presented on half the trials. This allows a comparison between prosaccade and antisaccade performance with and without an onset distractor.
Experiment 1
Methods
Participants
After giving their informed consent, eight students from the University of Illinois with normal or corrected-to-normal vision served as paid volunteers.
Apparatus
A Pentium-based computer with a 21-in color monitor controlled the timing of events and generated stimuli. Eye movements were recorded by means of an Eyelink II tracker. An eye movement was considered a saccade when the velocity exceeded 35°s-1 or the acceleration exceeded 9,500 °s-2. When participants were fixating the central fixation point at the start of each trial they pressed a key, which caused a recalibration of the participants’ gaze point to the central fixation point. After this the trial started. Each participant was tested in a dimly-lit room. They held their head on a chinrest, located 70 cm away from the monitor.
Stimuli, procedure and design
At the start of each trial participants viewed displays containing three gray circles (1.2° in diameter), which were randomly presented at three of six possible object locations around a central fixation point at an eccentricity of 7.6°. Two of the six possible object locations were directly above and below the central fixation point and the four other possible object locations were the corner positions of an imaginary square around the fixation point. At the three unfilled locations there were small white dots (0.2°). After 1,000 ms two of the three circles turned red, leaving one gray circle (the color singleton). At the same time a new red circle (the onset) appeared at one of the three unfilled locations. Thus, there was an equal location uncertainty between the color singleton and the onset (both one of three possible locations). See Fig. 1 for an example of the stimulus display.
There were four types of trials: on onset-pro trials the onset location was the saccade target. On onset-anti trials the location opposite the onset was the saccade target. On color-pro trials the location of the color singleton was the saccade target. Finally, on color-anti trials the location opposite the color singleton was the saccade target. Participants were required to execute a saccade toward the saccade target as quickly and accurately as possible. The search display was removed 500 ms after the saccade to the target. The four trial types were performed in separate blocks of 180 trials and each block was preceded by a practice session of 40 trials. The order of the blocks was counterbalanced across participants with the constraint that the search target was the same in the second block as in the first block and the task was the same in the third block as in the first block.
Results
Discarded data
Trials on which the initial saccade latency was below 80 ms (0.8% of trials) or above 1,000 ms (0.7% of trials) were discarded from further analyses.
Initial saccade destination
The distance between the initial saccade endpoint and the six possible object locations was used to determine the initial saccade destination. The saccade was assigned to the position closest to the saccade endpoint. In order to distinguish between the antisaccade location and the distractor location we excluded trials on which the distractor was presented at the antisaccade location. The data were collapsed across all directions. The average percentage of saccade errors per condition ranged between participants from 4.4 to 22.0%. Table 1 shows the percentage of initial saccades that were directed to each location in the pro-onset, pro-color, anti-onset and anti-color conditions. As can be seen in Table 1 there were more erroneous saccades on antisaccade trials than on prosaccade trials. In the pro-onset condition only 1.1% of the initial saccades were directed to the color singleton distractor, while in the anti-color condition 18.2% of the initial saccades were directed to the color singleton, t(7)=3.78, P<0.01. A similar pattern was found for the onset: In the pro-color condition only 2.0% of the initial saccades were directed to the onset distractor, while in the anti-onset condition 9.9% of the initial saccades were directed to the onset, t(7)=2.87, P<0.03. Furthermore, there was a trend towards more prosaccade errors to the color singleton than to the onset, t(7)=2.11, P<0.08. There was no significant difference in percentage of prosaccade errors toward the onset or the color singleton between the group of subjects who performed the antisaccade conditions first compared to those who performed the prosaccade conditions first (ts<1). A within-subjects analysis-of-variance (ANOVA) on the proportion of initial saccades to other locations (locations other than the color singleton, onset or antisaccade location) with search target (onset or color singleton) and task (prosaccade or antisaccade) as factors revealed a main effect of search target, F(1,7)=7.75, P<0.03, indicating that there were more erroneous saccades to other locations with a color singleton search target than with an onset search target. There was also a main effect of task, F(1,7)=81.92, P<0.005; there were more erroneous saccades to other locations in the antisaccade task than in the prosaccade task.
Saccade latency
The mean prosaccade and antisaccade latencies for each condition are shown in Fig. 2. Similar to the analysis of saccade destination we excluded trials on which the distractor was presented at the antisaccade location. An ANOVA with search target (onset or color singleton) and task (prosaccade or antisaccade) as factors was conducted on the mean latencies of the correct saccades. A main effect of search target was found, F(1,7)=20.89, P<0.005, indicating that the correct saccade latencies were shorter with an onset search target (mean 309 ms) than with a color singleton search target (mean 387 ms). There was also a main effect of task, F(1,7)=102.54, P<0.001; Correct prosaccade latencies were shorter (mean 279 ms) than antisaccade latencies (mean 416 ms). Finally, there was a task × search target interaction, F(1,7)=10.83, P<0.02, indicating that the antisaccade cost (difference in mean saccade latency between the pro and anti conditions) was greater for color singleton search targets (mean antisaccade cost = 179 ms) than for onset search targets (mean antisaccade cost = 96 ms).

Mean saccade latencies of Experiment 1
A further ANOVA with task (prosaccade or antisaccade) and search target (onset or color singleton) as factors on the mean latencies of prosaccades revealed a main effect of task, F(1,7)=5.02, P<0.02, which indicated that prosaccades had shorter latencies in the prosaccade conditions than in the antisaccade conditions.
Since the objects were presented at random locations around the central fixation point, on some trials an object was presented at the antisaccade location, while on other trials the antisaccade location was empty (the empty locations were marked with a small white dot). Planned comparisons revealed that there was no significant difference in mean antisaccade latency between trials on which an object was presented at the antisaccade location and trials on which the antisaccade location was empty (anti-onset condition t(7)=1.62, P>0.10, anti-color condition t(7)<1).
Inter-saccade-interval
When participants erroneously execute a prosaccade on antisaccade trials fixation durations on the prosaccade location tend to be relatively short prior to the correct saccade to the antisaccade location. In the present experiment we found a mean ISI of 139 ms in the anti-color condition and a mean ISI of 160 ms in the anti-onset condition. This difference did not reach significance, t(7)=1.13, P>0.25.
Discussion
The results of Experiment 1 showed that there were many more erroneous saccades toward the onset or color singleton when they were presented as search target in the antisaccade conditions than when they were task-irrelevant and presented as distractors. Furthermore, the antisaccade cost was greater for color singleton search targets than for onset search targets. These findings are clearly inconsistent with the view that erroneous prosaccades are purely exogenous. If that were the case there should have been no difference in frequency between erroneous prosaccades and oculomotor capture. One possible explanation for the finding of a greater antisaccade cost with color singletons than with onsets, is that the attentional demands are higher when searching for a color singleton than when searching for an onset. If there is a stronger attentional allocation at the color singleton search target than at the onset search target it may take more time to disengage attention and switch attention to the antisaccade location in order to execute an antisaccade in the anti-color condition than in the anti-onset condition. However, an alternative explanation is that the antisaccade cost is greater with a color singleton search target, because participants need to inhibit a saccade to the onset distractor. Previous research has shown that onsets have a strong effect on oculomotor behavior when participants search for a color singleton (e.g. Theeuwes et al. 1998; Godijn and Theeuwes 2002; Kramer et al. 1999). It is typically assumed that participants are required to inhibit a saccade to the onset distractor. Therefore, when participants are required to execute an antisaccade away from the color singleton they not only need to inhibit a prosaccade, but also a saccade to the onset distractor. In contrast, color singletons do not appear to have much effect on oculomotor behavior when subjects search for an onset target (Irwin et al. 2000). Since an onset distractor was always presented when subjects searched for a color singleton and a color singleton distractor was always presented when subjects searched for an onset, the difference between onset distractors and color singleton distractors may have contributed to the greater antisaccade cost for color singleton search targets. This idea is examined in Experiment 2.
Experiment 2
In order to examine whether the presence of an onset distractor has an effect on the antisaccade cost Experiment 2 examined the antisaccade cost with color singleton search targets with and without an onset distractor. The pro-color and anti-color conditions of Experiment 1 were repeated with the difference that an onset distractor was only presented on half of the trials.
Method
Participants
After giving their informed consent, ten students of the University of Illinois with normal or corrected-to-normal vision served as paid volunteers.
Stimuli, procedure and design
The stimuli were the same as in Experiment 1 with one exception: On half of the trials an onset was randomly presented at one of the unfilled locations, on the other half of the trials there was no onset. There were four types of trials: on prosaccade-onset present trials the saccade target was the color singleton and no onset distractor was presented. On prosaccade-onset absent trials the saccade target was the color singleton and an onset distractor was presented. On antisaccade-onset present trials the location opposite the color singleton was the saccade target and an onset distractor was presented. On antisaccade-onset absent trials the location opposite the color singleton was the saccade target and no onset distractor was presented. Each participant performed one block of prosaccade trials and one block of antisaccade trials. Half of the participants started with the prosaccade block, the other half started with the antisaccade block. There were 360 trials per block and each block was preceded by a practice block of 40 trials. The order of the onset present and onset absent trials was randomized within blocks.
Results
Discarded data
Trials on which the initial saccade latency was below 80 ms (1.0% of trials) or above 1,000 ms (0.3% of trials) were discarded from further analyses.
Initial saccade destination
Table 2 shows the percentage of initial saccades that were directed to each location in the Prosaccade and Antisaccade Conditions separately for onset present and onset absent trials. The average percentage of saccade errors per condition ranged between participants from 10.0 to 33.9%. Similar to Experiment 1 there were very few trials on which the eyes went to the onset distractor (2.8 and 3.7%), but there was a relatively high percentage of erroneous prosaccades to the color singleton target. There was no significant difference in percentage of erroneous prosaccades between the no onset and onset trials, t(9)<1. An ANOVA was conducted on the proportion of initial saccades to other locations (locations other than the color singleton, onset or antisaccade location) with Task (Prosaccade or Antisaccade) and Onset Presence (Present or Absent) as factors. There was a main effect of Task, F(1,9)=10.57, P<0.01, indicating that there were more erroneous saccades to other locations on antisaccade trials than on prosaccade trials. There was no significant effect of Onset, F(1,9)=3.30, P>0.10, nor was there a Task × Onset Presence interaction, F(1,9)<1.
Saccade latency
The mean saccade latencies are shown in Fig. 3. An ANOVA on the mean latencies of the correct saccades with Task (Prosaccade or Antisaccade) and Onset Presence (Present or Absent) as factors revealed a main effect of task, F(1,9)=91.87, P<0.001. Correct saccade latencies were shorter in the prosaccade condition than in the antisaccade condition. There was no main effect of onset presence, F(1,9)=1.84, P>0.20, but there was an interaction between task and onset presence, F(1,9)=17.33, P<0.005; The antisaccade cost was greater when an onset was present 27 ms (142–115) than when there was no onset.

Mean saccade latencies of Experiment 2
An ANOVA was also conducted on prosaccade latencies with Task (Prosaccade and Antisaccade) and Onset Presence as factors. The results showed a main effect of task, F(1,9)=7.50, P<0.03, indicating that prosaccade latencies were shorter in the prosaccade condition (mean 299 ms) than in the antisaccade condition (mean 321 ms).
Inter-saccade-interval
The mean ISI between initial erroneous prosaccades and antisaccades was 122 ms when there was an onset present and 137 ms when there was no onset. This difference in ISI was reliable, t(9)=2.53, P<0.04.
Discussion
The main result of Experiment 2 was that the antisaccade cost was greater when an onset distractor was present than when there was no onset distractor present. This provides support for the idea that the programming of an antisaccade or the inhibition of an erroneous prosaccade is more time-consuming when a saccade to an onset also needs to be inhibited. However, the presence of an onset had an effect of 27 ms on the antisaccade cost, while the difference in antisaccade cost between the color singleton search target condition and the onset search target condition in Experiment 1 was 83 ms. Therefore, it is unlikely that presence of an onset distractor is the only factor contributing to the difference in antisaccade cost between color singletons and onsets. This issue will further be discussed in the section General discussion.
The results of Experiment 2 replicated the findings from the pro-color and anti-color conditions of Experiment 1. That is, similar to Experiment 1 there were very few erroneous saccades to the onset distractor, but the large majority of erroneous saccades were prosaccades directed to the color singleton search target.
General discussion
The present study examined observers’ ability to execute prosaccades and antisaccades in response to onset and color singleton search targets. The displays of Experiment 1 always contained both an onset and a color singleton thereby allowing a comparison between oculomotor capture (erroneous saccade to a task-irrelevant distractor) and erroneous prosaccades (prosaccades in the antisaccade task). The results revealed a much higher frequency of erroneous prosaccades than oculomotor capture. That is, there were more erroneous saccades to the onset or the color singleton when they were task-relevant (i.e. the search target in the antisaccade task) than when they were task-irrelevant (i.e. a distractor in the prosaccade or antisaccade conditions). Furthermore, a greater antisaccade cost was found for color singleton search targets than for onset search targets. Finally, the antisaccade cost for color singleton search targets was slightly greater when an onset distractor was presented than when there was no onset distractor.
Endogenous component of erroneous prosaccades
The proportion of erroneous prosaccade errors ranged from approximately 10 to 22%, while the proportion of oculomotor capture only ranged from approximately 1 to 4%. This finding provides evidence for an endogenous component to the production of erroneous prosaccades. That is, erroneous prosaccades are not merely executed in a stimulus-driven manner, based on the saliency of the search target. Instead, the results indicate that the task demands play an important role in erroneous prosaccade execution.
In accordance with previous studies we propose that the working memory contents determine to a large extent the probability that an erroneous saccade is executed. First of all, participants in the present study are required to keep the identity of the search target in working memory. They are required to localize either the color singleton or the onset. Moreover, in the antisaccade task they are instructed not to execute a saccade to this search target, but instead to execute a saccade in the opposite direction. Thus, the requirement to suppress a saccade to the search target is kept in working memory. However, outside of the laboratory visual search is typically conducted in an overt fashion. That is, although it is clear that observers can perform search covertly, searching through a cluttered visual environment under natural conditions involves the execution of saccades (see Findlay and Gilchrist 2003, for a review); we usually look at what we’re attending. Finally, because participants were instructed to execute speeded responses participants keep in working memory the requirement to execute a speeded saccade to the target location.
Although these task demands do not result in an especially high working memory load, the requirement for speeded responses does pose a challenge to the application of the working memory contents. Furthermore, given the natural tendency to overtly respond to the search target the relatively frequent erroneous prosaccades should not be surprising. The present results not only suggest that the task demands play a role in erroneous prosaccades, but also in the execution of other erroneous saccades. That is, in addition to the erroneous prosaccades participants also executed erroneous saccades to other locations (to locations other than those occupied by the onset or the color singleton). The results showed that the proportion of these errors depended on the task demands; there were more saccade errors to other locations with a color singleton target than with an onset target and there were more in the antisaccade task than in the prosaccade task. This suggests that participants followed the speed instruction and allowed themselves a limited time to covertly determine the correct saccade location.
The idea that working memory contents play a role in the production of erroneous prosaccades is consistent with a number of previous studies. First, Roberts et al. (1994) showed that the percentage of erroneous prosaccades was increased when the antisaccade task was combined with a concurrent working memory task. Thus, when working memory is overloaded participants are often unable to prevent executing an erroneous prosaccade. Second, several lesion studies have shown that patients with frontal lesions such as the dorsolateral prefrontal cortex (e.g. Pierrot-Deseilligny et al. 1991, 2002) and the anterior cingulate cortex (Milea et al. 2003) have an increased percentage of erroneous prosaccades. Since there is overwhelming evidence that areas of the frontal cortex are involved in working memory (e.g. Goldman-Rakic 1987; Asaad et al. 1998; Rao et al. 1997) these findings provide further evidence that maintaining multiple task-sets in working memory is crucial for correct antisaccade performance and that working memory failure in this task often results in erroneous prosaccades. Furthermore, Walker et al. (1998) in a single-case study showed that although a patient with a lesion of the frontal cortex was unable to prevent erroneous prosaccades in the antisaccade task he performed much better in a fixation task, in which a peripheral stimulus was presented and the patient was instructed to remain centrally fixated (and could thus ignore the peripheral stimulus). Walker et al. proposed that the greater impairment of the frontal patient in the antisaccade task compared to the fixation task was the result of greater working memory demands. A third line of evidence was recently provided by Kramer et al. (2005). The results of this study showed that the percentage of erroneous prosaccades declined from age 8 to 16, while the percentage of oculomotor capture remained relatively stable. Since working memory continues to develop into the mid-teens, these findings provide evidence that correct antisaccade performance and the inhibition of prosaccades relies on the ability to keep multiple task-sets in working memory.
Antisaccade costs
Previous research has shown that antisaccade latencies are typically longer than prosaccade latencies. The additional cost in programming an antisaccade, which we call the antisaccade cost, has been related to the time required to inhibit a reflexive saccade to the onset stimulus (e.g. Pratt and Trottier 2005; Olk and Kingstone 2003). Specifically, it is argued that before an antisaccade can be programmed the prosaccade must be inhibited. Olk and Kingstone (2003) provided evidence for this view by inducing oculomotor inhibition to prosaccades by requiring participants to identify the prosaccade stimulus. Based on its orientation participants were either required to execute a prosaccade or an antisaccade. It was assumed that this manipulation equated the degree to which oculomotor inhibition was applied between the prosaccade and antisaccade conditions. That is, participants were required to inhibit a saccade to the prosaccade stimulus in both prosaccade and antisaccade conditions in order to determine the orientation of the prosaccade stimulus. It was found that this manipulation reduced the antisaccade cost, but it did not completely disappear. Olk and Kingstone (2003) concluded that the time needed to inhibit a prosaccade accounts for a large proportion of the antisaccade cost.
In the present study we found that the antisaccade cost was greater for color singleton search targets than for onset search targets. This finding seems surprising given the assumption that onsets have a stronger exogenous effect on the oculomotor system (this idea is consistent with our finding of shorter saccade latencies to onset than to color singletons) and that it should therefore be more time-consuming to inhibit a prosaccade to an onset than a prosaccade to a color singleton. One possible explanation for these results is that the speed of inhibition depends to some extent on the task demands. That is, the localization of the onset is relatively straightforward compared to localization of the color singleton (evidence for this idea is provided by the shorter latency prosaccades in the onset condition relative to the color singleton condition) and it is possible that the cancellation of the prepotent response to the search target is facilitated by a relatively fast localization.
Another finding in the present study consistent with the idea that oculomotor inhibition is partly responsible for the antisaccade cost is the finding that the antisaccade cost for color singleton targets is greater when an onset distractor is presented than when there is no onset distractor. It is assumed that inhibitory mechanisms are also involved in preventing a saccade to the onset distractor (e.g. Godijn and Theeuwes 2002). Therefore, the presence of an onset distractor provides an additional load on the inhibitory mechanisms resulting in a reduction of the efficiency with which a prosaccade can be inhibited thereby increasing the antisaccade cost.
References
Asaad WT, Rainer G, Miller EK (1998) Neural activity in the primate prefrontal cortex during associative learning. Neuron 21:1399–1407
Everling S, Fischer B (1998) The antisaccade: a review of basic research and clinical studies. Neuropsychologica 36:885–899
Everling S, Munoz DP (2000) Neuronal correlates for preparatory set associated with pro-saccades and anti-saccades in the primate frontal eye field. J Neurosci 20:387–400
Findlay JM, Gilchrist ID (2003) Active vision: the psychology of looking and seeing. Oxford University Press, Oxford
Fischer B, Weber H (1992) Characteristics of “anti” saccades in man. Exp Brain Res 89:415–424
Fischer B, Weber H (1996) Effects of precues on error rate and reaction times of antisaccades in human subjects. Exp Brain Res 109:507–512
Godijn R, Theeuwes J (2002) Programming of endogenous and exogenous saccades: evidence for a competitive integration model. J Exp Psychol Hum Percept Perform 28:1039–1054
Goldman-Rakic P (1987) Circuitry of primate prefrontal cortex and regulation of behavior by representational memory. In: Plum F (ed) Handbook of physiology: the nervous system. American Physiology Society, Bethesda pp 373–417
Guitton D, Buchtel HA, Douglas RM (1985) Frontal lobe lesions in man cause difficulties in suppressing reflexive glances and in generating goal directed saccades. Exp Brain Res 58:455–472
Hallet PE (1978) Primary and secondary saccades to goals defined by instructions. Vision Res 18:1279–1296
Hallet PE, Adams WD (1980) The predictability of saccade latency in a novel oculomotor task. Vision Res 20:329–339
Irwin DE, Colcombe AM, Kramer AF, Hahn S (2000) Attentional and oculomotor capture by onset luminance and color singletons. Vision Res 40:1443–1458
Kramer AF, Gonzalez de Sather JCM, Cassavaugh ND (2005) Development of attentional and oculomotor control. Dev Psychol 41:760–772
Kramer AF, Hahn S, Irwin DE, Theeuwes J (1999). Attentional capture and aging: implications for visual search performance and oculomotor control. Psychol Aging 14:135–154
Milea D, Lehericy S, Rivaud-Pechoux S, Duffau H, Lobel E, Capelle L, Marsault C, Berthoz A, Pierrot-Deseilligny Ch (2003) Antisaccade deficit after anterior cingulate cortex resection. Neuroreport 14:283–287
Mokler A, Fischer B (1999) The recognition and correction of involuntary prosaccades in an anti-saccade task. Exp Brain Res 125:511–516
Olk B, Kingstone A (2003) Why are antisaccades slower than prosaccades? A novel finding using a new paradigm. Neuroreport 14:151–155
Pierrot-Deseilligny C, Rivaud S, Gaymard B, Agid Y (1991) Cortical control of reflexive visually-guided saccades. Brain 114:1473–1485
Pierrot-Deseilligny C, Ploner CJ, Muri RM, Gaymard B, Rivaud-Pechoux S (2002) Effects of cortical lesions on saccadic eye movements in humans. Ann N Y Acad Sci 956:216–229
Pratt J, Trottier L (2005) Pro-saccades and anti-saccades to onset and offset targets. Vision Res 45:765–774
Rao SC, Rainer G, Miller EK (1997) Integration of what and where in the primate prefrontal cortex. Science 276:821–824
Roberts RJ, Hager LC, Heron C (1994) Prefrontal cognitive processes: working memory and inhibition in the antisaccade task. J Exp Psychol Gen 123:374–393
Theeuwes J, Kramer AF, Hahn S, Irwin DE (1998) Our eyes do not always go where we want them to go: capture of the eyes by new objects. Psychol Sci 9:379–385
Theeuwes J, Kramer AF, Hahn S, Irwin DE, Zelinsky GJ (1999) Influence of attentional capture on oculomotor control. J Exp Psychol Hum Percept Perform 25:1595–1608
Walker R, Husain M, Hodgson TL, Harrison J, Kennard C (1998) Saccadic eye movement and working memory deficits following damage to human prefrontal cortex. Neuropsychologia 36:1141–1159
Walker R, Walker DG, Husain M, Kennard C (2000) Control of voluntary and reflexive saccades. Exp Brain Res 130:540–544
Wu SC, Remington RW (2003) Characteristics of covert and overt visual orienting: evidence from attentional and oculomotor capture. J Exp Psychol Hum Percept Perform 29:1050–67
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Godijn, R., Kramer, A.F. Prosaccades and antisaccades to onsets and color singletons: evidence that erroneous prosaccades are not reflexive. Exp Brain Res 172, 439–448 (2006). https://doi.org/10.1007/s00221-006-0351-8
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00221-006-0351-8