
Abstract
The latency of eye movements is influenced by the fixation task; when the fixation stimulus is switched off before the target presentation (gap paradigm) the latency becomes short and express movements occur. In contrast, when the fixation stimulus remains on when the target appears (overlap paradigm), eye movement latency is longer. Several previous studies have shown increased rates of express saccades in children; however the presence of an express type of latency for vergence and combined movements in children has never been explored. The present study examines the effects of the gap and the overlap paradigms on horizontal saccades at far (150 cm) and at close (20 cm) viewing distances, on vergence along the median plane, and on saccades combined with convergence or divergence in 15 normal seven-year-old children. The results show that the gap paradigm produced shorter latency for all eye movements than the overlap paradigm, but the difference was only significant for saccades at close viewing distances, for divergence (pure and combined), and for saccades combined with vergence. The gap paradigm produced significantly higher rates of express latencies for saccades at close viewing distances, for divergence, and for saccades combined with divergence; in contrast, the frequencies of express latencies for saccades at far viewing distances and for convergence (pure or combined) were similar in the gap and the overlap paradigms. Interestingly, the rate of anticipatory latencies (<80 ms) was particularly high for divergence in the gap paradigm. Our collective findings suggest that the initiation of saccades at close viewing distances and of divergence is more reflexive, particularly in the gap paradigm. The finding of frequent anticipatory divergence that occurs at similar rates for seven-year-old children (this study) and for adults (Coubard et al., 2004, Exp Brain Res 154:368–381) indicates that predictive initiation of divergence is dominant.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
To perform an eye movement, several cortical and sub-cortical areas are activated; the visual information from the retina is sent to the visual cortex, parietal cortex, frontal lobe, and the superior colliculus, and from there, via the brain-stem, the motor command is sent to the extraocular muscles (Leigh and Zee 1999). When preparing for an eye movement, several processes take place, such as shift of the visual attention to the new target, disengagement of oculomotor fixation, and computation of the parameters of the movement (see Fischer and Ramsperger 1984; Findlay and Walker 1999).
It is well known that the time needed to prepare for an eye movement is influenced by the fixation task; the saccade latency when the fixation stimulus is turned off before the onset of the target (gap paradigm) is shorter than the latency measured when the fixation stimulus remains present when the target appears (overlap paradigm). Another phenomenon related to the gap is the facilitation of the occurrence of express saccades—saccades with latencies between 80 and 120 ms (Saslow 1967; Fischer and Boch 1983; Fischer and Ramsperger 1984; Fischer and Weber 1993).
The shortening of saccade latency in the gap paradigm could be due to facilitation of the disengagement of fixation and attention (Reuter-Lorenz et al 1991; Kingstone and Klein 1993). This idea is integrated into models like that of Findlay and Walker (1999), who suggested that there is a decrease in fixation activity during the stimulus offset that promotes rapid movement initiation. Nevertheless, the mechanisms underlying the occurrence of express saccades are still controversial. Indeed, some authors have proposed the possibility of a specific short cortical–subcortical circuit facilitated by release of visual fixation for express saccade generation (for a review, see Pierrot-Deseilligny et al 1995, 2002), while other authors, using results from physiological studies in monkeys (Pare and Munoz 1996; Dorris et al 1997), have advanced the so-called ’oculomotor preparation” hypothesis in which a predictive saccade command could be the origin of express saccades.
The latency of saccades has been extensively studied in children (Ross et al 1994; Biscaldi et al 1996; Fischer et al 1997; Munoz et al 1998; Fukushima et al 2000; Klein and Foerster 2001). All of these studies have shown that saccade latency is longer in children than in adults, and that its shortens with age. This has been attributed to underdeveloped cortical structures involved in eye movement preparation. This finding is confirmed by the fRMI study of Luna et al (2001), which investigated the brain activity in 8–30-year-old subjects during pro- and antisaccades tasks. Activation of the frontal, parietal, striatal and thalamic regions increased progressively between childhood and adolescence.
Express saccades have also been studied extensively in children. In particular, Munoz et al (1998) showed that young children (5–8 years old) make frequent express saccades (18%); Klein and Foerster (2001) also examined the gap effect in children and adults (6–26 years old), and they found that express saccades are more frequent in young children (6–11 years old) than in adults (18–26 years old): 10% versus 5%. These authors suggested that the high number of express saccades, together with the high variability of saccade latency observed in younger children, could be due to poor control of the fixation system; fixation cells in the superior colliculus are found to have a mutually inhibitory relationship with visuo-motor cells (Munoz and Wurtz 1993).
All of these behavioral studies dealing with the gap effect are limited to saccades, and there are no studies that examine vergence latency as well as combined (saccades–vergence) eye movements in children. The goal of the present study was to explore the effects of the gap and overlap paradigms on the latency of saccades, vergence and combined saccade–vergence movements in a population of school children aged between six and seven years old. According to the above studies (Munoz et al 1998; Klein and Foerster 2001), mean saccade latency is longer in children of that age than in adults, but the rate of express latencies is higher in the former. We aimed to determine whether similar phenomena—increased mean latency in children and increased rates of express latencies—occur for the other types of eye movements (for pure vergence or that combined with saccades).
A few studies have explored the influence of the gap/overlap effect on vergence latency and on the rate of express vergence in adult subjects (Tam and Ono 1994; Takagi et al 1995; Coubard et al 2004). Interestingly, Coubard et al (2004) showed that the gap paradigm shortens the overall latency of both saccades and vergence, but the rate of express vergence is different to that of express saccades and it depends on the type of vergence (1 and 13% for convergence and divergence, respectively). This finding suggests the presence of distinct cortical structures controlling the preparation of saccades and the two types of vergence.
Given the immaturity of the fixation system in young children, according to the hypotheses of Fischer and Weber (1993), Munoz et al (1998) and Klein and Foerster (2001), we expected to find a high percentage of express eye movements (saccades, vergence and combined movements). This was not the case; rather, our findings suggest the presence of distinct gap effects depending on the type of eye movement.
Materials and methods
Subjects
Fifteen children from the first grade of an elementary French school participated in the study. The mean age was 7.1±0.5 years. The investigation adhered to the principles of the Declaration of Helsinki and was approved by our institutional human experimentation committee. Informed consent was obtained from the children’s parents after the procedure for the experiment was explained. All of the children underwent a complete ophthalmologic/orthoptic examination.
All children had normal binocular vision (60 seconds of arc or better), which was evaluated with the TNO (Test of Netherlands Organisation) for stereoscopic depth discrimination. This test is based on random dot stereograms that contain a distribution of binocular disparity; it provides retinal disparities ranging from 15 to 480 seconds of arc (see von Noorden and Campos 2002). One child (S3) wore glasses (hypermetropia of 0.5 for both eyes). Visual acuity was normal (≥ 9/10) for all children at both near and far distances. Orthoptic evaluation of vergence (performed using prisms and a Maddox rod) was in the normal range with respect to reference values based on clinical studies (Ygge et al 1993; Evans 1997; von Noorden and Campos 2002); distant near point of convergence was on average ≤7 cm; exophoria (latent deviation of one eye when the other eye is covered) at near viewing was less than six prism diopters for all children; the accommodative convergence/accommodation ratio, AC/A, was 5.3±0.96: only one child (S14) had an AC/A ratio ≥6.
Oculomotor paradigm
Spatial arrangement
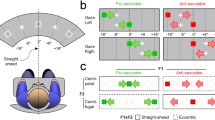
The spatial arrangement is shown in Fig. 1a. A computer controlled the LED display. LEDs were placed in two isovergence circles at different distances (at 20 cm and 150 cm) from the child. Three LEDs were placed on the circle closest to the child, one at the center and the others at ±20°. The required mean vergence angle for fixating any of these three LEDs on the far circle was 17°. Five LEDs were placed on the circle most distant from the child: one at the center, two at ±10° and two at ± 20°; fixation to any of these LEDs required a vergence angle of 2.3°. To change fixation from the distant to the close LEDs requires a large vergence change, and a sustained vergence and accommodative effort at near distance (20 cm) was demanded of the child. However, such situations are common in everyday life, particularly at school, where the child frequently changes their fixation between far distances (blackboard) and close distances (desk, book).

A Spatial arrangement: different types of eye movements are elicited depending on the combination of the fixation and target LEDs: pure saccades at far (150 cm) and at close (20 cm) viewing distance, pure vergence (convergence and divergence) along the median plane, and combined saccade–vergence movements. B Temporal arrangement: schematic diagram of the temporal arrangement used in the two different paradigms (gap and overlap)
Three types of eye movements were elicited: pure saccades rightward or leftward, pure vergence along the median plane (convergence or divergence), and combined movements. At the start of each trial, a fixation LED was lit up at the center of one of the two circles (far or close viewing). When the target LED was on the same circle it called for a pure saccade (rightward or leftward) at far (150 cm) or at close (20 cm) viewing distance. When the target LED was on the center of the other circle it called for a pure vergence eye movement along the median plane (convergence or divergence), and when it was lateral and on the other circle the required eye movement was a combined saccade and vergence eye movement. The required saccade amplitude was always 20°, and the required vergence change along the median plane (for pure vergence) or along lateral axes (for combined movements) was 15°.
Temporal arrangement
Two temporal paradigms (gap and overlap) were used (see Fig. 1b). At the start of each trial, a fixation LED was lit up at the center of one of the circles, which remained lit for 2.5 s. In the gap paradigm, after this fixation period, the central LED was turned off, and there was a 200 ms period where no LEDs were illuminated (gap). At the end of this period, a target LED appeared for 1.5 s. A delay of 0.5 s was introduced before the next trial.
The overlap paradigm began similarly, with the central fixation LED illuminated for 2.5 s. In this case, the central fixation LED remained illuminated for a further 200 ms, and then the target LED appeared (overlap), so that during the overlap period both the fixation and the target were visible. The target LED remained illuminated for 1.5 s. A delay of 0.5 s was introduced before the next trial. The gap and the overlap trials were randomly mixed.
Recording eye movements
Data collection was controlled via REX, software developed for real-time experiments and visual display, run on a PC. Horizontal eye movements from both eyes were recorded simultaneously with a photoelectric device (Oculometer, Bouis). This system has an optimal resolution of 2′′ of arc. Eye-position signals were low-pass filtered with a cutoff frequency of 200 Hz and digitized with a 12-bit analogue-to-digital converter; each channel was sampled at 500 Hz.
Oculomotor task
The child was seated in a chair with a head and chin support in a dark room. He/she faced a horizontal surface at eye level where the LEDs were positioned; viewing was binocular.
The child performed a sequence of saccades to the target LEDs, moving from 0° to ±10° or 0° to ± 20° on the far isovergence circle. During each of these trials the target LEDs remained on at each location for 2 s. The child was instructed to fixate the target LED as accurately as possible; the duration of the target LED presentation was sufficiently long to allow accurate and stable fixation. The calibration factors were extracted from these recordings.
Each child performed four blocks, each block separated by a few minutes of rest. In each block, the three types of trials (saccades, vergence and combined saccade–vergence movements) were interleaved randomly. Each block contained 36 trials—six saccades at far viewing distance (three rightwards and three leftwards), six saccades at close viewing distance (three rightwards and three leftwards), six pure convergence (along the median plane), six pure divergence, six saccades combined with divergence, and six saccades combined with convergence. Calibration was repeated before and after each block.
Data analysis
The analysis methods employed were similar to those used in prior studies (Yang et al 2002; Bucci et al 2003). Briefly, a linear function was used to fit the calibration data. From the two individual calibrated eye position signals we calculated the conjugate [(left eye + right eye)/2] and the vergence (left eye − right eye) signal. Figure 2 shows an example of pure rightward saccades (A), pure divergence (B) and rightward saccades combined with divergence (C). Markers were placed at different points in the eye position signals automatically by the computer; an investigator verified the placement of markers; and, most important, excluded the trials in which the movement was in the wrong direction with respect to the stimulus or was contaminated by blinks. The onset of the conjugate saccadic component was defined as the time when the eye velocity reached 5% of the saccadic peak velocity. Onset of the vergence signal (for pure vergence movements and for the vergence component of the combined movements) was defined as the time point when the eye velocity exceeded or dropped by 5°/s. These criteria are standard and similar to those used by other authors (Takagi et al 1995; Bucci et al 2003; Yang et al 2002). Like in other studies (such as Klein and Foerster 2001), the upper limit of latency was set to 700 ms. The percentage of movements rejected (blinks, movements in wrong direction, movements with long latencies) ranged between 8–15% and 10–16% in the gap and in the overlap conditions respectively. Anticipatory movements—those with latencies shorter than 80 ms—were not included in the analysis; the percentage of such movements was less than 10% for all types of eye movement except for one (divergence, 14%). Again these criteria are standard and have been used by others (Takagi et al 1995; Munoz et al 1998; Klein and Foerster 2001; Yang et al 2002; Coubard et al 2004). The percentage of rejected trials was similar for all types of eye movements and for the two conditions (gap/overlap). Express latency for vergence (in the pure as well as combined forms) was defined as having the same latency as express saccades (80–120 ms); this is consistent with other studies (see Takagi et al 1995; Coubard et al 2004).

Examples of different types of recorded eye movements: pure rightward saccade (A), pure divergence (B) and rightward saccade combined with divergence (C). The excursion of the target is shown on the right side
Fischer and Ramsperger (1984) were the first to observe a distinct population of saccades with short latency in adults, the so-called express saccades. Subsequent studies in children dealing with the influence of the gap effect on saccade latency used the same criterion as Fischer and Ramsperger (1984), even though saccade latencies in children are on average longer than those found in adults (Munoz et al 1998; Klein and Foerster 2001). Thus, the use of the same criterion for express saccades (80–120 ms) is valid. Concerning the definition of express latency for vergence, the same arbitrary criterion was used by others (Takagi et al 1995; Coubard et al 2004). In the present study we use the same criterion and this allows us to compare our data with other studies that have examined the influence of the gap effect on vergence movements in adults.
For each type of eye movement (saccade, vergence and combined movements), we measured the latency (the time between the onset of the target LED and the beginning of the movement) in ms. The latency was measured for each component (saccade and vergence component) for combined movements. The percentage of express movements (with latencies between 80 ms and 120 ms) and anticipatory movements (with latencies shorter than 80 ms) was calculated. We performed a non-parametric analysis of mean latency (Friedman) to compare the gap and the overlap paradigms and also to compare the numbers of express and anticipatory movements.
Results
Pure saccades
Figure 3 shows the saccade latency distributions at far (A) and at close (B) viewing distances for both gap and overlap paradigms. The mean latencies of saccades at far and at close viewing distances are shorter in the gap paradigm than in the overlap paradigm; the mean values are 284±69 ms and 233±98 ms, respectively, for saccades at far and at close viewing distances in the gap paradigm, and 345±100 ms and 305±100 ms in the overlap paradigm. The mean latency difference between the two paradigms (gap–overlap) is 61 ms for saccades at far and 72 ms for saccades at close viewing distances, and is only statistically significant for saccades at close viewing distance (χ2 (15,1)=5.45, P=0.020).

Latency distribution in the gap (solid line) and overlap (dotted line) paradigms for saccades at far (A) and at close (B) viewing distances. The bin width is 20 ms. The shadow zone in the graph indicates express latencies between 80 and 120 ms. The mean latencies and the standard deviations in the gap and in the overlap paradigms are also indicated
Is the short latency observed in the gap paradigm due to a higher rate of express saccades? For saccades made at far viewing distance, the percentage of express saccades is similar in the two paradigms: only 5 and 3%, respectively in the gap and overlap paradigms. In contrast, for saccades performed at close viewing distance in the gap paradigm, the percentage of express saccades is significantly higher (χ2 (15,1)=6.44, P=0.010) than that observed in the overlap paradigm (22 vs. 3%). In other words, there is an almost distinct population of express latencies in the gap paradigm for saccades made at close viewing distance.
Pure vergence
Figure 4 shows the distribution of latencies for convergence (A) and divergence (B) for both gap and overlap paradigms. The mean latencies are shorter for both convergence and divergence in the gap than those in the overlap paradigm: 330±107 ms for convergence and 242±99 ms for divergence in the gap paradigm, and 350±77 ms for convergence and 362±84 ms for divergence in the overlap paradigm. The gap with respect to the overlap paradigm decreases the latency by 20 ms and by 120 ms respectively for convergence and divergence; this decrease is only statistically significant for divergence (χ2 (15,1)=8.06, P=0.004).

Latency distribution in the gap (solid line) and overlap (dotted line) paradigms for convergence (A) and divergence (B). Other notations as in Fig. 3
The percentage of express movements is low for convergence (4 and 3% in the gap and overlap paradigms, respectively) while it is significantly higher (χ2 (15,1)=4.55, P=0.034) for divergence in the gap (14%) with respect to the overlap (1%) paradigms.
Combined movements
The latency distribution for combined movements is shown in Fig. 5. The mean latencies of the saccade components of the movements combined with convergence and divergence are 299±77 ms and 252±66 ms respectively in the gap paradigm and 375±110 ms and 386±107 ms in the overlap paradigm. The latency is shorter in the gap than in the overlap paradigm by 76 ms and 134 ms respectively for saccades combined with convergence and divergence; such differences are statistically different (χ2 (15,1)=5.40, P=0.050 and χ2 (15,1)=10.28, P=0.001, for saccades combined with convergence and divergence, respectively).

Latency distribution in the gap (solid line) and overlap (dotted line) paradigms for the saccade components of movements combined with convergence (A) and with divergence (B), and for the convergence (C) and divergence (D) components of combined movement. Other notations as in Fig. 3
For saccades combined with convergence, the percentage of express movements is low: 2% in both gap and overlap paradigms. In contrast, for the saccade components of movements combined with divergence in the gap paradigm there is a high rate of express movements (12% versus only 3% in the overlap paradigm); such a difference is statistically significant (χ2 (15,1)=5.33, P=0.021).
The latency distributions of the convergence and divergence components of combined movements are shown in Fig. 5c,d. The group mean latencies in the gap paradigm are 312±74 ms and 259±75 ms respectively for the convergence and divergence components, and 378±106 ms and 394±107 ms respectively in the overlap paradigm. The gap paradigm decreases the latency of the convergence component by 66 ms and that of the divergence component by 135 ms; this decrease is only statistically significant for the divergent component (χ2 (15,1)=10.28, P=0.001).
As for the saccade component, the percentage of express movements is low for the convergence component, and equal in the two paradigms (2%); in contrast, for the divergence component, the percentage of express movements tends to be significantly higher (χ2 (15,1)=4.87, P=0.05) in the gap (10%) than in the overlap (only 1%) paradigms.
Frequency of anticipatory movements
Table 1 shows the percentage of anticipatory movements (with latencies <80 ms) observed in all children tested. For saccades at far and close distances, the percentage of anticipatory movements is similar in the gap and in the overlap paradigms (4–6%). In contrast, vergence movements—particularly divergence—show a higher percentage of anticipatory movements in the gap paradigm than in the overlap paradigm; the difference between the two paradigms is only significant for divergence (χ2 (15,1)=4.50, P=0.034).
The percentage of anticipatory movements does not change significantly for either component of the combined movement, in both the gap and the overlap paradigms.
Discussion
The main findings of this study of fifteen children are as follows: (1) the gap paradigm reduced the mean latencies of saccades and vergence eye movements in both their pure and combined form; however, such decreases reached significance only for saccades at close distance, for divergence (pure and combined), and for saccades combined with vergence; (2) the gap paradigm significantly increased the frequency of express movements for saccades at close viewing distance, for divergence, and for saccades combined with divergence; in contrast, the frequency of express movements for saccades at far viewing distance and for convergence (pure or combined) was similar in the gap and in the overlap paradigm; (3) the gap paradigm significantly increased the rate of anticipatory divergence movements. Next we will discuss these findings.
Pure saccades
First, this study showed that, independent of the paradigm used (gap/overlap), the latencies of saccades made at far viewing distance are longer than those of saccades made at close viewing distance. Such a distance-dependency for saccade latency confirms the previous observation in normal children and adults found by Yang et al (2002) using the same spatial arrangement as ours but a different temporal arrangement (simultaneous paradigm). As suggested by Yang and collaborators, the shorter latencies for saccades at close viewing distances could be due to sensory (higher luminosity of the close LEDs), motor or attention factors. For instance, disengagement of oculomotor fixation when the eyes are converging to fixate a target LED at a close distance could be facilitated. Similarly, when the eyes are converging, the disengagement of visual attention from this point may be faster (see Yang et al 2002). Indeed, psychophysical studies (such as McCourt and Garlinghouse 2000) show that visuo-spatial attention can be modulated by the viewing distance; furthermore Maringelli et al (2001) suggest the presence of distinct attentional systems that control far and close visual space. In line with these observations, we can assume that visuo-spatial attention can be oriented more easily in close than in far space. However, to better understand how this mechanism works, further psychophysical studies testing visual attention capabilities at far and close space are needed.
The second important finding is that the gap paradigm reduced the mean latencies of saccades at both far and close viewing distances, but a significant increase in express saccades occurred only for saccades at close viewing distance. This result is, to our knowledge, new; indeed, previous studies (Munoz et al 1998; Klein and Foerster 2001) comparing the influence of gap/overlap effects on saccade latency in children have only used at one, intermediate viewing distance (100 and 60 cm, respectively). Our data suggest that saccades with shorter latencies (such as saccades at close viewing distance in this study) are more likely to become express saccades than saccades with longer latencies (such as saccades at far viewing distance).
Pure vergence
Shorter latencies for divergence than for convergence movements have also been observed in children of comparable age by Yang et al (2002). The new important finding is that the gap paradigm has a different effect for convergence than for divergence. Indeed, the decrease of the mean latency by the gap paradigm was larger for divergence (120 ms) than for convergence (only 20 ms); the gap paradigm increased the rate of express latency for divergence. This observation is compatible with the studies of Takagi et al (1995) and Coubard et al (2004) showing that, even in adults, the gap paradigm can produce movements with express latencies, but only for divergence, not for convergence.
Furthermore, it is important to point out that, even if the ages of the subjects examined differed between our study and the above-cited study (seven-year-old children in the present study, and adults in Coubard’s study), the percentage of express divergence in the gap paradigm was the same (14%) in both studies. This means that the frequency of express vergence does not depend on the age of the subject. Recall that several studies (Munoz et al 1998; Klein and Foerster 2001) have shown that the percentage of express saccades is higher in children than in adults. This finding suggests that there are different mechanisms for triggering express saccades and express divergence: the latter is a stable mechanism which is not age-dependent.
Combined movements
The saccade and vergence components of combined movements have latencies longer than the corresponding pure movements; this is the case for both gap/overlap paradigms. Longer latencies have been found for saccades combined with convergence.
This result is in line with our previous findings (Yang et al 2002) in children of a similar age for combined movements stimulated with the synchronous paradigm. The present study expands this observation and confirms the existence of differences between pure and combined movements, regardless of the fixation task. The data are also compatible with the idea that combined movements are produced by co-activation of the two sub-systems, saccade and vergence (Zee et al 1992; Collewijn et al 1995; Takagi et al 1995).
The effect of the gap paradigm with respect to the overlap paradigm was similar to that found for pure movements: the gap reduced the mean latency for the saccade and vergence components of combined movements. Moreover, similar to pure movements, the increased number of express latencies caused by the gap paradigm has only been observed for saccades combined with divergence, which have shorter latencies than the saccades combined with convergence.
The few studies dealing with the influence of the gap effect on combined movements (Takagi et al 1995; Coubard et al 2004) examined adults, and they reported (as in our study) decreased mean latencies for both of the components of combined movements. Moreover, in support of our findings on children, Coubard et al found that express saccades mainly occurred for saccades combined with divergence in adults; saccades combined with convergence express movements were rare.
Anticipatory movements
The type of fixation task has no influence on the percentage of anticipatory saccades (pure or combined) and of anticipatory convergence; this finding expands upon previous observations of pure saccades by Ross et al (1994) and Klein and Foerster (2001). Frequent anticipatory saccades in young children (<10 years old) have been attributed to limitations in their capacity to inhibit unwanted movements, which is not completely developed in children; such capacity matures slowly, at the age of 10–13 years. This could be true for both pure and combined saccades, and for convergence.
In contrast, for divergence, the gap paradigm significantly increased the rate of anticipatory movements with respect to the overlap paradigm. Moreover, it should be noted that this rate is not age-dependent; indeed, the same percentage of anticipatory divergence (16%) observed in children has also been found by Coubard et al (2004) in adults. We suggest that anticipatory or express divergence could be related to predictive control. This finding is in line with the observations of Dorris et al (1997), made on examining the effect of the gap paradigm in monkeys. These authors showed that during the gap period—before the appearance of the target—saccade-related neuron activity in the superior colliculus increased, suggesting that short latency is due to a predictive oculomotor program. As already mentioned in the “Introduction”, the controversy regarding the origin of express movements has not yet been resolved; however, our findings—that express divergence and anticipatory divergence are present and stable with age—are in line with the idea of predictive control. Thus, electrophysiological studies of express divergence in monkeys may be a useful route to obtaining more insight into this mechanism.
Mechanisms
The present study shows two distinct effects of the gap paradigm on the latency of eye movements. The first is a global effect—a decrease of the mean latency; such an effect occurs for all types of eye movements (saccades as well as vergence). This effect can be attributed to disengaging the fixation; if this occurs before the stimulus onset it could cause a decrease in the fixation activity, therefore accelerating movement initiation (see Findlay and Walker 1999).
Another more specific effect of the gap paradigm is the occurrence of express latencies. In this study, we shown for the first time tha this effect only occurs in children for eye movements that have naturally shorter latency—divergence and saccades at close distance. It is not known whether divergence and saccades made at close viewing distance activate different circuits at the cortical level than those activated by saccades made at far viewing distance and convergence. According to Pierrot-Deseilligny et al (1995), the longer the latency of a movement, the more volitional the movement is believed to be; consequently, we suggest that eye movement initiation can be more or less volitional, depending on the location of the target in space relative to the fixation point. Divergence and saccades at close viewing distance would be initiated in more reflexive manner, perhaps by activating a different cortical circuit; evidence for differential cortical activation related to target location in space has been presented recently by Tzelepi et al (2004).
Finally, one should recall that the convergence condition studied here requires a great deal of effort in terms of vergence and accommodation. The relevance of this condition to everyday life was mentioned earlier (see “Methods” section). Nevertheless, further vergence studies are needed on children viewing more remote distances to examine the difference between convergence and divergence, independent of accommodation effort.
References
Biscaldi M, Fischer B, Stuhr V (1996) Human express saccade makers are impaired at suppressing visually evoked saccades. J Neurophysiol 76:199–214
Bucci MP, Kapoula Z, Yang Q, Wiener-Vacher S, Brémond-Gignac D (2003) Eye movements in 3D space in a young subject with Congenital Central Hypoventilation Syndrome (CCHS). Strabismus II 2:95–107
Collewijn H, Erkelens CJ, Steinman RM (1995) Voluntary binocular gaze-shifts in the plane of regard: dynamics of version and vergence. Vision Res 35:3335–3358
Coubard O, Daunys G, Kapoula Z (2004) Gap effects and vergence latency. Exp Brain Res 154:368–381
Dorris MC, Pare M, Munoz DP (1997) Neuronal activity in monkey superior colliculus related to the initiation of saccadic eye movements. J Neurosci 17:8566–8579
Evans BJW (1997) Pickwell’s binocular vision anomalies: investigation and treatement. Butterworth-Heinemann, Oxford
Findlay JM, Walker R (1999) A model of saccade generation based on parallel processing and competitive inhibition. Behav Brain Sci 22:661–674; discussion 674–721
Fischer B, Boch R (1983) Saccadic eye movements after extremely short reaction times in the monkey. Brain Res 260:21–26
Fischer B, Ramsperger E (1984) Human express saccades: extremely short reaction times of goal directed eye movements. Exp Brain Res 57:191–195
Fischer B, Weber H (1993) Express saccades and visual attention. Behav Brain Sci 16:415–424
Fischer B, Biscaldi M, Gezeck S (1997) On the development of voluntary and reflexive components in human saccade generation. Brain Res 754:285–297
Fukushima J, Hatta T, Fukushima K (2000) Development of voluntary control of saccadic eye movements I: age-related changes in normal children. Brain Dev 22:173–180
Kingstone A, Klein RM (1993) Visual offsets facilitate saccadic latency: does predisengagement of visuospatial attention mediate this gap effect? J Exp Psychol Hum Percept Perform 19:1251–1265
Klein C, Foerster F (2001) Development of prosaccade and antisaccade task performance in participants aged 6 to 26 years. Psychophysiology 38:179–189
Leigh RJ, Zee DS (1999) The neurology of eye movement. Oxford University Press, New York
Luna B, Thulborn KR, Munoz DP, Merriam EP, Garver KE, Minshew NJ, Keshavan MS, Genovese CR, Eddy WF, Sweeney JA (2001) Maturation of widely distributed brain function subserves cognitive development. Neuroimage 13:786–793
Maringelli F, McCarthy J, Steed A, Slater M, Umilta C (2001) Shifting visuo-spatial attention in a virtual three-dimensional space. Brain Res Cogn Brain Res 10(3):317–322
McCourt ME, Garlinghouse M (2000) Asymmetries of visuospatial attention are modulated by viewing distance and visual field elevation: pseudoneglect in peripersonal and extrapersonal space. Cortex 36(5):715–731
Munoz DP, Wurtz RH (1993) Fixation cells in monkey superior colliculus. II. Reversible activation and deactivation. J Neurophysiol 70(2):576-589
Munoz DP, Broughton JR, Goldring JE, Armstrong IT (1998) Age-related performance of human subjects on saccadic eye movement tasks. Exp Brain Res 121:391–400
von Noorden GK, Campos EC (2002) Binocular vision and ocular motility: theory and management of strabismus. Mosby, St. Louis, MO
Pare M, Munoz DP (1996) Saccadic reaction time in the monkey: advanced preparation of oculomotor programs is primarily responsible for express saccade occurrence. J Neurophysiol 76:3666–3681
Pierrot-Deseilligny C, Rivaud S, Gaymard B, Muri R, Vermersch AI (1995) Cortical control of saccades. Ann Neurol 37:557–567
Pierrot-Deseilligny C, Ploner CJ, Muri RM, Gaymard B, Rivaud-Pechoux S (2002) Effects of cortical lesions on saccadic eye movements in humans. Ann N Y Acad Sci 956:216-29
Reuter-Lorenz PA, Hughes HC, Fendrich R (1991) The reduction of saccadic latency by prior offset of the fixation point: an analysis of the gap effect. Percept Psychophys 49:167–175
Ross RG, Radant AD, Young DA, Hommer DW (1994) Saccadic eye movements in normal children from 8 to 15 years of age: a developmental study of visuospatial attention. J Autism Dev Disord 24:413–431
Saslow MG (1967) Effects of components of displacement-step stimuli upon latency for saccadic eye movements. J Optic Soc Am 57:1024–1029
Takagi M, Frohman EM, Zee DS (1995) Gap-overlap effects on latencies of saccades, vergence and combined vergence-saccades in humans. Vision Res 35:3373–3388
Tam WJ, Ono H (1994) Fixation disengagement and eye-movement latency. Perception Psychophys 56(3):251–260
Tzelepi A, Lutz A, Kapoula Z (2004) EEG activity related to preparation and suppression of eye movements in 3-D space. Exp Brain Res 155(4):439–449
Yang Q, Bucci MP, Kapoula Z (2002) The latency of saccades, vergence, and combined eye movements in children and in adults. Invest Ophthalmol Vis Sci 43:2939–2949
Ygge J, Lennerstrand G, Rydberg A, Wijecoon S, Petterson B (1993) Oculomotor functions in a Swedish population of dyslexic and normally reading children. Acta Ophthalmol 71:10–21
Zee DS, Fitzgibbon EJ, Optican LM (1992) Saccade-vergence interactions in humans. J Neurophysiol 68:1624–1641
Acknowledgements
The work presented here was part of the Maîtrise diploma “Cellular biology and physiology”, run at René Descartes University (Paris V) of N. Pouvreau. The authors thank the director of the school where the experiment took place, the teacher, and the children of the class who participated in the experiment. The authors are particularly grateful to the optometrist H. Puech for conducting the clinical examinations.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Bucci, M.P., Pouvreau, N., Yang, Q. et al. Influence of gap and overlap paradigms on saccade latencies and vergence eye movements in seven-year-old children. Exp Brain Res 164, 48–57 (2005). https://doi.org/10.1007/s00221-005-2214-0
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00221-005-2214-0