
Abstract
Using the in vitro isolated whole brain preparation of the guinea pig, we tested the synaptic effects induced by the stimulation of pontine nuclei (PN) in intracellularly recorded and stained principal cells of the cochlear nucleus (CN). Twenty percent of the recorded cells in all CN subdivisions responded to stimulation of either ipsilateral or contralateral PN, and 12% of the cells exhibited convergence of inputs from both sides. The responses were recorded only in stellate cells of the ventral CN and in the pyramidal cells of the dorsal CN, whereas no responses were observed in bushy, octopus, and giant cells. PN stimulation produced excitatory and inhibitory postsynaptic potentials as well as mixed responses. The heterogeneous nature and the wide latency range (3.2–18 ms) of observed responses suggest significant variability in the underlying synaptic mechanisms and the implicated pathways. We propose that PN projections to the CN, terminating mainly in the granule cell domain (GCD), together with other non-auditory and auditory inputs contribute to multimodal convergence in the GCD leading ultimately to modulatory actions on the output activity of CN principal cells.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
In addition to the major auditory input from the ipsilateral auditory nerve and inputs from other auditory structures (e.g., contralateral cochlear nucleus, superior olive, inferior colliculi, auditory cortex), the cochlear nucleus (CN) appears to receive a number of non-auditory projections. Among those are inputs from the dorsal column nuclei (Itoh et al. 1987; Weinberg and Rustioni 1987; Wright and Ryugo 1996), spinal trigeminal nucleus (Itoh et al. 1987; Zhou and Shore 2004), trigeminal nerve (Shore et al. 2000), vestibular afferents (Burian and Gstoettner 1988; Kevetter and Perachio 1989), as well as projections from some other brainstem nuclei (Ryugo et al. 2003). Despite the solid morphological evidence for the existence of various non-auditory inputs to the CN, their physiological influences were so far studied only for somatosensory inputs (Saadé et al. 1989; Young et al. 1995; Kanold and Young 2001; Shore et al. 2003) that are thought to be involved in the coordination of pinna orientation and sound sources as well as in the modulation of sound perceptions by somatic information from the head and neck. A large proportion of non-auditory and some descending auditory projections to the CN terminate in the granule cell domain (GCD), a thin layer of small granule cells overlying most of the CN and separating the ventral cochlear nucleus (VCN) from the dorsal cochlear nucleus (DCN). Thus, the GCD, which is considered as a shell of the CN (see for review Ryugo et al. 2003), is well positioned to integrate a variety of non-auditory information and feedback auditory inputs to modulate the output activity of neurons in the CN core (VCN and DCN). A recent neuroanatomical study has demonstrated the existence of prominent bilateral projections from pontine nuclei (PN) to the CN, also terminating principally in the GCD of the CN (Ohlrogge et al. 2001). The aim of the present work was to assess the physiological properties of PN inputs to the CN cells. Using the in vitro isolated whole brain (IWB) preparation of the guinea pig, we intracellularly recorded from the CN neurons and tested their responses to electrical stimulation of PN on both sides. In addition, recorded CN neurons were labeled with neurobiotin in order to identify cells from which recordings were made. However, as it is very difficult, if not impossible, to record intracellularly from very small granule cells, the modulatory action of PN stimulation could not be studied in the primary target of PN projections terminating in the GCD, but was rather assessed in principal CN cells. An indirect action of PN inputs on principal CN cells through GCD can be hypothesized, since it is well established that, for example, in the cerebellar-like circuit of the DCN superficial layers, granule cell axons form excitatory synapses on the dendrites of principal cells and also contact inhibitory interneurons that project to the principal cells (Mugnaini et al. 1980a, b; Doucet and Ryugo 1997; see also for review Oertel and Young 2004). A preliminary account of this work was presented in an abstract form (Babalian and Rouiller 2004).
Materials and methods
The experiments were performed on pigmented guinea pigs of both genders (body weight 200–500 g) in accordance with animal care rules of Swiss Veterinary Authorities. The methods used to prepare and maintain the IWB preparation in vitro were similar to those described in detail previously (Babalian et al. 1997). Briefly, the guinea pigs were anesthetized with a lethal dose of pentobarbital (150–300 mg/kg, i.p.) and perfused through the heart with cold (10–13°C) Ringer’s solution for 5–6 min. Following decapitation, the brain was dissected from the skull and transferred to the perfusion/recording chamber. The brain was perfused through a fine metallic cannula inserted in one of the vertebral arteries. The second vertebral artery and the other major arteries cut during the dissection were ligated to minimize the leaks of the perfusion solution from the brain. The perfusion rate was progressively increased from 1.5–2 ml/min to 4.5–5 ml/min as the solution and the brain in the recording chamber were progressively warmed from 13°C to the final temperature of 29°C. The perfusion solution had the following composition (in mM): NaCl 126; NaHCO3 26; MgSO4 1.3; KH2PO4 1.2; KCl 3; CaCl2 2.4; glucose 15. It also contained either 1.5% of dextran (MW 70,000, Macrodex®, Braun Medical AG, Switzerland) or 1–2% of gelatin (MW 30,000, Physiogel®, Braun Medical AG, Switzerland). The solution was continuously bubbled with a mixture of 95% O2 and 5% CO2.
The PN and auditory nerves (AN) were stimulated with single monopolar electrical pulses of 0.2 ms duration delivered through self-made stainless steel bipolar electrodes at a rate of 1 Hz. As the PN are relatively large structures located close to the ventral surface of the pons, the stimulating electrodes could be routinely correctly placed with the brain secured in the perfusion chamber in the ventral side up position. The placement of electrodes in the PN was histologically verified in the initial experiments and could be consistently reproduced using external landmarks. Stimulating electrodes were inserted l mm laterally to the midline on both sides, at a depth of 1 mm from the ventral surface, and 1–1.5 mm caudally to the rostral edge of the pons according to the atlas of the guinea pig brainstem (Voitenko and Marlinskii 1993). In order to minimize the possibility of current spread to the neighboring structures, the upper limit of PN stimulation intensity was set at 0.25 mA. Intracellular recording and iontophoretical labeling of CN neurons were made using micropipettes filled with 1–2% solution of tracer neurobiotin in 2 M K-acetate. Conventional electrophysiological equipment was used for the recording and storage of signals. Labeled cells were visualized on frontal brain sections (100 μm) using standard ABC histochemistry with some modifications (Wan et al. 1992).
Results
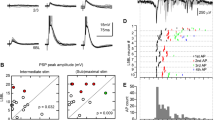
Only cells in good physiological condition (stable membrane potential of at least −55 mV) were retained for physiological testing and subsequent labeling. The effects of PN stimulation were tested in 42 principal CN cells recorded in 27 experiments (isolated brains). The cells were distributed in all the three CN subdivisions: anteroventral (AVCN), posteroventral (PVCN), and dorsal (DCN). Among 13 cells tested in the AVCN, 6 were identified as bushy cells, 6 others as stellate cells, and 1 neuron could not be recovered on histological sections. Eighteen recorded cells in the PVCN were classified as stellate cells (10), octopus cells (3), and unidentified cells (5) as they were not recovered and/or poorly labeled. The nine tested DCN neurons were pyramidal (6), giant (2), and stellate (1) cells. Finally, two tested stellate cells were found in the granule cell lamina separating the PVCN and DCN. The distribution of cells in the CN and the synaptic effects produced by the single-pulse stimulation of PN on both sides are summarized in Table 1. Altogether, ten tested cells responded to stimulation of the PN with a comparable number of cells influenced from the ipsilateral (seven cells) and contralateral (eight cells) sides. In five of the ten responding cells, a convergence of influences from the ipsilateral and contralateral PN was observed. The remaining five cells received inputs either from the ipsilateral (two cells) or the contralateral (three cells) PN. The highest proportion of responding cells was in the DCN followed by AVCN and PVCN. It should be noted that PN stimulation effects were observed only in some categories of tested cells. In the VCN and GCD lamina, only stellate cells responded to PN stimulation (six cells in total), whereas bushy and octopus cells were devoid of PN influences. In the DCN, we observed PN-induced responses exclusively in the pyramidal cells (four cells). Figure 1 illustrates the morphological and physiological characteristics of a cell in the AVCN influenced by PN. The neuron had large radiate stellate cell morphology with long dendrites extending through the entire cross-section of the CN and reaching its medial and lateral edges (Fig. 1b, c). The lateral dendrites of the cell reached the granule cell layer covering the dorso-lateral surface of the CN. Although the axon of the neuron could not be traced for a long distance from the soma, it was clearly directed dorso-caudally within the CN. In this cell, the stimulation of the ipsilateral PN induced a long-latency (12.5 ms) inhibitory postsynaptic potential (IPSP), whereas the contralateral PN stimulation produced a mixed response that included a short-latency (3.2 ms) excitatory postsynaptic potential (EPSP) followed by a longer latency (12 ms) IPSP (Fig. 1a).

Physiological and morphological characteristics of a cell in the AVCN responding to the stimulation of pontine nuclei (PN). a From top to bottom: responses of the cell to a single-pulse stimulation of the ipsilateral auditory nerve (ANi), ipsilateral (PNi), and contralateral (PNc) pontine nuclei, respectively. For PNi stimulation, both the average recording (average) and individual superimposed traces (single) are shown. The remaining recordings are superimposed individual traces. The resting membrane potential of the cell was −60 mV. Small spike amplitude was most likely due to intradendritic recording from the cell. b High-magnification reconstruction of the cell. c Position of the cell in the AVCN. Arrows point to dendrites of the neuron that reach the dorso-lateral surface of the CN, where the granule cell layer and other small interneurons are located
IPSPs accounted for 10 out of the total of 15 recorded responses (Table 1). The remaining responses were either EPSPs (three) or mixed synaptic potentials (two). In five cells that exhibited convergence of inputs from both ipsi- and contralateral PN, the observed combinations of responses were: IPSP–IPSP (two cases), EPSP–EPSP (one case), mixed response–EPSP (one case), and IPSP–mixed response (one case). Response latencies were distributed in a wide range from 3.2 to 18 ms (Fig. 2). The responses with shortest latencies were EPSPs, which had latencies in the range of 3.2–3.5 ms, suggesting a disynaptic transmission. The short-latency EPSPs were observed in the cell illustrated in Fig. 1 and a similar radiate cell with widely spreading dendrites and dorso-caudally oriented axon in the AVCN (not shown). The remaining responses had latencies of 5.3 ms and longer, most likely indicating their polysynaptic origin. Interestingly, only one of the two stellate cells recorded in the granule cell lamina responded to PN stimulation and exhibited a long-latency (17 ms) IPSP to stimulation of the contralateral PN.

Latency distribution of synaptic responses produced in CN cells by stimulation of the ipsilateral (PNi) and contralateral (PNc) pontine nuclei. The histogram includes 16 measurements, as for only one of the two mixed responses the latency of both components could be measured
The observed EPSPs were large enough to induce action potentials in the recorded cells (Fig. 1). Although the amplitudes of recorded IPSPs were usually small, ranging from 0.9 to 4.2 mV, they were functionally efficient as indicated by the discharge inhibition of some spontaneously active pyramidal neurons in the DCN. The duration of IPSPs ranged from 15 to 80 ms. The proportion of excitatory and inhibitory responses was similar in the stellate and pyramidal cells and, although some stellate cells tended to respond with shorter latencies than pyramidal cells, no obvious differences were observed between the response characteristics of the two cell types.
Discussion
Our present work demonstrates the existence of functional modulatory inputs from PN to the CN. About 20% of the principal cells in the CN responded to stimulation of either ipsilateral or contralateral PN. In addition, some of these cells (12%) exhibited a convergence of bilateral PN inputs.
The first principal issue related to our data is how specific are the synaptic responses in CN cells to inputs from PN. There are several auditory structures in the vicinity of PN, including periolivary nuclei, ventral nucleus of the lateral lemniscus, as well as fibers of the trapezoid body (TB) and intermediate acoustic stria (IAS), that are known to project to the CN (Winter et al. 1989; Benson and Potashner 1990; Shore et al. 1991). Thus, it is possible to argue that the observed responses could be at least partially due to the spread of stimulating current from PN to neighboring structures projecting to the CN. We think that such a possibility can be safely ruled out for the following reasons. It has been demonstrated that stimulation current intensities of 0.3–0.5 mA are efficient within a distance of 1 mm from the electrode tip (Sasaki et al. 1970; Shinoda et al. 1985), which can be even smaller with bipolar stimulation. In our experiments, the upper limit of PN stimulating current was set at 0.25 mA, suggesting that the effective current spread was restricted to a distance less than 1 mm from the electrodes. However, the distance between stimulating electrodes and the auditory structures projecting to the CN was bigger (at least 1.5 mm according to our estimates), making the possibility of the non-specific current spread very unlikely. Indeed, in some control experiments, we placed electrodes caudally to PN in order to produce a massive stimulation of the superior olivary structures as well as of the TB and IAS bundles. Such stimulation usually produced large IPSPs in almost all the recorded CN cells and, in addition, in many of them, sending their axons through TB or IAS, generated antidromic action potentials. Moreover, the latencies of synaptic responses were significantly shorter (mean value 3.7 ms, range 2.1–6.3 ms; unpublished data) than those found with PN stimulation. This response pattern was very different from the one observed with our standard PN stimulation intensities and could be sometimes seen in our experiments when the intensities were increased to above 0.5 mA. The threshold of responding cells was 50–100 μA, and a gradual increase of stimulation intensity up to the maximal tested value (0.25 mA) never resulted in sudden changes of response latencies and/or nature, which could be an indicative of a current spread to neighboring structures. Since the PN are large structures, it is not even excluded that the limitation of the current strength in the present study could result in failure of stimulating remote parts of the PN and, consequently, in underestimation of the real strength of PN inputs to the CN. There are some fiber tracts running in the vicinity and through the PN (e.g., pyramidal tract, medial lemniscus) that could be activated by stimuli applied to PN. However, no projections from these fiber systems to the CN have been described so far. Our results did not confirm the existence of direct afferent projections from the CN to PN (Kandler and Herbert 1991), as we never observed an antidromic activation of CN cells in response to PN stimulation. However, it is possible that, for some reason (e.g., cell size, location in the CN), we did not penetrate CN cells projecting to PN.
A tracing study in rats (Ohlrogge et al. 2001) has demonstrated that projections from the contralateral PN to the CN outnumbered the ipsilateral projections by about four times. Our data are not in accordance with these observations, as comparable proportions of neurons were influenced by the stimulation of the ipsilateral and contralateral PN. One possible explanation for this discrepancy, though very speculative, could be related to the fact that the anatomical data showed a significant asymmetry of PN projections to the GCD, whereas our recordings were made from principal CN cells. The observation of predominant contralateral PN projection to the GCD should not necessarily result in stronger contralateral PN effects on principal CN cells. It is possible that various combinations of the strength and/or convergence of inputs from the GCD to the CN core might result in more balanced bilateral PN actions on principal CN cells. Other possible explanations for the discrepancy between our data and previous tracing study in rats could include the species differences and/or biased number/percentage of influenced cells due to the limited sample of cells recorded in the present study.
The population of responses produced by PN stimulation in CN principal cells was heterogeneous, both in terms of their nature (EPSPs, IPSPs, mixed responses) and large latency range (3.2–18 ms), suggesting significant variability in underlying synaptic mechanisms and implicated pathways. The responses with shortest latencies of 3.2–3.5 ms were EPSPs recorded in two radiate stellate cells of the AVCN. In the IWB preparation, these latency values most likely correspond to disynaptic transmission (Babalian et al. 1997, 1999, 2002). As both the AVCN cells extended their dendrites to the GCD and the PN input to GCD was shown to be excitatory (Ohlrogge et al. 2001), the most probable mechanism of observed disynaptic EPSPs would be an excitation of the granule cells by PN projections and following excitatory action from granule cells to target cells. The large majority of other responses in the present study were IPSPs at longer latencies (5.3 ms and longer), suggesting the involvement of additional synaptic relays. Cells similar to the two AVCN cells that exhibited disynaptic EPSPs to stimulation of PN could mediate some of the polysynaptic responses. Indeed, the morphology of the two AVCN cells, with large dendrites and dorso-caudally oriented axon, is similar to the characteristics of type II, large radiate stellate cells of D-type, immunoreactive for glycine, with onset responses (see for review Arnott et al. 2004). These cells were demonstrated to have an axon terminating in the DCN and partially in the VCN and are good candidates for providing a wideband inhibition of PVCN and DCN neurons including pyramidal cells (Arnott et al. 2004). Thus, the polysynaptic inhibitory effects of PN stimulation observed in the pyramidal cells and some planar, TB-projecting, stellate cells in the PVCN could be at least partially mediated through the disynaptic excitation of some VCN radiate cells, which in turn inhibit target cells in the DCN and VCN. An additional or alternative mechanism for polysynaptic inhibition of pyramidal cells from PN may function through a cerebellum-like circuit of the DCN (reviewed in Oertel and Young 2004) and would include PN–GCD–carthweel cell–pyramidal cells synaptic pathway. Even if no PN endings were found in the layer 2 of the DCN containing granule cells (Ohlrogge et al. 2001), such a pathway may function through granule cells in all other CN areas known to send parallel fibers to the DCN (Mugnaini et al. 1980b). It should also be noted that additional synaptic relays in the GCD and/or within the magnocellular CN core, including, for example, Golgi cells, unipolar brush cells, interneurons, and CN cells projecting to the contralateral CN, may account for PN-induced responses with very long latencies. Finally, mixed responses observed in the present study would result from convergence of different synaptic pathways on the same target cell. In 16 cells that did not respond to single-pulse stimulation of PN, we applied a stimulation protocol using pulse trains (four pulses at 150 Hz). Train stimulation induced responses in only one of the tested cells, but these responses were not taken into consideration in our database as they could be, at least partially, produced by temporal summation of synaptic responses in polysynaptic circuits other than PN projections to CN.
Our data demonstrate the existence of bilateral modulatory action of PN on CN principal cells. The question arises of the possible functional significance of these inputs in the auditory function. The PN are important relay structures of a major pathway in the central nervous system, the cerebral–cerebellar–cerebral loop (see for reviews Brodal and Bjaalie 1992; Schmahmann and Pandya 1997; Schwarz and Thier 1999). The PN were traditionally considered to be mainly involved in motor control (Allen and Tsukahara 1974). However, although the majority of inputs to PN originate from the motor and sensorimotor cortices, the PN apparently integrate various inputs. The PN receive projections from all parts of the cortex, including the visual (Hollander et al. 1979; Wiesendanger and Wiesendanger 1982a, b; Legg et al. 1989) and auditory cortices (Wiesendanger and Wiesendanger 1982a; Azizi et al. 1985; Knowlton et al. 1993; Kimura et al. 2004). In line with these observations are the findings of PN neurons responding to visual and auditory stimuli (Glickstein et al. 1972; Baker et al. 1976; Aitkin and Boyd 1978), as well as to stimulation of visual and auditory cortices (Potter et al. 1978). Based on these data it has been argued that the major role of the cerebro–ponto–cerebellar pathway is the sensory guidance of movements (Stein and Glickstein 1992), including a contribution to such specialized behaviors as smooth-pursuit eye movements and visually guided saccades. Finally, it has been proposed that the cerebro–ponto–cerebellar loop might have a role in higher brain functions as the prefrontal areas, classically associated with cognitive functions, contribute to the pathway (reviewed in Middleton and Strick 1997; Schmahmann and Pandya 1997). In addition to cortical inputs, the PN receive afferents from subcortical sources, particularly auditory inputs (Aitkin and Boyd 1978; Kandler and Herbert 1991). Hence, the studies summarized above indicate that the PN, instead of being confined to one functional system, might mediate a large spectrum of signals and functions. Such an organization of the PN suggests that they are well positioned for bringing together and integrating inputs from various functionally diverse cortical areas and subcortical regions (Brodal and Bjaalie 1992; Schwarz and Thier 1999). The multimodal integration at the level of PN does not probably occur in a form of direct convergence of various inputs on single PN neurons, but is rather ensured through bidirectional dynamic coupling between different functional PN patches and the cerebellum and/or through distant connections within the PN (Schwarz and Thier 1999). It is unknown what kind of information processed in the PN finally reaches the CN. However, irrespective of the nature of PN signals to the CN, one can speculate that the resulting influences together with other feedback and feedforward auditory and non-auditory inputs (e.g., auditory cortex, somatosensory inputs) are “weighed” up in the GCD to generate a mosaic of modulatory actions on the output activity of CN principal cells. This kind of modulation would be important for the evaluation of auditory signals with respect to external and internal non-auditory cues.
References
Aitkin LM, Boyd J (1978) Acoustic input to the lateral pontine nuclei. Hear Res 1:67–77
Allen GI, Tsukahara N (1974) Cerebrocerebellar communication systems. Physiol Rev 54:957–1006
Arnott RH, Wallace MN, Shackleton TM, Palmer AR (2004) Onset neurons in the anteroventral cochlear nucleus project to the dorsal cochlear nucleus. J Assoc Res Otolaryngol 5:153–170
Azizi SA, Burne RA, Woodward DJ (1985) The auditory corticopontocerebellar projection in the rat: inputs to the paraflocculus and midvermis. An anatomical and physiological study. Exp Brain Res 59:36–49
Babalian AL, Rouiller EM (2004) Projections from pontine nuclei and auditory cortex exert comparable functional influences on principal cells of the cochlear nucleus. FENS Forum Abstract: 155
Babalian AL, Vibert N, Assie G, Serafin M, Mühlethaler M, Vidal PP (1997) Central vestibular networks in the guinea-pig: functional characterization in the isolated whole brain in vitro. Neuroscience 81:405–426
Babalian AL, Ryugo DK, Vischer MW, Rouiller EM (1999) Inhibitory synaptic interactions between cochlear nuclei: evidence from an in vitro whole brain study. Neuroreport 10:1913–1917
Babalian AL, Jacomme AV, Doucet JR, Ryugo DK, Rouiller EM (2002) Commissural glycinergic inhibition of bushy and stellate cells in the anteroventral cochlear nucleus. Neuroreport 13:555–558
Baker J, Gibson A, Glickstein M, Stein J (1976) Visual cells in the pontine nuclei of the cat. J Physiol 255:415–433
Benson CG, Potashner SJ (1990) Retrograde transport of [3H]glycine from the cochlear nucleus to the superior olive in the guinea pig. J Comp Neurol 296:415–426
Brodal P, Bjaalie JG (1992) Organization of the pontine nuclei. Neurosci Res 13:83–118
Burian M, Gstoettner W (1988) Projection of primary vestibular afferent fibers to the cochlear nucleus in the guinea pig. Neurosci Lett 84:13–17
Doucet JR, Ryugo DK (1997) Projections from the ventral cochlear nucleus to the dorsal cochlear nucleus in rats. J Comp Neurol 385:245–264
Glickstein M, Stein M, King RA (1972) Visual input to the pontine nuclei. Science 178:1110–1111
Hollander H, Tietze J, Distel H (1979) An autoradiographic study of the subcortical projections of the rabbit striate cortex in the adult and during postnatal development. J Comp Neurol 184:783–794
Itoh K, Kamiya H, Mitani A, Yasui Y, Takada M, Mizuno N (1987) Direct projection from the dorsal column nuclei and the spinal trigeminal nuclei to the cochlear nuclei in the cat. Brain Res 400:145–150
Kandler K, Herbert H (1991) Auditory projections from the cochlear nucleus to pontine and mesencephalic reticular nuclei in the rat. Brain Res 562:230–242
Kanold PO, Young ED (2001) Proprioceptive information from the pinna provides somatosensory input to cat dorsal cochlear nucleus. J Neurosci 21:7848–7858
Kevetter GA, Perachio AA (1989) Projections from the sacculus to the cochlear nuclei in the mongolian gerbil. Brain Behav Evol 34:193–200
Kimura A, Donishi T, Okamoto K, Tamai Y (2004) Efferent connections of “posterodorsal” auditory area in the rat cortex: implications for auditory spatial processing. Neuroscience 128:399–419
Knowlton BJ, Thompson JK, Thompson RF (1993) Projections from the auditory cortex to the pontine nuclei in the rabbit. Behav Brain Res 56:23–30
Legg CR, Mercier B, Glickstein M (1989) Corticopontine projection in the rat: the distribution of labelled cortical cells after large injections of horseradish peroxidase in the pontine nuclei. J Comp Neurol 286:427–441
Middleton FA, Strick PL (1997) Dentate output channels: motor and cognitive components. Prog Brain Res 114:553–566
Mugnaini E, Osen KK, Dahl AL, Friedrich VL Jr, Korte G (1980a) Fine structure of granule cells and related interneurons (termed Golgi cells) in the cochlear nuclear complex of cat, rat and mouse. J Neurocytol 9:537–570
Mugnaini E, Warr WB, Osen KK (1980b) Distribution and light microscopic features of granule cells in the cochlear nuclei of cat, rat, and mouse. J Comp Neurol 191:581–606
Oertel D, Young ED (2004) What’s a cerebellar circuit doing in the auditory system. Trends Neurosci 27:104–110
Ohlrogge M, Doucet JR, Ryugo DK (2001) Projections of the pontine nuclei to the cochlear nucleus in rats. J Comp Neurol 436:290–303
Potter RF, Ruegg DG, Wiesendanger M (1978) Responses of neurons of the pontine nuclei to stimulation of the sensorimotor, visual and auditory cortex of rats. Brain Res Bull 3:15–19
Ryugo DK, Haenggeli CA, Doucet JR (2003) Multimodal inputs to the granule cell domain of the cochlear nucleus. Exp Brain Res 153:477–485
Saadé NE, Frangieh AS, Atweh SF, Jabbur SJ (1989) Dorsal column input to cochlear neurons in decerebrate–decerebellate cats. Brain Res 486:399–402
Sasaki K, Staunton HP, Dieckmann G (1970) Characteristic features of augmenting and recruiting responses in the cerebral cortex. Exp Neurol 26:369–392
Schmahmann JD, Pandya DN (1997) The cerebrocerebellar system. Int Rev Neurobiol 41:31–60
Schwarz C, Thier P (1999) Binding of signals relevant for action: towards a hypothesis of the functional role of the pontine nuclei. Trends Neurosci 22:443–451
Shinoda Y, Kano M, Futami T (1985) Synaptic organization of the cerebello–thalamo–cerebral pathway in the cat. I. Projection of individual cerebellar nuclei to single pyramidal tract neurons in areas 4 and 6. Neurosci Res 2:133–156
Shore SE, Helfert RH, Bledsoe SC Jr, Altschuler RA, Godfrey DA (1991) Descending projections to the dorsal and ventral divisions of the cochlear nucleus in guinea pig. Hear Res 52:255–268
Shore SE, Vass Z, Wys NL, Altschuler RA (2000) Trigeminal ganglion innervates the auditory brainstem. J Comp Neurol 419:271–285
Shore SE, El Kashlan H, Lu J (2003) Effects of trigeminal ganglion stimulation on unit activity of ventral cochlear nucleus neurons. Neuroscience 119:1085–1101
Stein JF, Glickstein M (1992) Role of the cerebellum in visual guidance of movement. Physiol Rev 72:967–1017
Voitenko LP, Marlinskii VV (1993) Stereotaxic atlas of the guinea pig brainstem. Naukova Dumka, Kiev, pp 1–16
Wan XS, Liang F, Moret V, Wiesendanger M, Rouiller EM (1992) Mapping of the motor pathways in rats: c-fos induction by intracortical microstimulation of the motor cortex correlated with efferent connectivity of the site of cortical stimulation. Neuroscience 49:749–761
Weinberg RJ, Rustioni A (1987) A cuneocochlear pathway in the rat. Neuroscience 20:209–219
Wiesendanger R, Wiesendanger M (1982a) The corticopontine system in the rat. I. Mapping of corticopontine neurons. J Comp Neurol 208:215–226
Wiesendanger R, Wiesendanger M (1982b) The corticopontine system in the rat. II. The projection pattern. J Comp Neurol 208:227–238
Winter IM, Robertson D, Cole KS (1989) Descending projections from auditory brainstem nuclei to the cochlea and cochlear nucleus of the guinea pig. J Comp Neurol 280:143–157
Wright DD, Ryugo DK (1996) Mossy fiber projections from the cuneate nucleus to the cochlear nucleus in the rat. J Comp Neurol 365:159–172
Young ED, Nelken I, Conley RA (1995) Somatosensory effects on neurons in dorsal cochlear nucleus. J Neurophysiol 73:743–765
Zhou J, Shore S (2004) Projections from the trigeminal nuclear complex to the cochlear nuclei: a retrograde and anterograde tracing study in the guinea pig. J Neurosci Res 78:901–907
Acknowledgments
This work was supported by the Swiss National Science Foundation grant 31-66731.01. The author would like to thank E.M. Rouiller for the helpful comments on the manuscript and C. Roulin for the technical assistance.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Babalian, A.L. Synaptic influences of pontine nuclei on cochlear nucleus cells. Exp Brain Res 167, 451–457 (2005). https://doi.org/10.1007/s00221-005-0178-8
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00221-005-0178-8