
Abstract
A force-matching task was used to study the influence of constraining tactile information from the hand on the perception of forces generated with the index finger flexors, the palmar prehensile grasp and with the elbow flexors. Subjects generated the same reference forces (2–10 N) with each muscle group and matched these using the corresponding muscle group in the other arm. Force perception was studied under normal conditions and when the tactile feedback from the fingertips was constrained through the use of rigid finger splints. There was no difference between the three muscle groups when matching forces under normal conditions, but when spatial tactile information from the fingertips was attenuated forces were underestimated in perceived magnitude as compared to control conditions. These results suggest that the perception of force is influenced by tactile cues that convey information about the contact surface and that distributed spatial force cues are normally used in the perception of forces generated by the hand.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Many factors have been shown to influence the perceived magnitude of muscular forces including fatigue, cutaneous anesthesia, the contractile state of the muscle, and the muscle group generating the force (Gandevia and Kilbreath 1990; Mai et al. 1991). Under normal conditions, the average error in matching the forces produced by two muscle groups is around 3.5%, when forces are expressed in terms of the maximum voluntary contraction (MVC) of each muscle group (Jones 1989). The accuracy with which forces are matched (as defined by the coefficient of variation) varies across different muscle groups, although the variation in accuracy does not appear to be associated with a proximal to distal gradient. The most accurate performance has been reported for the flexor pollicis longus, a muscle controlling flexion of the distal joint of the thumb, which is significantly more accurate than other hand muscles (Kilbreath and Gandevia 1993).
Sensory inputs from cutaneous mechanoreceptors influence the perceived magnitude of forces generated by individual fingers (Gandevia and McCloskey 1977; Henningsen et al. 1995) and the hand (Flanagan and Bandomir 2000). When the skin on the thumb is anesthetized, the perceived heaviness of weights lifted by flexing the thumb increases by over 40%, whereas for the index finger skin anesthesia results in only a 13% increase in perceived heaviness (Kilbreath et al. 1997). The effect of skin anesthesia on force and weight perception is thought to reflect the loss of the net facilitatory effect of cutaneous afferents from the thumb and index finger on the descending motor command. The greater change in perceived heaviness with thumb anesthesia is consistent with a critical role for cutaneous mechanoreceptors in controlling forces generated by the thumb. These findings together with many others on the perception of force indicate that judgments of the heaviness of weights and of the magnitude of muscle forces depend primarily on the centrally generated motor command and not just on peripheral sensory signals arising from muscles (Gandevia and Klibreath 1990; Jones 1986).
The objective of the present experiment was to evaluate the contribution of tactile afferent information from the hand to the perception of forces produced by different muscle groups in the arm. Forces were generated by three muscle groups: the elbow flexors, the forearm and hand muscles involved in the palmar prehensile grasp, and the index finger flexors. Subjects were required to match the reference force produced by one muscle group by producing a perceptually equivalent force with the same muscle group in the contralateral arm under normal conditions and when tactile cues from the grasping surface were constrained.
Materials and methods
Participants
Nine normal healthy adult subjects (three women and six men) participated in this experiment. They had no known abnormalities of the neuromuscular or peripheral sensory systems. They ranged in age from 20 to 37 years (mean 26 years). The experiments were approved by the local ethics committee.
Apparatus
The experimental apparatus has been described in detail (Jones 2003). Elbow flexion forces and palmar pinch forces were measured using shear beam load cell (Omega model LCC) force transducers. The outputs of the force transducers were processed by signal conditioning amplifiers prior to being sampled by an Agilent Data Acquisition Unit (Model 34970A) that was connected to a Dell (XPS) computer. The output from the signal conditioning amplifier was also fed to an oscilloscope (Tektronix TDS 3012B) mounted on the rig in front of the subject. This displayed the reference forces being produced by the subject when required. Index finger flexion forces were measured using force gauges (Shimpo model DFS-20). The distal phalanx of each index finger rested on a 25-mm diameter disk that was screwed onto the shaft of the force gauge. The RS232 output of the force gauge was connected to the Data Acquisition Unit and the oscilloscope mounted in front of the subject.
Elbow, palmar pinch or finger forces were sampled at 10 Hz during each trial. A D/A converter was used to control the amplitude of the target force presented on the oscilloscope in front of the subject. The operating system sound function was used to present auditory signals to the subject, which indicated the start of each trial and the matching interval.
Procedure
Subjects sat in the experimental rig with their arm and hand in the appropriate position for the particular set of trials being conducted. At the beginning of each block of trials subjects were told which muscle group was to be used. Subjects grasped a cylindrical rod in their left and right hands to generate elbow flexion forces. The rods projected from motors on which the force transducers were mounted, and they were pulled in the sagittal plane to generate force. This did not result in any perceptible movement of the motor or rod. Although the normal force was primarily generated by the biceps brachii muscles that flex the elbow, the grip force produced by the hand as it grasped the handle was also a component of force production. To produce forces using the palmar pinch, subjects grasped the force transducers between the pads of the thumb and index and middle fingers. The palmar grasp involves flexion of the metacarpophalangeal (MCP) and of the proximal and distal interphalangeal joints of the index and middle fingers, and adduction and flexion of the MCP joint of the thumb. The forces produced in the palmar grasp therefore involve the extrinsic flexor muscles, the interosseous and lumbrical muscles, and the thenar muscles. Index finger forces were produced by flexing the index finger at the MCP joint, again using the interosseous and lumbrical muscles in the hand; the MCP joint maintained an angle of 170–180°. Forces were matched using corresponding muscle groups (left and right elbow flexors) under normal conditions and with the tactile sensory feedback from the right, reference hand constrained.
Sensory feedback from the hand was modified by eliminating spatial tactile feedback from the grasping surface. In the finger and grasp matching conditions, plastic finger splints (Smith and Nephew Rolyan Inc., USA) completely covered the ventral surfaces of the digits producing the reference forces. The splints were rigid (3 mm thick) and concave in shape to mold to the surface of the finger pad. They effectively eliminated any spatial force cues from the finger pads. When matching forces by flexing the forearm, subjects wore a fabric glove which had a rigid surface embedded in the distal part of each finger sleeve. This permitted subjects to flex the fingers so that they could grasp the rod, but again eliminated most spatial force cues from the hand.
Subjects were required to produce a reference force of 2, 5, 8, or 10 N, which was presented randomly within each block of trials. There were five repetitions of each force, giving a total of 20 trials per matching condition, and for each of the three muscle groups forces were matched under normal and modified sensory conditions. At the beginning of each trial the target force was displayed on the oscilloscope together with the actual force being produced by the reference limb. Subjects had to produce the reference force within 4 s using this visual feedback and were then required to match the perceived amplitude of the reference force by generating a similar force with the matching limb without visual feedback of the matching force. The instructions given to subjects were to make the forces produced by the two muscle groups the same. They were given 3 s to match the forces, followed by a 3-s rest period. A sequence of tones indicated to subjects the start of each trial and the time to match the forces. Each trial lasted 10 s and there was a 3–4 min break between each of the five experimental conditions, which were presented in a random order. Subjects were given at least ten practice trials before the experiment to familiarize them with the procedure.
Results
The reference forces were readily attained by subjects on each trial and on average were within 0.5 N of the target force. The mean reference and matching forces produced by subjects in the final 1 s of the matching interval were calculated for each subject. An initial analysis was performed of the matching forces produced under normal conditions in order to see whether there was a difference between the three muscle groups with respect to the accuracy with which forces were matched. A repeated-measures analysis of variance (anova) performed on these data indicated that there was a significant effect of force amplitude [F(2, 16)=131.26, p<0.001] but no effect of muscle group. The interaction between force and muscle group was also not significant. This finding shows that the resolution with which the forces are perceived is similar across the three muscle groups.
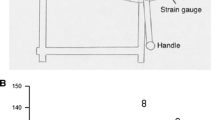
One of the objectives of this experiment was to examine how tactile sensory feedback from the grasping surface in the hand influenced the perceived magnitude of forces produced by different muscle groups. The matching forces produced under normal conditions and when the sensory feedback from the reference limb was constrained are shown for each muscle group in Fig. 1. It appears that when tactile sensory feedback from the hand is constrained, forces produced by muscles controlling movements of the hand and arm are underestimated in magnitude as compared to the normal feedback condition. A repeated measures anova was conducted on these matching forces with muscle group, sensory feedback condition, and force as within-subjects factors. The results showed that there was a main effect of sensory feedback condition [F(1, 8)=22.37, p=0.001], and force [F(3 24)=139.03, p<0.001], but not of muscle group. The interaction between sensory condition and force [F(3, 24)=4.78, p=0.009] was also significant. Post hoc tests revealed that the matching forces were significantly smaller when the sensory feedback from the hand was constrained and that the effect of altering sensory feedback was greater at larger forces. The relation between the reference and matching forces was compared in the two sensory feedback conditions. The correlation coefficient was essentially the same (r=0.98 normal, r=0.99 reduced feedback), and the slope of the relation was slightly lower in the constrained sensory feedback condition (0.77 as compared to 0.82).

Group mean matching forces as a function of reference force. Forces produced under normal conditions (filled) and when sensory feedback from the hand is constrained (unfilled) by the index finger flexors (triangles), palmar pinch (circles), and elbow flexor (squares) muscles
The precision with which the forces were matched was evaluated in terms of the constant and absolute errors. Constant errors were calculated by subtracting the matching force from the reference force on each trial. The group mean constant forces were relatively small and ranged from −1.05 to 2.12 N. The absolute errors were also small and ranged from 0.36 to 2.8 N across the various conditions, with an overall mean value of 1.6 N. An analysis of these errors indicated a main effect of force [F(3, 24)=11.47, p=0.007] and of sensory feedback condition [F(1, 8)=8.69, p=0.02], but not of muscle group. The interactions were not significant. As muscle group was not a significant factor in these analyses, the errors have been collapsed across muscle group in Fig. 2 to demonstrate the effect of constraining tactile sensory feedback on matching accuracy.

Group mean absolute errors when matching forces under normal conditions (striped bars) and when sensory feedback from the hand is constrained (white bars)
The accuracy with which the forces were matched was also evaluated in terms of the coefficient of variation (CV) associated with each experimental condition. With the exception of the 2-N condition the CVs were relatively consistent across the various experimental conditions (mean 17%, range: 10–36%). The larger variation associated with matching 2-N forces is not unexpected as any fluctuation in force represents a significant percentage of the mean value. The group mean coefficient of variation without the 2-N condition is 13%. An analysis of these coefficients of variation indicated that there was a significant main effect of force [F(3, 24)=59.11, p<0.001], that reflected the larger coefficients associated with the 2-N force, but no effect of muscle group or sensory condition. None of the interactions were significant.
Discussion
The results from the present experiment indicate that with respect to both the control and perception of force there is no significant difference between the muscles controlling movements of the fingers and forearm. The lack of any distal to proximal gradient in sensitivity was found when the muscles were required to produce the same forces, which were within the range that the hand typically uses for skilled tasks such as grasping small objects or typing on a keyboard (Johansson and Westling 1987). These findings on the perception of force are consistent with those on force discrimination, which show that the threshold remains relatively constant at 7% across a broad range of muscle groups and forces (Jones 1986).
This experiment also showed that the perceived magnitude of forces generated by muscles controlling the hand and arm are influenced by cutaneous feedback from the grasping or contact surface, even when those forces are produced by muscles in the upper arm. For each of the three muscle groups studied removing feedback about the spatial distribution of forces on the hand resulted in a change in the perceived magnitude of forces generated. However, in contrast to the effects of cutaneous anesthesia which results in an increase in perceived force or heaviness (Kilbreath et al. 1997) and a decrease in the maximum forces that the fingers can produce (Augurelle et al. 2003), in the present experiment constraining the sensory input from the hand caused subjects to underestimate the magnitude of forces. It appears that when tactile feedback from the finger pad is available but attenuated, it affects the perception of force differently from when all sensory feedback from the finger is eliminated. In the latter situation, there is a loss of the cutaneous reflexes that are facilitatory to homonymous motoneurons (Kilbreath et al. 1997), whereas these reflexes are still functional when sensory feedback is constrained.
The underestimation of force was not accompanied by a change in the consistency with which forces were perceived, as the coefficients of variation did not change when the tactile cues were constrained. It would appear that in the absence of feedback about the spatial distribution of forces on the fingers, muscle forces are perceived to be smaller than they actually are. Information about the spatial distribution of forces on the finger pads comes from slowly adapting cutaneous mechanoreceptors (SA1) that signal the local stress-strain field on the skin and are particularly sensitive to skin indentation (Westling and Johansson 1984). The present results suggest that these cutaneous signals contribute to the perception of muscle force and that when they are diminished, forces are perceived to be smaller.
It is known that the surface features of objects such as their texture (Flanagan et al. 1995) or surface curvature (Goodwin and Wheat 1992) influence their perceived heaviness or contact force. Henningsen et al. (1995) found that the shape of the contact pad against which the index finger generated forces influenced force perception with forces being over-estimated when they were generated against a conical as compared to a flat surface. These studies, together with the present findings, are consistent in demonstrating that cutaneous feedback from the contact surface of an object influences the perception of force used to support the object. These interactions between cutaneous signals and central motor commands occur even when the forces are not required to support the object against gravity, as in the present experiment.
References
Augurelle AS, Smith AM, Lejeune T, Thonnard JL (2003) Importance of cutaneous feedback in maintaining a secure grip during manipulation of hand-held objects. J Neurophys 89:665–671
Flanagan JR, Wing AM, Allison S, Spenceley A (1995) Effects of surface texture on weight perception when lifting objects with a precision grip. Percept Psychophys 57:282–290
Flanagan JR, Bandomir CA (2000) Coming to grips with weight perception: effects of grasp configuration on perceived heaviness. Percept Psychophys 62:1204–1219
Gandevia SC, Kilbreath SL (1990) Accuracy of weight estimation for weights lifted by proximal and distal muscles of the human upper limb. J Physiol 423:299–310
Gandevia SC, McCloskey DI (1977) Changes in motor commands, as shown by changes in perceived heaviness, during partial curarization and peripheral anaesthesia in man. J Physiol 272:673–689
Goodwin AW, Wheat HE (1992) Magnitude estimation of contact force when objects with different shapes are applied passively to the fingerpad. Somatosens Mot Res 9:339–344
Henningsen H, Ende-Henningsen B, Gordon AM (1995) Contribution of tactile afferent information to the control of isometric finger forces. Exp Brain Res 105:312–217
Johansson RS, Westling G (1987) Signals in tactile afferents from the fingers eliciting adaptive motor responses during precision grip. Exp Brain Res 66:141–154
Jones LA (1986) Perception of force and weight: theory and research. Psychol Bull 100:29–42
Jones LA (1989) Matching forces: constant errors and differential thresholds. Perception 18:681–687
Jones LA (2003) Perceptual constancy and the perceived magnitude of muscle forces. Exp Brain Res 151:197–203
Kilbreath SL, Gandevia SC (1993) Neural and biomechanical specialization of human thumb muscles revealed by matching weights and grasping objects. J Physiol 472:537–556
Kilbreath SL, Refshauge K, Gandevia SC (1997) Differential control of the digits of the human hand: evidence from digital anaesthesia and weight matching. Exp Brain Res 117:507–511
Mai N, Schreiber P, Hermsdörfer J (1991) Changes in perceived finger force produced by muscular contractions under isometric and anisometric conditions. Exp Brain Res 84:453–460
Westling G, Johansson RS (1984) Factors influencing the force control during precision grip. Exp Brain Res 53:277–284
Acknowledgments
This research was supported by grant NS-40836 from the National Institutes of Health.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Jones, L.A., Piateski, E. Contribution of tactile feedback from the hand to the perception of force. Exp Brain Res 168, 298–302 (2006). https://doi.org/10.1007/s00221-005-0259-8
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00221-005-0259-8