
Abstract
It has been suggested that multijoint movements result from the specification of a referent configuration of the body. The activity of muscles and forces required for movements emerge depending on the difference between the actual and referent body configurations. We identified the referent arm configurations specified by the nervous system to bring the arm to the target position both in healthy individuals and in those with arm motor paresis due to stroke. From an initial position of the right arm, subjects matched a force equivalent to 30% of their maximal voluntary force in that position. The external force, produced at the handle of a double-joint manipulandum by two torque motors, pulled the hand to the left (165°) or pushed it to the right (0°). For both the initial conditions, three directions of the final force (0°, +20°, and −20°) with respect to the direction of the initial force were used. Subjects were instructed not to intervene when the load was unexpectedly partially or completely removed. Both groups of subjects produced similar responses to unloading of the double-joint arm system. Partial removal of the load resulted in distinct final hand positions associated with unique shoulder-elbow configurations and joint torques. The net static torque at each joint before and after unloading was represented as a function of the two joint angles describing a planar surface or invariant characteristic in 3D torque/angle coordinates. For each initial condition, the referent arm configuration was identified as the combination of elbow and shoulder angles at which the net torques at the two joints were zero. These configurations were different for different initial conditions. The identification of the referent configuration was possible for all healthy participants and for most individuals with hemiparesis suggesting that they preserved the ability to adapt their central commands—the referent arm configurations—to accommodate changes in external load conditions. Despite the preservation of the basic response patterns, individuals with stroke damage had a more restricted range of hand trajectories following unloading, an increased instability around the final endpoint position, altered patterns of elbow and shoulder muscle coactivation, and differences in the dispersion of referent configurations in elbow-shoulder joint space compared to healthy individuals. Moreover, 4 out of 12 individuals with hemiparesis were unable to specify referent configurations of the arm in a consistent way. It is suggested that problems in the specification of the referent configuration may be responsible for the inability of some individuals with stroke to produce coordinated multijoint movements. The present work adds three findings to the motor control literature concerning stroke: non-significant torque/angle relationships in some subjects, narrower range of referent arm configurations, and instability about the final position. This is the first demonstration of the feasibility of the concept of the referent configuration for the double-joint muscle-reflex system and the ability of some individuals with stroke to produce task-specific adjustments of this configuration.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
After unilateral stroke-related brain damage affecting areas subserving movement control, individuals have a number of deficits in stabilizing arm postures and in producing functional arm movement. A large number of motor deficits can occur such as muscle weakness (Bohannon et al. 1987; Bourbonnais and Vanden Noven 1989; Colebatch et al. 1986), increased tone (Burke 1988; Wiesendanger 1991), and improper spatial and temporal muscle recruitment including inappropriate agonist/antagonist coactivation (Conrad et al. 1985; Dewald et al. 1995; Hammond et al. 1988; Knutsson and Richards 1979; Levin and Dimov 1997; Reinkensmeyer et al. 2002). Sensorimotor deficits also influence the ability to coordinate movement between adjacent joints (Levin 1996). These impairments may be related to altered mechanical properties of motor units (Hufschmidt and Mauritz 1985; Jakobsson et al. 1992), abnormal agonist motor unit activation (Gowland et al. 1992; Tang and Rymer 1981), and deficits in segmental reflex organization such as antagonist inhibition (Hammond et al. 1988), reciprocal inhibition (Yanagisawa and Tanaka 1978), and stretch reflex threshold regulation (Levin and Feldman 1994; Levin et al. 2000). Several studies have examined control of the multijoint arm in individuals with stroke-related brain damage (Beer et al. 2000; Dewald et al. 1995; Kamper et al. 2002; Levin 1996; Roby-Brami et al. 1997; Trombly 1993; Wing et al. 1990). Deficits in multijoint movement have been alternatively attributed to disruptions in interjoint coordination (Cirstea et al. 2003; Levin 1996), the presence of pathological movement synergies (Dewald et al. 1995; Reinkensmeyer et al. 2002), and impaired feedforward control of the passive interaction torques at the elbow joint (Beer et al. 2000). This has led to a somewhat confusing picture of the origin of the multijoint control deficits in individuals with stroke.
A better understanding of motor deficits after stroke-related brain damage might be reached by analyzing them in the context of a physiologically feasible theory of motor control. In this respect, it has been suggested that global factors are used by the nervous system to control all skeletal muscles of the body and its degrees of freedom in a coherent and task-specific way (Feldman and Levin 1995). Specifically, it has been hypothesized that one such global factor is a virtual or referent (R) configuration of the body determined by muscle recruitment thresholds, the latter being specified by neural control levels. In other words, at the R position, the muscles are silent and generate zero active torques. If a joint is moved passively from this position (for example, due to weights of body segments), muscles stretched by the motion are activated and generate torques opposing the deviation whereas the antagonist muscles will be activated when the joint is moved passively in the opposite direction. The system will be deviated from the R configuration until, at some steady state configuration Q, the muscle activity and torques become sufficient to balance the external torques. By shifting the muscle activation thresholds, neural control levels may intentionally influence the R configuration and thus the distance between the R and Q. Reacting to this distance, the muscle-reflex system will generate activity and forces bringing the body to another steady state configuration. The control of muscle activation thresholds thus underlies intentional movements. Another type of threshold control does not influence the R configuration but surrounds this configuration with a spatial zone in which opposing muscle groups are coactivated (the C command; see Levin and Dimov 1997).
Two predictions stemming from the referent configuration hypothesis have been tested and confirmed in healthy subjects. One prediction is that although the Q and R configurations are usually different in the presence of external forces (see above), they may intermittently match each other in dynamic situations, most likely in movements with reversals in direction, resulting in a minimum in the electromyographic (EMG) activity level of the muscles involved. The depth of the minimum is restricted by the C command. Another prediction is related to the fact that some reversals in movement direction may be assisted while others are opposed by external forces. Both types of reversals occur, for example, during jumping. From a mechanical standpoint, by minimizing EMG activity, the system may enhance the role of passive, external forces in the production of movement. The referent configuration hypothesis implies, however, that neural (such as matching the Q and R configurations) rather than mechanical factors alone are responsible for the occurrence of EMG minima and, therefore, EMG minimizations may occur in both types of reversals. These predictions have been confirmed in studies of several whole body movements in humans, head movements in monkeys, and jaw movements in rabbits lending support to the referent configuration hypothesis (Coté et al. 2002; Feldman et al. 1998; Lestienne et al. 2000; St-Onge and Feldman 2004; Weijs et al. 1999). For a more detailed description of the referent configuration hypothesis in the context of the λ model for motor control, see St-Onge and Feldman (2004).
Deficits in the ability of the nervous system to specify and modify the R configuration may provide the theoretical framework from which problems of multijoint movement control may be better understood. Previous studies of the single-joint system of individuals with hemiparesis after stroke have shown that deficits in the specification and regulation of stretch reflex threshold angles (or λs) in elbow flexors and extensors can account for different motor impairments such as spasticity, weakness, and abnormal muscle activation patterns in specific joint ranges delimited by these threshold angles (Levin and Dimov 1997; Levin et al. 2000). These studies showed that the flexor and extensor muscle activation thresholds determined the angular limits within which normal muscle activation patterns could be produced during voluntary attempts to flex or extend the elbow. Attempts to make movements beyond these limits resulted in abnormal patterns of muscle coactivation and problems in movement production. We hypothesized that similar limitations in λs of muscles of adjacent joints would lead to a deficit in the specification of the R configuration which would, in turn, contribute to incapacities in the production of coordinated multijoint movement (Feldman and Levin 1995; Levin et al. 2000).
The purpose of this study was to test this hypothesis by measuring R configurations of the double-joint arm that are specified to reach and hold the same initial position while compensating different loads in patients with unilateral brain damage. Once the initial arm position has been established for a given load condition, the R configuration specified to reach and hold this position was identified using an unloading procedure. Results were compared to those recorded in healthy age-matched control subjects. The method relies on the assumption (empirically justified for single-joint systems; see Feldman and Levin 1995) that responses to unloading with the instruction ‘do not voluntarily intervene’ do not involve changes in central control variables. In terms of the λ model, this means, in particular, that the threshold position at which muscles of a single joint begin to be recruited (or the R configuration for a multijoint system) remained the same during the transition from one steady state to another resulting from unloading. At a steady state, all muscle and external forces (torques) are balanced at a specific position so that, once this state is reached, the system should remain motionless, if stability requirements are met. The combination of the net joint torques and positions of the limb segments in this state is called the equilibrium point (EP) of the system organism-environment. Each EP characterizes a steady state in the interaction of the organism with the environment and thus reflects a common property of the two systems. Therefore, by specifying control variables R and C the nervous system does not predetermine a certain EP. One can say, however, that, once specified, these variables establish a certain range of possible EPs. Represented as points in torque-angle space, these EPs produce a curve or a surface (depending on the number of degrees of freedom) that is called the invariant characteristic (IC) of the muscle-reflex system. A specific EP from this redundant set of EPs emerges in the process of the interaction of the organism with the environment so that, depending on external forces, one or another EP is established.
For a single joint, each IC is a monotonic function relating steady state values of the net joint torque to position when control variables remain constant (Feldman and Levin 1995). For a double-joint system, the steady state torque at each joint is presumably a function of two joint angles and thus it can be represented as a surface in a 3D torque-angle plane. Thus there are two such surfaces that taken together are called the IC of the double-joint system where each surface is a component of the IC. Note that the term ‘invariant’ refers to values of control variables (R and C) for each IC and does not imply that the shape of different ICs is the same.
Finding invariant values of R and C associated with a single IC could not per se justify the suggestion that these are actually control variables. It is necessary to demonstrate that these variables can actually be changed when intentional motor actions are produced. For such a demonstration, one can, for example, ask subjects to intentionally specify a new initial arm position or compensate a new external force at the same initial position and then record a new IC by the unloading method. By comparing the two (or more) ICs one can verify that the values of R and C remained invariant for each IC but changed when the intentional motor action resulted in another IC. This approach was successful in single-joint movement tasks demonstrating that a change in the threshold position underlies voluntary movements in the elbow, knee, wrist, and jaw joints (Feldman 1966, 1979; Ostry et al. 1997). Families of ICs have also been recorded for different muscles in decerebrated cats (Feldman and Orlovsky 1972; Matthews 1959). The unloading method was used in the present study to record two ICs and verify whether or not intentional motor actions resulting in transition from one IC to another involve a change in the R configuration and C command. We required subjects to specify an initial position of the arm while counteracting a predetermined amount of force acting in a particular horizontal direction. Then, for each initial condition, we mapped out the relationship between net static torques and shoulder and elbow joint angles by removing increments of the load in a random order and recording two double-joint ICs (Archambault et al. 2001). We extended this analysis by including individuals with motor deficits due to stroke-related brain damage to investigate the similarities and differences in the ability of the two groups of subjects to specify R configurations of the arm to accommodate different external loads. Some of the results have appeared in abstract form (Mihaltchev et al. 2002).
Materials and methods
Subjects
The study was approved by the ethics committee of the Rehabilitation Institute of Montreal and has therefore been performed in accordance with the ethical standards laid down in the 1964 Declaration of Helsinki. Thirteen individuals with stroke (mean age 51.9±12.1 years; 5 women and 8 men) and 10 healthy subjects (mean age 49.0±9.0 years; 4 women and 6 men) participated in the study after giving their informed consent.
The participants with stroke had right spastic hemiparesis due to a cerebrovascular accident (CVA) in the left hemisphere at least 6 months previously. Only those with lesions in the dominant left hemisphere were studied in order to control for response variability due to differences in task performance by the dominant or non-dominant arm and hand (Chen et al. 1997; Winstein and Pohl 1995). Participants had full passive range of movement at the shoulder, elbow, and wrist, partial control of the arm, and no severe apraxia or receptive aphasia. To perform the experimental tasks, participants did not need to have control over grasping or grip strength. Those with bilateral stroke, pain in the arm or trunk, dysmetria, and static or dynamic tremor were excluded. Patient demographic and lesion location information is presented in Table 1. The healthy group consisted of right-handed individuals with no sensory or motor impairments or orthopedic problems affecting the arm or trunk. Those with uncorrected visual disturbances were excluded.
Individuals participated in a single experimental session consisting of the determination of maximal voluntary force (MVF) followed by an unloading experiment. In addition, participants with stroke underwent a clinical evaluation to determine the sensorimotor status of their affected arm.
Arm motor impairment and spasticity in the elbow were assessed by a qualified physical therapist using two clinical measures:
-
A.
Arm motor impairment was measured with the valid and reliable Fugl-Meyer scale (Fugl-Meyer et al. 1975). Since we focused on arm and not hand movement, we used only the arm section of the scale (36) plus the coordination score (4) for a maximum score of 40 points corresponding to normal function. The test evaluates reflex activity, flexion and extension movement synergies, performance of isolated movements, and coordination and speed of arm movements. According to this scale, 7 (P1–P7) had moderate to severe impairment with scores from 10 to 24 and 6 individuals (P8–P13) had mild motor impairment with scores ranging from 34 to 39.
-
B.
Spasticity of the elbow flexor muscles was assessed with the valid and reliable Composite Spasticity Index (CSI; Goulet et al. 1996; Levin and Hui-Chan 1992; Nadeau et al. 1999). The CSI rates the excitability of biceps-brachii tendon jerks, the resistance to passive elbow extension applied at a moderate speed, and the amount of wrist clonus. Each subtest is rated on 4-point scales while the resistance scale is doubly-weighted since this measure most closely resembles tone. The three scores were summed. Based on clinical experience and results of previous studies, total scores ranging from 5 to 9, 10 to 12, and 13 to 16 corresponded to mild, moderate, and severe spasticity, respectively. According to this scale, 11 participants had mild and 2 had moderate spasticity in the arm.
Determination of MVF
We first measured the MVF produced by the right arm in the two principal directions to be used subsequently in the unloading experiments. The initial force in the unloading experiments was set at 30% MVF. Thirty percent MVF and not maximal forces were used as initial torques since maximal torques in the arm of individuals with hemiparesis are known to be significantly lower than those in healthy age- and sex-matched subjects (Bohannon et al. 1987; Colebatch et al. 1986). Thus, by using 30% MVF, all subjects performed a comparable task in terms of the amount of effort required and fatigue was minimized.
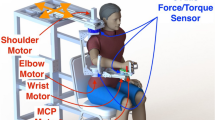
Testing of MVFs was done with the arm in the same configuration as in the initial position for the unloading experiment. To determine MVF, the participant was seated in a chair with a back support and the right shoulder was positioned in front of a handle connected to a force transducer fixed to a shaft of adjustable height (see Fig. 1A). Pulling and pushing arm forces acting in a horizontal plane were measured at the hand. The hand was placed at the shoulder level in the same plane. The shoulder was in 90° flexion and 45° horizontal adduction (0° is full horizontal abduction defined with the arm in line with the subject’s right and left shoulders). The elbow angle was about 45° (full elbow extension was defined as 180°) and the wrist was in the neutral position between supination and pronation. The left arm was alongside the body. No compensation in the form of trunk forward or lateral inclination was permitted during the testing. Participants performed three consecutive trials per direction separated by 1-min pauses. For the pushing and pulling directions, the subject pushed the handle horizontally to the left or right, respectively. Off-axis forces were not recorded. The participants started to push or pull on a verbal signal and were encouraged vocally by the examiner to maintain the maximal force for 3 s. The force signal was monitored on an oscilloscope (Tektronix, type RM 561A) and the means of the three trials in each direction were used in calculations of 30% MVF. In those participants with hand paresis (P1–P7), it was necessary to attach the hand to the handle of the force transducer with a specially made mitt but no participant required any additional support of the arm or forearm to maintain their arm in the required position.

Schematic diagram of the experimental set-up and examples of non-corrected and corrected responses to unloading. A The subject sat in front of a computer screen with their trunk strapped to the back of the chair and their right forearm and hand supported by a splint attached to the handle of a double-joint manipulandum controlled by two torque motors. Subjects were required to match the force of the motors by pushing the handle to the left (165°) or pulling it to the right (0°). For both initial conditions, three directions of the final force were used, 0°, 20°, and −20° with respect to the direction of the initial force. B, C Hand velocity/position (phase) diagrams showing uncorrected (B) and corrected (C, upper arrow) responses to unloading in one healthy subject (H6). X position refers to the horizontal displacement in the sagittal direction. Position and velocity shown in B were scaled for C by factors of 2.5 and 2, respectively. Arced arrows in B and C show the movement direction
Experimental set-up
The lengths of the right arm from the acromion to the lateral epicondyle of the elbow and of the forearm from the lateral epicondyle to the distal end of the first metacarpo-phalangeal joint were measured in order to calculate the torques at elbow and shoulder joints in later analysis. For the unloading experiment, the participant was seated on an adjustable chair in front of a computer screen. The chair was placed so that in the initial position, the hand was at a distance of 30 cm from the subject’s sternum. The initial position of the arm was identical to that used to determine MVFs (see above). The trunk was attached to the back of the chair by two 10-cm-wide Velcro straps to avoid trunk movements often observed in individuals with hemiparesis during arm movements (Cirstea and Levin 2000; Michaelsen et al. 2001; Roby-Brami et al. 1997). The right hand, wrist, and forearm were placed in a polypropylene bi-valve splint attached to the handle of a manipulandum. The splint prevented relative movements of the wrist, forearm, and hand leaving only the shoulder and elbow free to participate in the movement. It also assured a firm grip on the handle for those who had disturbed control of grasping (P1–P6) and partially supported the weight of the arm against gravity. Both participants with hemiparesis and healthy subjects used the same splint during the experiment. The double-joint manipulandum was controlled in the horizontal plane by two torque motors (Mavilar motors MT2000), each motor creating torque at one joint of the manipulandum independently of the torque produced at the other joint. A software procedure was developed to reduce the effects of the manipulandum’s inertia on hand movement. Specifically, positive feedback was introduced in the torque output, based on the acceleration (directly recorded with accelerometers) and the moment of inertia of each segment of the manipulandum. The feedback factor was selected by trial and error so as to reduce inertia without introducing oscillations. The moment of inertia was measured experimentally for each segment of the manipulandum by applying a torque pulse of known value and measuring the angular velocity. This measurement was repeated after the introduction of the inertia-correcting feedback. In this way, the effective moment of inertia was reduced from 0.209 to 0.120 kg m2 (reduction of 62%) for the proximal limb of the manipulandum, and from 0.038 to 0.031 kg m2 (reduction of 18%) for the distal segment.
Three safety features were incorporated into the apparatus. First, interrupters were placed near the limits of the movement of each segment of the manipulandum (about 110° in the horizontal plane for each articulation), which automatically switched off the motors when the segment reached these limits. Second, the motors were automatically turned off if the speed of the manipulandum approached the limit of the natural speed of the arm and hand (2 m/s). Third, subjects held a ‘panic’ button that instantly interrupted the electrical supply to the torque motors when pressed. Finally, the torque in each motor was limited to about 30% (15 Nm) of its maximal output.
Experimental procedure
The task was to bring the handle of the manipulandum to the specified initial position and stabilize it while balancing an external load, which was subsequently reduced or removed. The resultant final positions of the handle and joint torques after different levels of unloading were used to construct double-joint torque/angle characteristics. Two characteristics were measured: one when the subject’s intent was to push the initial load (PUSH) and the other when the direction of the initial load was changed so that balancing the load required the subject to change the direction of the applied force (PULL). By specifying different initial load directions, we implicitly induced subjects to produce different intentional actions to bring the hand to the same position before unloading while compensating different loads. In terms of the λ model, subjects were forced to specify different ICs to accommodate the different initial conditions.
To place the hand at the initial position, the subject moved the handle of the manipulandum until the position of the hand, indicated by a cursor on the computer screen, reached a target indicated by a 2-cm red circle at the center of the screen. Elastic resistance was applied to the handle by two torque motors. The resistance increased linearly as the hand approached the target so that when the hand reached the target, the load corresponded to its maximal value (30% of the MVF).
The direction of the initial load was set so that subjects either had to pull (0°) or push (165°) against it (Fig. 1A, B). A load direction of 165° was chosen for pushing since pilot studies showed that when a pushing load directly opposite to the pulling load (180°) was used, some unloading conditions caused the subject’s arm to hit their body. Unloading was performed first for PULL and then for PUSH. The magnitude of the load at the initial target corresponded to 30% of each subject’s MVF, measured prior to the experiment for each direction. In addition, in order to determine if any subsequent differences in behavior between groups were due to differences in initial load levels, we recorded data from an additional 3 healthy subjects (‘low-load’ control group: 3 men, mean age 55.3±13.3 years) using initial loads similar to the mean initial loads of the stroke group.
For the PUSH condition, the arm would move to the left in the horizontal plane, in the direction of 165° to the frontal plane (Fig. 1A), if it were not opposed by the load. For the PULL condition, the arm pulled the load to the right (0° to the frontal plane). The subject was thus required to match the initial force in order to maintain the hand at the target position.
After a randomized delay (2–4 s), the load force was unexpectedly decreased, resulting in motion of the arm to a new position. The subject was instructed not to intervene, i.e., let the arm go in a natural way to a new position without trying to intentionally modify the position. We used an unloading instead of a loading paradigm since stretching of active muscles produced by sudden loading elicits protective voluntary or triggered reactions associated with changes in central commands (Feldman and Levin 1995). The participants had full vision of their arm and of the computer monitor during target attainment. However, the position of the cursor on the screen was not updated after changes in the load. It has been shown that, with this instruction, the changes in the EMG activity, muscle forces, and arm position are reflex-like reactions to changes in the load, typically not involving modifications of the central control signals in the sense defined in the λ model (see above; Feldman and Levin 1995). Several training trials (5–15) were done before data were recorded. Surprisingly, participants with hemiparesis required fewer practice trials (about 5) than healthy subjects (about 10–20) since, due to longer reaction times, they were less able to voluntarily correct their hand position in response to the rapid unloading. In contrast, healthy subjects had a tendency to ‘help stabilize’ the arm position following the unloading. Practice ended when subjects reliably produced uncorrected movements characterized by a smooth transition to a new position with a single peak in the hand velocity profile for at least three consecutive trials. In the subsequent experiment, trials in which subjects intervened and corrected the arm position following unloading could be identified by visual inspection as those in which inflections (one or several additional loops) in velocity-position diagrams occurred (compare trace in Fig. 1C to traces in Fig. 1B for a healthy subject, H6) or when terminal oscillations were present, a drift in the mean position of the oscillations was observed. Whether or not the subject intervened was determined by the experimenter by inspecting the phase diagram of the just completed trial. Overall, 15–20% of trials in each group were excluded on-line in this manner. Excluded trials were repeated so that the total number of recorded trials per subject was the same.
We anticipated that, for each initial condition, the relationship between torques and joint angles in steady states resulting from unloading would be described by a smooth surface in torque/angle coordinates (i.e., an IC plane). We used a large number of unloading conditions in order to better characterize this plane. For each of the initial loads, there were 12 unloading conditions. Six of these involved a change of load magnitude while maintaining its initial direction (unloading to 60%, 40%, 20%, 10%, 0%, and −10% of the initial value). For the other six conditions, both the magnitude and direction of the load were changed in randomly selected trials (unloading to 40%, 20%, and −10% of the initial value with a ±20° change in direction relative to the direction of the initial load). Blocks of 12 trials were used, in which each of the 12 final load conditions was presented once, in a random order. This was repeated six times (72 trials for each initial direction of the load). The experiment thus consisted of 144 trials and lasted 2.5–3 h. The 12 data points for each initial direction (PUSH and PULL) were used to determine two referent (R) arm configurations accordingly. In principle, a greater number of R configurations could have been obtained using additional initial load directions and hand positions. However, because of the lengthy recording procedure, this was not feasible in individuals with stroke. Indeed, since fatigue may lead to increased response variability especially in individuals with stroke, subjects were allowed 2- to 5-min periods of rest every 30 min or less if desired. The development of fatigue was monitored by verifying that throughout the experiment, all subjects were able to maintain initial force levels of 30% MVF constant.
Data recording and analysis
The position of four active infrared-emitting diodes (IREDs) placed on the acromion processes of the two shoulders, the lateral epicondyle of the right humerus, and the handle of the manipulandum were recorded using an Optotrak Motion Analysis System (Northern Digital, model 3010, Waterloo, Ontario; sampling rate 100 Hz, 3 s/trial). The angular position and velocity of the segments of the manipulandum were measured with two axial resolvers. The shoulder and elbow angles in the horizontal plane were calculated based on the scalar products of the vectors joining the appropriate IREDs. Velocity and acceleration of the handle were computed using x-y positional data. Movement onsets and offsets were defined as the times at which the hand velocity exceeded and remained above and then decreased and remained below 20 m/s, respectively. Torques generated by the motors were measured by strain gauges incorporated into the axes of rotation. Using these torque values and the basic geometry of the arm-manipulandum system, we computed shoulder and elbow torques and the forces applied to the handle. From the joint angles and force at the level of the handle, static elbow and shoulder torques before the onset and after the end of movements were calculated. These static torques were derived from the strain gauge data on the manipulandum’s axis, as well as data on the arm configuration at respective steady state positions. To accurately estimate joint torques during movement would have required taking into account the dynamics of the arm-manipulandum system (inertia, elasticity, viscosity), which was unnecessary since we were mostly interested in steady states of the arm before and after unloading. In most cases, the hand achieved a stable position after unloading within 1 s. However, in some individuals with stroke, unloading resulted in terminal oscillations about the final position that achieved a steady state after two or three cycles at which point, the final position was measured. In the few cases when the hand continued to oscillate until the end of the trial, the mean final positions were measured as the midpoint of the last oscillatory cycle.
Electromyographic data were recorded from six pairs of bipolar surface electrodes. The activity of brachioradialis (BR), anconeus (AN), biceps brachii (BB), the lateral head of triceps brachii (TB), posterior deltoid (DP), and the clavicular portion of pectoralis major (PM) were recorded. Electrodes were placed so that crosstalk contamination was eliminated by observing the response to isolated and associated contractions of the target muscle. EMG signals (Grass Instruments, Quincy, MA) were amplified (gain 10–20), filtered (5–500 Hz), and sampled at a rate of 1,500 Hz. The signals were filtered offline using high-pass filters (cutoff=35 Hz) to remove motion artifacts and rectified. The level of tonic activity after unloading was calculated as the root-mean square (RMS) value of the EMG activity in a 100-ms window centered at the 2.5-s mark of the recorded trial. To determine coactivation ratios, the RMS value of the antagonist activity was expressed as a percentage of the total agonist plus antagonist activity.
Instability index
Tangential velocity profiles of the hand were used to determine instability indices for each subject based on the amplitude of oscillations around the final hand position after unloading. We computed the inverse of the logarithmic decrement of decay (D) of the oscillations of the arm (Levin and Dimov 1997):
where V1 and V2 are the velocities of the first overshoot and the first undershoot of the hand around the target after the peak velocity and T12 is the time between them (Fig. 2). The decrement is related to the damping and stiffness of the system so that higher values indicate decreased damping or greater oscillations. The instability index is defined as the inverse of the decrement (1/D)—the time required for the velocity oscillations to decrease in amplitude by a factor of 1/e≅0.37. For example, 1/D=0 if the transition from one position to the other is produced aperiodically, i.e., without terminal oscillations, and 1/D=∞ if oscillations are self-sustained, i.e., proceed without decay.

Representative velocity profiles of hand movements in the PUSH direction for one healthy subject and one individual with stroke (patient 6, Fugl-Meyer score FM=24/40). In the healthy subject, the hand came to a stable final position after a small terminal overshoot (V 1 ) and undershoot (V 2 ). In the participant with stroke, the peak velocity was lower and terminal overshoots (V 1 , V 2 ) were greater. The instability index is defined as 1/D where D was computed as shown in the figure
Torque/angle characteristics and R configurations of the arm
For the reconstruction of double-joint ICs, we used the same procedure as that described in Archambault et al. (2001). The procedure was as follows: for each initial direction of the external force (PULL or PUSH), the net static torque generated at each joint before and after partial or full unloading was considered as a function of the two (elbow and shoulder) joint angles. Different conditions of unloading served to map-out the single-joint IC for the shoulder and elbow. The combinations of net static torque for one or the other joint together with the respective values of the two joint angles measured before and after unloading were plotted as points in Cartesian 3D space (shoulder or elbow torques versus shoulder and elbow angles; the bar diagrams on Fig. 6A, B represent elbow or shoulder torque produced at a specific position of those two joints). Using regression analysis, we computed the best fitting 3D planar surface for the set of points associated with torques of the respective joint. For each initial direction of the external force (PUSH or PULL), two surfaces were thus obtained, one for the shoulder and the other for the elbow. In fact, these surfaces separately represent the single-joint IC for the shoulder or elbow, respectively. Taken together, however, they construct an IC of the double-joint system. These surfaces were characterized by slopes representing stiffness coefficients (two per each single-joint IC). For example, each elbow torque/angle characteristic was characterized by two stiffness coefficients, See and Ses (Nm/rad), where the first subscript (e) refers to the elbow joint from which the net joint torque was measured, and the second subscript refers to the angle that influences this torque (e, elbow or s, shoulder; for example, Ses represents the stiffness coefficient for the elbow torque in relation to shoulder angle).
The intersection of each surface with the elbow-shoulder angle plane (the denser mesh on Fig. 6A, B), represents the combinations of the two joint angles at which arm muscles generate zero net torque at the respective joint—a zero torque line. For each initial condition there were two surfaces and thus two zero torque lines. The point of intersection of these lines represents the configuration of the arm at which all joint torques are zero. By definition, this point is a referent (R) configuration of the arm (see Introduction). The R configurations were compared across initial conditions and groups of subjects.
We defined torques that flexed the elbow and adducted the shoulder as being positive.
Statistical analysis
Student t-tests were used to compare variables between groups or conditions (initial arm positions and torques, peak velocities, spatial dispersions of the trajectories). To confirm that the final hand positions were distinct points lying on the IC for each joint and that they depended on the amount of unloading, we reduced the two-dimensional position data to a single dimension. The mean final hand position was first normalized to its maximum displacement along the x-axis and then three third-order polynomials were fit to the final endpoint positions, one for each set of unloading conditions with the same direction of the final load (change with respect to the direction of the initial load of 0°, +20°, or −20°). To each set of points, we added the initial endpoint position and the position after complete unloading (i.e., the steady state points also observed in the same unloading experiments). The order of the polynomial fits was selected as the minimum order yielding a statistical fit in all cases (P<0.05, with a mean r2 of 0.87). The distances along the fitted curve to each final endpoint position were considered trajectory lengths for each unloading condition and were computed for the two initial load directions as well as for each of the three directions of the final load. Differences between endpoint positions were determined with repeated measures ANOVAs and post hoc contrasts using the Bonferroni procedure.
Three-dimensional torque versus angle surfaces were constructed using regression analysis and their correlations and slopes were compared between conditions and groups with Student t-tests. To determine the dispersion of referent configurations in Cartesian space, we applied geometrical analysis and compared the slopes and areas of the computed 95% confidence ellipses with Student t-tests. The instability indices were compared between groups by Student t-tests. Relationships between coactivation ratios, instability indices, and levels of clinical impairment were determined by Pearson product moment statistics. Non-parametric statistics (Mann-Whitney U-test and Kruskal-Wallis ANOVA) were substituted when requirements for homogeneity of variance (Levene’s test) were not met. These instances are indicated throughout the text. The significance level was set at P<0.05 for all tests and Bonferroni corrections for multiple comparisons were made.
Results
Responses to unloading: general characteristics
The mean maximal forces (MVF) produced by the healthy group were 82.6±33.2 N for PUSH and 79.2±51.7 N for PULL which were 41% and 65% higher (P<0.05) than those of the stroke group (Table 1). Based on these forces, the initial loads (30% MVF) used in the unloading experiment ranged from 12.0 to 39.9 N (mean=24.8±9.3 N) for PUSH and 11.0 to 37.7 N (mean=19.8±8.1 N) for PULL in healthy subjects and from 6.3 to 37.0 N (mean=17.8±10.5 N) for PUSH and 5.0 to 29.6 N (mean=14.2±10.0 N) for PULL in individuals with hemiparesis. In order to determine if any subsequent differences in behavior between groups were due to differences in initial loads, we recorded data from an additional three healthy subjects (‘low-load’ control group) using initial loads similar to the mean initial loads of the stroke group. Thus, the initial loads for the low-load control group were 17.0 N for PUSH and 14.0 N for PULL. Throughout the recording session, initial loads were constant and were easily reproduced by all subjects.
In healthy subjects, after complete or partial removal of the load, the hand began to move and, after a transient overshoot, reached a new position at which the residual load was balanced after about 1 s (Fig. 4A–E). Training effectively diminished the number of trials in which subjects made voluntary corrections sometimes present in practice trials (see inflection point in Fig. 1C). During the testing period, trials with corrections were eliminated on-line (see Materials and methods).
For all groups, the hand displacement was in the direction opposite to that of the initial load (Fig. 3). Figure 3A, B shows that hand trajectories were slightly curved. Similar responses to unloading were observed in individuals with stroke but, in the latter, hand trajectories at the end of movements were sometimes more hook-shaped (Fig. 3C, D) and terminal overshoots and oscillations were more marked (Fig. 4F–H; see Instability indices below).

Examples of mean trajectories for each of the 12 unloading conditions for the pushing (left panels) and pulling (right panels) directions in a healthy subject (A, B) and in participants with mild (C, D) and moderate (E, F) arm motor impairment

Representative kinematic data (means by condition) showing responses to six different combinations of unloading in the pushing direction (165°) in one healthy subject (left panels) and one participant with moderate arm motor impairment (patient 6, Fugl-Myer score, FM=24/40). A, F Hand displacements. B, G Tangential velocities. C, H Hand velocity/position phase diagrams. D, I Shoulder horizontal adduction angles. E, J Elbow extension angles
Trajectory lengths (Figs. 3, 5) and joint angles (Fig. 4D, E, I, J) changed monotonically with increasing amounts of unloading in each subject for each of the three directions of the final load. Changes in arm position were observed in response to even the smallest decreases in the load (10% of the load; 4.7 N in the healthy group, 2.0 N in the stroke group, and 5.6 N in ‘low-load’ control group), showing that the neuromuscular system was sensitive to even small perturbations. Hand trajectories and final position depended on the change in load. Hand trajectories for the subject in Fig. 3A, B were slightly curved for both directions. Different levels of unloading without any change in load direction brought the hand to final points located at the same curve at distances gradually increasing with the amount of unloading as shown by different and distinct trajectory lengths for both groups of subjects in Fig. 5. There were no differences between the trajectory lengths (mean of all 12 combinations) between all three groups for PUSH or for PULL (Table 2). Repeated measures ANOVAs on the final positions after unloading performed for each subject for PUSH and PULL conditions showed that the majority (>60%) of intermediate positions were significantly different from each other suggesting that there was a unique relationship between final position and load.

Mean (+SD) trajectory lengths for the pushing (left panels) and pulling (right panels) directions for the 12 combinations of unloading in healthy subjects (top panels) and in participants with hemiparesis (bottom panels). The first six bars in each panel show data for the six different levels of unloading when the direction of the final load coincided with that of the initial load (0°). The next six bars show data for different levels of unloading when the direction of the final load was changed by −20° (next three bars) and 20° (last three bars)
To determine if the variability of final positions after unloading was affected by fatigue, we compared the mean final positions in the main (x) direction in the first three trials with that of the last three trials in two unloading combinations (40% and full unloading). Comparisons of these results were not significant in all but one healthy subject in whom the final position in the complete unloading condition shifted by 40 mm in the second half of the experiment. In the stroke group, only two subjects had a drift in final position, one of 28 mm for the 40% condition and another of 38 mm for the full unloading condition. This analysis suggested that, in the majority of cases, variability in the final positions may not have been related to the development of fatigue in either subject group.
For complete unloading, the mean maximal peak velocities of the hand were higher in the healthy compared to the stroke group only for the PUSH condition (t22=−2.23, P<0.04; Table 2). Values of the low-torque control group were similar to those of the stroke group (Table 2).
Shoulder torques decreased monotonically with the increasing amount of unloading for both initial conditions and groups of subjects. While elbow torques also changed monotonically with the change in the load for the PULL condition, this was not always the case for the PUSH condition in seven individuals (P1, P2, P3, P5, P6, P7, P10) with hemiparesis.
Changes in shoulder and elbow angles as a result of unloading for both conditions in each of the three groups are shown in Table 2. In response to unloading, the shoulder adducted in all subjects for PUSH and abducted for PULL. However, the shoulder movement was combined with elbow flexion and extension differently in each individual with no differences between the groups. Subjects combined shoulder horizontal adduction with elbow extension for unloading in the PUSH condition (67% of the healthy group, 70% of the stroke group, 67% of the low-load control group) and shoulder horizontal abduction with elbow extension for unloading in the PULL condition (83% of the healthy group and 80% of the stroke group). For the PULL condition, subjects in the low-torque control group combined shoulder horizontal abduction with slight elbow flexion.
The hand trajectories in most participants with stroke were restricted in terms of their variability as estimated by their dispersion in the sagittal direction (for example, Fig. 3E, F). For each of these individuals, the spatial dispersion was smaller for both PUSH and PULL conditions compared to the healthy subjects (Table 2). The dispersion of trajectories in the low-load control group was also significantly smaller than the healthy group but larger than that of the stroke group.
Torque/angle characteristics and referent configurations of the double-joint arm
The net static muscle torque at each joint was considered as a function of two joint angles (elbow, shoulder) that could be represented as a planar surface in the 3D torque versus joint angles space. We constructed two such surfaces (one for the elbow and one for the shoulder torques for each condition for a total of four surfaces per subject). The points forming each surface were obtained by averaging the final joint torques and angles for all trials per condition. Together with the point representing the initial condition, 13 points were used to construct each of the four torque/angle 3D surfaces. Using regression analysis, we approximated each surface by a plane. Examples of the planar fits obtained for the elbow and shoulder torques with the pushing load are shown in Fig. 6 for a healthy subject and in Fig. 7 for a subject with mild hemiparesis. The regression analyses yielded significant r2 values (0.83±0.12, range 0.48–0.99) in all healthy subjects using either regular or low initial torques (Table 3). In the participants with hemiparesis, the r2 values were lower (0.70±0.26, range 0.04–0.97). Individuals P7 and P8 had non-significant r2 values for the elbow torque/angles surface for PUSH (r2 values were 0.33, 0.09) and two other individuals (P2 and P3) had non-significant values for the elbow torque/angle surface for PULL (0.04, 0.33). One of these participants also had a non-significant r2 value (0.28) for the shoulder torque/angle surface for PULL. There was no correlation between clinical severity and r2 values. Thus, the lack of ability to specify an invariant torque/angle relationship in these four individuals was not related to their level of clinical impairment (Table 1). Since these participants could not reliably specify an IC, their data could not be used for estimation of the referent configurations (see below and Discussion).

Torque/angle characteristics of the arm in one healthy subject, for the pushing direction. A Elbow torque as a function of two joint angles (elbow and shoulder) approximated by a planar surface (wide grids). 3D bars indicate torque/angle combinations for each condition (0°, gray; +20°, black; –20°, white).The intersection of the planar surface with the zero torque plane (narrow grids) forms a line that describes all combinations of elbow and shoulder angles (arm configurations) at which the elbow torque is zero for this condition. B Same as in A but for shoulder torque

Torque/angle characteristics in one participant with stroke with mild hemiparesis (patient 9, Fugl-Meyer score, FM=36/40) for the pushing direction. Notations as in Fig. 6
Each regression surface was characterized by two slopes and there were two regression surfaces per condition (see Materials and methods). The slopes (S) for the PUSH condition were different from those for the PULL condition, both in healthy subjects and in participants with stroke (Table 3) confirming the suggestion of the EP hypothesis that ICs were different for different initial load directions. Between groups comparison, however, revealed significantly lower slopes (i.e., low stiffness) for individuals with hemiparesis compared to healthy subjects for See in the PUSH condition and for Sse and Sss in the PULL condition (Table 3) while there were no differences in slopes between the healthy and low-load control group.
The range of combinations of elbow and shoulder angles for which the respective joint torque is zero was described as the intersection of each regression surface with the angle plane. One zero torque line for each joint was computed. Zero torque lines are shown as thick straight lines in Figs. 6A and 7A for the elbow and in Figs. 6B and 7B for the shoulder. The intersection of the two zero torque lines described a point (Fig. 8 open circles) at which both elbow and shoulder torques were zero. This point represents the configuration that the arm reaches when the initial load is completely removed and, in terms of the EP hypothesis, the point that describes the referent configuration (R) of the arm. According to the EP hypothesis, the neuromuscular system generates torques when the actual arm configuration (Q) deviates from R where the amount of torque generated is a function of the difference between R and Q.

The referent configurations of the arm determined from the data shown in Figs. 6 and 7. In the left panels, the intersection of the two zero torques lines in elbow-shoulder joint space for the elbow (dashed lines) and shoulder (solid lines) taken from Figs. 6 and 7 identify the unique, referent configuration of the arm (open circle, R) at which all joint torques are zero. The solid square shows the arm configuration at the initial position of the hand (Q). Right panels As in left panels, but shown in the coordinates of external space. Stick diagrams show the actual positions of arm segments at the R and Q configurations
For comparison, the final arm configurations of all participants (Fig. 9 open circles) were plotted in the same joint space for the PUSH and PULL conditions for both groups of subjects. The areas occupied by the R configurations of the stroke and healthy participants were different for PUSH and for PULL, implying that subjects changed the referent configuration of the arm to accommodate the changes in the initial load conditions. The dispersions of R were characterized by 95% confidence ellipses of different orientations (slopes) and sizes (areas) listed in Table 4. R configurations from the ‘low-load’ control group fell within the ellipses of the healthy group. In participants with hemiparesis, the area was smaller for PUSH while it was oriented differently for PULL compared to healthy subjects (Table 4).

Referent arm configurations in elbow versus shoulder angle coordinates for the pushing (squares) and pulling (circles) directions for all healthy subjects (open symbols) and participants with stroke (filled symbols). The 95% confidence ellipse for each group of points is indicated. Open hexagons show the initial configurations of the arm (Q) that was used in the experiments. Note that the clusters of points obtained for the two initial conditions are clearly separated, implying that subjects modified the referent configurations of the arm to accommodate the changes in these conditions
Stability of the end-point position after unloading
In healthy subjects and in the low-load control group, the hand stabilized in the final position after a transient overshoot without oscillating around the final position (Figs. 1B, 4A, C, E). Mean instability indices (1/D, see Materials and methods) for all 12 conditions in the healthy group were 240±5 ms for PUSH and 280±6 ms for PULL (Table 5) and were similar in the low-load control group (320±137 ms for PUSH and 234±74 ms for PULL; ANOVA, P>0.05). In contrast, the movement in participants with hemiparesis usually terminated after several cycles of oscillations (Fig. 4F, H, J). Compared to the healthy and low-load control groups, the stroke group had significantly larger instability indices for PUSH [582±185 ms; H(2, n=36)=21.0, P<0.001] and for PULL [338±63 ms; H(2, n=36)=8.89, P<0.02]. In addition, only in the stroke group, the values for PUSH were significantly larger than those for PULL (U=2.0; P<0.001).
In the four participants with stroke identified previously as having insignificant r2 values for the elbow torque/angle surfaces, a distinguishing feature was that they also had the highest mean instability indices ranging from approximately 400 to 645 for PUSH (P6, P7) and from 1,000 to 1,410 for PULL (P2, P3).
Electromyographic activity
The roles of the two single-joint muscles (PM and DP) acting at the shoulder were clearly classified as agonists or antagonists according to whether there was a silent period or a stretch reflex, respectively, in response to unloading. According to these criteria, PM acted as an agonist and DP as an antagonist for PUSH and vice versa for PULL. The identification of the roles of the two single-joint elbow muscles was more difficult because of the variability of the EMG activity. In the majority of cases, AN was an antagonist and BR was an agonist for PUSH and vice versa for PULL. For the two double-joint muscles (BB and TB), a consistent main role was undetermined since both muscles were active throughout the unloading. For convenience, we assigned BB the role of agonist and TB the role of antagonist for PUSH and vice versa for PULL conditions.
In healthy subjects, the tonic EMG activity of all muscles changed systematically with the reduction in the external load (Fig. 10 top rows). The modulation of EMG activity was less systematic in the individuals with stroke (Fig. 10 bottom rows). Mean coactivation ratios for each AG/ANT pair were computed for each of the six levels of unloading in the 0° direction and values for single joint elbow (BR/AN) and shoulder muscles (PM/DP) are shown in Table 5. In general, coactivation indices were higher in elbow muscles for PUSH (P<0.0001 for four out of six levels; see Table 5) and in shoulder muscles for PULL (P<0.0002 for five out of six levels). There was no correlation between coactivation and instability indices indicating that any level of unloading could be associated with any level of coactivation.

Smoothed and rectified EMG activity for PUSH in one healthy subject (top) and one participant with moderate motor impairment (patient 3, Fugl-Meyer score, FM=15/40; bottom). In each panel, the individual muscle activity for the six conditions of unloading in the 0° direction are shown in descending order from the top (60% unloading) to the bottom (−10% unloading). The intermediate levels are 40%, 20%, 10%, and 0% of unloading. Muscle activity from brachioradialis (BR), anconeus (AN), biceps brachii (BB), triceps brachii (TB), pectoralis major (PM), and posterior deltoid (DP)
In individuals with stroke, coactivation ratios in both muscle groups were not different from those in healthy subjects for the PUSH direction, but those of the shoulder muscles were significantly lower for PULL (F(1,5)=6.11, P<0.02). The lowest levels of coactivation were found in the four subjects in whom ICs could not be identified. Indeed, participants with stroke did not increase coactivation in the shoulder muscles for PULL as did the healthy subjects. Also unlike healthy subjects, the amount of instability for PUSH was inversely correlated with elbow coactivation (BR/AN, r=−0.58), while for PULL there was a significant positive correlation between the level of instability and the amount of shoulder muscle (PM/DP) coactivation (r=0.60).
The amount of instability following unloading was also related to the clinical impairment level (FM scores) in the individuals with stroke. For PUSH the arm impairment level was negatively correlated with instability indices (r=−0.60) as well as with the elbow (BR/AN) coactivation ratio (r=−0.58). For PULL, there was only a significant correlation between arm impairment and shoulder muscle (PM/DP) coactivation (r=−0.57).
Discussion
Basic findings
Healthy subjects and most participants with arm paresis (8 of 12) responded similarly to unloading of the double-joint arm system. Even in participants who used low initial torques (Table 1), partial removal of the load resulted in distinct final hand positions (Fig. 5) associated with unique shoulder-elbow configurations and joint torques. The net static torque at each joint before and after unloading could be represented as a function of the two joint angles describing a planar surface in 3D torque/angle coordinates, or IC (Figs. 6, 7). Subjects were also able to specify different ICs related to the two different initial conditions (PUSH or PULL). This indicates that most individuals with stroke-related brain damage and hemiparesis preserved the ability to adapt their control variables to accommodate changes in the load conditions. One of these control variables, the referent configuration of the arm, was identified based on two criteria: it changed according to the initial conditions and it remained invariant for the whole set of unloading responses obtained for the same initial condition. This invariance justifies the term invariant characteristic (IC) applied to the torque/angle relationships associated with this set of responses.
The referent configuration of the arm describes the combination of the elbow and shoulder angles at which all joint torques are zero. This analysis thus shows that the net static torque generated at each joint is a function of the difference between the actual and the referent configurations of the arm and that the adaptation to the initial condition was produced by appropriate adjustments in the referent arm configuration. With this adjustment, similar responses to unloading were reproduced but in another part of the arm workspace. The finding that the slopes of the ICs were different for different initial conditions suggests the involvement of an additional control variable presumably modifying the degree of coactivation of opposing muscle groups. It has been shown that non-linear torque/angle characteristics recorded at the elbow joint may be transformed to linear characteristics by the coactivation of opposing muscle groups (Asatryan and Feldman 1965; Levin and Dimov 1997). Coactivation was likely responsible for the linearization of the torque/angle characteristics in our experiments so that the IC surfaces could be presented as planes with high r2 values.
Despite the preservation of the basic response patterns, individuals with stroke had a more restricted range of hand trajectories following unloading (Fig. 3), an increased instability around the final endpoint position (Fig. 4), differences in the dispersion of referent configurations in elbow-shoulder joint space (Fig. 9), and an altered pattern of elbow and shoulder muscle coactivation compared to healthy individuals. Moreover, in 4 out of 12 individuals with stroke, ICs could not be identified, implying that these individuals could not specify referent arm configurations in a consistent way. Distinguishing features of these 4 individuals were greater instability about the final position after unloading and lower levels of coactivation at the elbow and shoulder. Healthy subjects using lower torques did not have behavior similar to participants with stroke using correspondingly low loads, suggesting that the differences in the ranges of torques used might not be the cause of the differences in the unloading responses in the two groups of subjects.
‘Do not intervene’ paradigm
Our data from healthy subjects are consistent with previous observations that responses to unloading leading to transitions to a new position at which equilibrium with the final load is achieved, are very robust and reproducible (Asatryan and Feldman 1965; Forget and Lamarre 1987). In naïve subjects, they can be observed when no instruction is given. With repetition, however, subjects may try to correct unloading responses. The comparatively short latency of voluntary corrections observed in healthy subjects (about 150 ms) suggests that corrections could be initiated before movement offset in the present paradigm in which the movement time was about 1 s. Reaction times for corrections of elbow positions in individuals with hemiparesis are reportedly longer (later than 400 ms; Dickstein et al. 1993) making it less likely that individuals with stroke could correct their responses. In order to decrease the likelihood of voluntary corrections, subjects were instructed not to intervene to the perturbation. In addition, we used unloading instead of loading perturbations since it has been shown that subjects often generate involuntary (‘triggered’) corrective responses to the latter (Fig. 4 in Feldman and Levin 1995; see also Crago et al. 1976; Newell and Houk 1983).
Several findings indicate that subjects did not make corrections to unloading in the present experiment. All subjects were given adequate practice until uncorrected responses were reliably made prior to the start of recording. Trials in which subjects did not comply with the instruction and made corrections (Fig. 1C) were identified by inspection of the phase diagram after each unloading trial and those trials in which subjects intervened (15–20% of all trials) were excluded and repeated. In addition, in the majority of cases, intermediate positions after partial unloading were significantly different from each other suggesting that there was a unique relationship between final position and load. This implies that the change in the load, rather than other factors such as voluntary corrections or anticipatory responses to unloading, determined the set of steady state postures observed after unloading.
Invariant characteristics
Although individuals with stroke had lower initial torques compared to healthy subjects, (Table 1), partial removal of the load resulted in a significant number of distinct final hand positions (Fig. 5) associated with unique shoulder-elbow configurations and joint torques. Both groups were also able to adapt their control variables and specify different ICs related to the two different initial conditions (PUSH or PULL). In 4 out of 12 individuals with stroke, however, ICs could not be identified, implying that these individuals were unable to consistently specify referent arm configurations. This inability was not related to the severity of the arm paresis since their arm impairments ranged from moderate to severe and other individuals with more severe impairments did not have similar problems. In addition, this inability may not have been related to the use of low initial loads since the low-load control group working at similar levels of initial torque had no difficulties specifying referent configurations. The role of fatigue in the inability of these 4 individuals was also minimal since the loads they compensated were set well below their maximal voluntary efforts (30% MVF) and intertrial time was sufficient to prevent fatigue.
It is more likely that the inability to specify ICs may have been related to the increased instability and response variability in these 4 patients who had the highest and most variable instability indices of all the patients tested (see below). Similar to what has been previously shown in the single-joint system (elbow joint; Levin et al. 2000), the inability to adopt a stable final position after unloading in these patients was also accompanied by changed patterns of agonist-antagonist muscular coactivation at the elbow and shoulder joints. Problems in the specification of referent arm configurations in these individuals could not be predicted on the basis of their motor impairments measured by standard clinical tests. This implies that clinical testing is not sensitive to detect motor control deficits of this type. The testing of the ability to specify referent configurations may be more fundamental to the description of muscle activation patterns during coordinated movement than clinical tests, such as the Fugl-Meyer, that test whether or not a movement can be performed rather than how it is performed.
Referent arm configurations
The recording of two distinct R configurations for the two different initial conditions (PUSH and PULL) demonstrates the ability of the nervous system to modify control variables in a task-specific way. In other words, subjects were able to establish different R configurations for the same position of the hand to compensate different initial load directions. Once the appropriate R configuration was established, subjects were also able to maintain it thus allowing the neuromuscular system to generate automatic responses to unloading depending on the difference between the actual and the virtual, centrally specified R configuration of the arm.
The robustness of the R configurations was demonstrated by the high r2 values of the linear regressions fit to the individual joint IC surfaces in all healthy subjects regardless of the initial load level (Table 2). In most individuals with stroke, construction of the R configurations was also possible although the r2 values of the ICs were lower compared to the healthy group. It is unlikely that the poorer fit of the IC surfaces may have been related to difficulties in compensating interactive torques acting on one arm segment due to the motion of the other as reported by Beer et al. (2000). While such a deficit could be responsible for the increased variability and hook-like shape of hand trajectories during the dynamic phase of the movement (Fig. 3), it would have little influence on the final, steady state configurations of the arm. Since interactive torques are velocity- and acceleration-dependent, they are reduced to zero at the end of the movement and thus do not influence the final steady state. It is more likely that the poor fit of the IC in the four participants with stroke mirrors problems with the consistent specification of the R configuration in repeated trials.
Our data suggest that the variability of intermediate final positions in both groups of subjects was due to problems in the specification of R configurations and not due to fatigue for several reasons. First, efforts were made to minimize fatigue during data recording by allowing subjects adequate periods of rest. Second, all subjects had no difficulty maintaining initial force levels of 30% MVF throughout the experiment. Third, the mean final positions of trials recorded near the beginning and near the end of the recording session showed no evidence of increased variability in the majority of subjects.
It is also possible that deficits in the specification of R configurations in participants with hemiparesis may have been related to restrictions in their ability to produce movements in certain parts of the workspace. Following stroke, patients exhibit well-known pathological flexion and extension movement synergies at the early stages of recovery (Brunnström 1970; Twitchell 1951). In the arm, the extensor synergy is characterized by shoulder adduction and internal rotation, elbow extension, wrist pronation, and flexion. An opposite pattern of synergistic muscle activation is observed when attempts to move the arm evoke the flexor synergy. In our experiment, the movement in the PUSH direction could be considered as a movement made within the extensor synergy (shoulder adduction combined with elbow extension), whereas the movement in the PULL direction was not similar to either synergy (shoulder abduction combined with elbow extension). However, movements in the individuals participating in this study were only moderately to mildly impaired and they did not differ from those of healthy subjects making it unlikely that they were influenced by pathological synergies.
Alternatively, the deficits in the specification of R configurations may be related to the empirically observed limitations in the regulation of muscle activation thresholds that, in turn, restrict the range of shifts in the torque/angle characteristics, as revealed for elbow flexors and extensors in individuals with hemiparesis. Levin et al. (2000) showed that for the elbow joint, even patients with severe motor impairment could control elbow flexion and extension movements but this control was only achieved within a reduced angular range defined by flexor and extensor activation threshold angles. This angular range was described as a ‘reciprocal zone’ within which movement control resembled that in healthy subjects producing reciprocal activation of elbow flexors and extensors. Movements attempted to positions outside of this zone were accompanied by excessive coactivation. Projecting these observations to the multijoint system, Levin et al. (2002) suggested that deficits in the regulation of muscle activation thresholds of adjacent joints would restrict the range of available R configurations and constrain these joints to act together in specific areas of the workspace, thus providing an explanation for the appearance of abnormal movement synergies. The limited range of R configurations may, therefore, be a consequence of deficits in the specification of thresholds of muscle activation in mono- and biarticular muscles which together determine the R configurations (Feldman and Levin 1995). Future studies may be designed to address this specific issue.
Instability
Healthy subjects using moderate or low initial torques had stable end postures (low instability indices) for both PUSH and PULL. The amount of coactivation varied with the direction of voluntary effort. For PUSH healthy subjects had greater coactivation at the elbow than at the shoulder, while for PULL there was more coactivation at the shoulder than at the elbow (Table 5). In marked contrast, individuals with stroke had much greater endpoint instability. While the pattern of muscle coactivation at the elbow and shoulder was preserved for the PUSH direction, individuals with stroke used significantly more coactivation at the elbow and less coactivation at the shoulder for the PULL direction. In addition, in individuals with stroke, the amount of instability for PULL and PUSH was related to a change in the amount of coactivation at the elbow and shoulder and this deficit was related to the severity of the arm motor impairment.
The fact that both groups of healthy subjects had similar indices of instability suggests that the increased arm instability in individuals with stroke was not due to the use of lower initial torques. On the other hand, the results are consistent with several previous studies reporting increased arm postural instability in individuals with stroke (Beer et al. 2000; Levin and Dimov 1997).
In healthy subjects, stability of the single- and multijoint arm systems is related to the interdependence of the elbow and shoulder torques and the ability to specify appropriate agonist/antagonist muscle coactivation (Archambault et al. 2001; Franklin and Milner 2003; Gomi and Osu 1998; Hogan 1985; McIntyre et al. 1996; Milner 2002). Beer et al. (2000) used inverse dynamics to compute changes in the elbow/shoulder interaction torques during active arm movements in individuals with hemiparesis. They attributed inaccuracies of limb positioning not to such factors as weakness, spasticity, or stereotypic movements but to problems in the control of interactive torques. In our unloading experiments, neither these factors nor the control of interactive torques themselves play a major role in the specification of end positions in individuals with stroke, although problems in the control of interactive torques could partly be responsible for movement instability (terminal oscillations; see above). In addition, instability may be related to the deficits in the specification and regulation of coactivation as implied by the observations of disruptions in the specification of coactivation at the elbow (or the specification of arm stiffness) as well as by the correlation between abnormal coactivation and the instability indices. According to the EP hypothesis, in addition to stiffness regulation, coactivation commands have two important features. First, healthy subjects can regulate coactivation independently of position (Levin et al. 1995); one can easily coactivate arm muscles while maintaining the same arm posture. In contrast, many individuals with hemiparesis have problems producing coactivation without influencing arm position (Levin et al. 2000). Such deficient coactivation might be responsible for increased variability in the specification of the R configuration in these individuals. Second, according to the λ version of the EP hypothesis, coactivation has an important spatial characteristic—by specifying muscle activation thresholds, neural control levels predetermine where, in the arm workspace, muscles of opposing muscle groups can be coactive (‘coactivation zones’; see Feldman and Levin 1995). It has been shown that adults with stroke have problems in the task-related specification of the localization and extent of coactivation zones in the arm workspace and that this deficiency results in movement instability—prolonged terminal oscillations (Levin and Dimov 1997), similar to those found in the present study.
Thus, consideration of motor deficits of the double-joint arm system in terms of the EP hypothesis allows us to identify deficits in three basic control processes in individuals with stroke: a decreased range of specification of R configurations, problems with the stable reproduction of R configurations, and problems in producing functionally appropriate coactivation. These deficits may help to explain the increased instability and response variability observed when some patients attempt to make voluntary movements.
References
Archambault PS, Mihaltchev P, Levin MF, Feldman AG (2001) Parametric control of the arm analysed by unloading using a double-joint manipulandum. 31st Annual Meeting Soc Neurosci Abstracts 941.11, San Diego, CA
Asatryan DG, Feldman AG (1965) Functional tuning of the nervous system with control of movements or maintenance of a steady posture. I. Mechanographic analysis of the work of the joint on execution of a postural tasks Biophysics 10:925–935
Beer RF, Dewald JPA, Rymer WZ (2000) Deficits in the coordination of multijoint arm movements in patients with hemiparesis: evidence for disturbed control of limb dynamics. Exp Brain Res 131:305–319
Bohannon RW, Larkin PA, Smith MB, Horton MG (1987) Relationship between static muscle strength deficits and spasticity in stroke patients with hemiparesis. Phys Ther 67:1068–1071
Bourbonnais D, Vanden Noven S (1989) Weakness in patients with hemiparesis. Am J Occup Ther 43:313–317
Brunnström S (1970) Movement therapy in hemiplegia. A neurophysiological approach. Harper and Row, New York
Burke D (1988) Spasticity as an adaptation to pyramidal tract injury. Adv Neurol 7:401–423
Chen R, Cohen LG, Hallett M (1997) Role of the ipsilateral motor cortex in voluntary movement. Can J Neurol Sci 24:284–291
Cirstea MC, Levin MF (2000) Compensatory strategies for reaching in stroke. Brain 123:940–953
Cirstea MC, Mitnitski AB, Feldman AG, Levin MF (2003) Interjoint coordination dynamics during reaching in stroke. Exp Brain Res 151:289–300
Colebatch JG, Gandevia SC, Spira PJ (1986) Voluntary muscle strength in hemiparesis: distribution of weakness at the elbow. J Neurol Neurosurg Psychiatry 49:1019–1024
Conrad B, Benecke R, Meinck HM (1985) Gait disturbances in paraspastic patients. In: Delwaide PJ, Young RR (eds) Restorative neurology. Clinical neurophysiology in spasticity, vol 1. Elsevier, Amsterdam, pp 155–174
Coté JN, Mathieu PA, Levin MF, Feldman AG (2002) Movement reorganization to compensate for fatigue during sawing. Exp Brain Res 146:394–398
Crago PE, Houk JC, Hasan Z (1976) Regulatory action of human stretch reflex. J Neurophysiol 39:925–935
Dewald JP, Pope PS, Given JD, Buchanan TS, Rymer WZ (1995) Abnormal muscle coactivation patterns during isometric torque generation at the elbow and shoulder in hemiparetic subjects. Brain 118:495–510
Dickstein R, Hocherman S, Amdor G, Pillar T (1993) Reaction and movement times in patients with hemiparesis for unilateral and bilateral elbow flexion. Phys Ther 73:374–385
Feldman AG (1966) Functional tuning of the nervous system with control of movement or maintenance of a steady posture. II. Controllable parameters of the muscle. Biophysics 11:565–578
Feldman AG (1979) Central and reflex mechanisms in the control of movement. Nauka, Moscow
Feldman AG, Levin MF (1995) The origin and use of positional frames of reference in motor control. Behav Brain Sci 18:723–744
Feldman AG, Orlovsky GN (1972) The influence of different descending systems on the tonic stretch reflex in the cat. Exp Neurol 37:481–494
Feldman AG, Levin MF, Mitnitski A, Archambault P (1998) Multi-muscle control in human movements. Keynote lecture. J Electromyogr Kinesiol 8:383–390
Forget R, Lamarre Y (1987) Rapid elbow flexion in the absence of proprioceptive and cutaneous feedback. Hum Neurobiol 6:27–37
Franklin DW, Milner TE (2003) Adaptive control of stiffness to stabilise hand position with large loads. Exp Brain Res 152:211–220
Fugl-Meyer AR, Jääskö L, Leyman I, Olsson S, Steglind S (1975) The post-stroke hemiplegic patient. 1. A method for evaluation of physical performance. Scand J Rehabil Med 7:13–31
Gomi H, Osu R (1998) Task-dependent viscoelasticity of human multijoint arm and its spatial characteristics for interaction with environments. J Neurosci 18:8965–8978
Goulet C, Arsenault AB, Bourbonnais D, Laramee MT, Lepage Y (1996) Effects of transcutaneous electrical nerve stimulation on H-reflex and spinal spasticity. Scand J Rehabil Med 28:169–176
Gowland C, deBruin H, Basmajian JV, Plews N, Burcea I (1992) Agonist and antagonist activity during voluntary upper-limb movement in patients with stroke. Phys Ther 72:624–633
Hammond MC, Fitts SS, Kraft GH, Nutter PB, Trotter MJ, Robinson LM (1988) Co-contraction in the hemiparetic forearm: quantitative EMG evaluation. Arch Phys Med Rehabil 69:348–351
Hogan N (1985) The mechanics of multi-joint posture and movement control. Biol Cybern 52:315–331
Hufschmidt A, Mauritz K-H (1985) Chronic transformation of muscle in spasticity: a peripheral contribution to increased tone. J Neurol Neurosurg Psychiatry 48:676–685
Jakobsson F, Grimby L, Edstrom L (1992) Motoneuron activity and muscle fibre type composition in hemiparesis. Scand J Rehabil Med 24:115–119
Kamper DG, McKenna-Cole AN, Kahn LE, Reinkensmeyer DJ (2002) Alterations in reaching after stroke and their relation to movement direction and impairment severity. Arch Phys Med Rehabil 83:702–707
Knutsson E, Richards C (1979) Different types of disturbed motor control in gait of hemiparetic patients. Brain 102:405–430
Lestienne FG, Thullier F, Archambault P, Levin MF, Feldman AG (2000) Multi-muscle control of head movements in monkeys: the referent configuration hypothesis. Neurosci Lett 283:65–68
Levin MF (1996) Interjoint coordination during pointing movements is disrupted in spastic hemiparesis. Brain 119:281–293
Levin MF, Dimov M (1997) Spatial zones for muscle coactivation and the control of postural stability. Brain Res 757:43–59
Levin MF, Feldman AG (1994) The role of stretch reflex threshold regulation in normal and impaired motor control. Brain Res 657:23–30
Levin MF, Hui-Chan CW (1992) Relief of hemiparetic spasticity by TENS is associated with improvement in reflex and voluntary motor functions. Electroencephalogr Clin Neurophysiol 85:131–142
Levin MF, Lamarre Y, Feldman AG (1995) Control variables and proprioceptive feedback in fast single-joint movement. Can J Physiol Pharmacol 73:316–330
Levin MF, Selles RW, Verheul MH, Meijer OG (2000) Deficits in the coordination of agonist and antagonist muscles in stroke patients: implications for normal motor control. Brain Res 853:352–369
Levin MF, Cirstea CM, Archambault P, Son F, Roby-Brami A (2002) Impairment and compensation of reaching in patients with stroke and cerebral palsy. In: Latash ML (ed) Progress in motor control, vol 2. Structure-function relations in voluntary movements. Human Kinetics, Champaign, IL, pp 103–122
Matthews PBC (1959) A study of certain factors influencing the stretch reflex of the decerebrate cat. J Physiol 147:547–564
McIntyre J, Mussa-Ivaldi FA, Bizzi E (1996) The control of stable postures in multijoint arm. Exp Brain Res 110:248–264
Michaelsen SM, Luta A, Roby-Brami A, Levin MF (2001) Effect of trunk restraint on the recovery of reaching movements in hemiparetic patients. Stroke 32:1875–1883
Mihaltchev P, Archambault PS, Feldman AG, Levin MF (2002) Referent configuration of the arm in patients with stroke. Program no. 169.6, Abstract Viewer/Itinerary Planner, Washington DC, Soc for Neurosci, CD-ROM
Milner TE (2002) Adaptation to destabilising dynamics by means of muscle cocontraction. Exp Brain Res 143:406–416
Nadeau S, Arsenault AB, Gravel D, Bourbonnais D (1999) Analysis of the clinical factors determining natural and maximal gait speeds in adults with a stroke. Am J Phys Med Rehabil 78:123–130
Newell KM, Houk JC (1983) Speed and accuracy of compensatory responses to limb disturbance. J Exp Psychol 9:58–74
Ostry DJ, Gribble PL, Levin MF, Feldman AG (1997) Phasic and tonic stretch reflexes in muscles with few muscle spindles: human jaw-opener muscles. Exp Brain Res 116:299–308
Reinkensmeyer DJ, McKenna Cole A, Kahn LE, Kamper DG (2002) Directional control of reaching is preserved following mild/moderate stroke and stochastically constrained following severe stroke. Exp Brain Res 143:525–530
Roby-Brami A, Fuchs S, Mokhtari M, Bussel B (1997) Reaching and grasping strategies in hemiparetic patients. Motor Control 1:72–91
St-Onge N, Feldman AG (2004) Referent configuration of the body: a global factor in the control of multiple skeletal muscles. Exp Brain Res 155:291–300
Tang A, Rymer WZ (1981) Abnormal force-EMG relations in paretic limbs of hemiparetic human subjects. J Neurol Neurosurg Psychiatry 44:690–698
Trombly CA (1993) Observations of improvement of reaching in five subjects with left hemiparesis. J Neurol Neurosurg Psychiatry 56:40–45
Twitchell TE (1951) The restoration of motor function following hemiplegia in man. Brain 74:443–480
Weijs WA, Sugimura T, van Ruijven LJ (1999) Motor coordination in a multi-muscle system as revealed by principal components analysis of electromyographic variation. Exp Brain Res 127:233–243
Wiesendanger M (1991) Neurophysiological basis of spasticity. In: Sindou M, Abbott R, Keravel Y (eds) Neurosurgery for spasticity. Springer, Berlin Heidelberg New York, pp 15–19
Wing AM, Lough S, Turton A, Fraser C, Jenner JR (1990) Recovery of elbow function in voluntary positioning of the hand following hemiplegia due to stroke. J Neurol Neurosurg Psychiatry 53:126–134
Winstein CJ, Pohl PS (1995) Effects of unilateral brain damage on the control of goal-directed hand movements. Exp Brain Res 105:163–174
Yanagisawa N, Tanaka R (1978) Reciprocal Ia inhibition in spastic paralysis in man. In: Cobb WA, van Duijn H (eds) Contemporary clinical neurophysiology, EEG suppl 34. Elsevier, Amsterdam, pp 521–526
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Mihaltchev, P., Archambault, P.S., Feldman, A.G. et al. Control of double-joint arm posture in adults with unilateral brain damage. Exp Brain Res 163, 468–486 (2005). https://doi.org/10.1007/s00221-004-2202-9
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00221-004-2202-9