
Abstract
Participants with normal (StereoN) and weak (StereoW) stereopsis caught tennis balls under monocular and binocular viewing at three different speed conditions. Monocular or binocular viewing did not affect catching performance in catchers with weak stereopsis, while the StereoN group caught more balls under binocular vision as compared with the monocular condition. These effects were more pronounced with increasing ball speed. Kinematic analysis of the catch partially corroborated these findings. These results indicate that StereoW catchers have not developed a compensatory strategy for information pick-up, and that negative effects of a lack of stereopsis grow larger as temporal constraints become more severe. These findings also support the notion that several monocular and/or binocular information sources can be used in the control of interceptive action.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Catching a ball with one hand seems to be an easy thing to do. For successful interception, however, the catcher needs exact information about the spatio-temporal characteristics of the ball in flight. In particular the timing of the grasp, i.e. closing of the hand, must be very accurate if a successful catch is to be made (Alderson et al. 1974). The so called time-to-contact (ttc), i.e. the time remaining until the ball will arrive at the catchers hand, is considered as an important variable to guide the temporal control of interceptive actions. Monocular as well as binocular information sources for generating ttc information have already been identified. Lee (1976) suggested an optical variable tau that directly specifies ttc as the inverse of relative rate of the retinal expansion of the oncoming object generated in the optic array. Tau is monocularly available and has been shown to be a plausible informational candidate for the control of the temporal aspects of the grasp (Savelsbergh et al. 1991; Savelsbergh and Whiting 1992). However, tau alone does not seem to be sufficient. Several authors have demonstrated in both theoretical (Laurent et al. 1996) and experimental ways (Rushton and Wann 1999; van der Kamp et al. 1999) that binocular information sources such as retinal disparity, and more specifically the tau function of disparity, can be involved as well. In order to unravel which of these visual information sources, tau or disparity, is used in the timing of interceptive actions, visual conditions in people with normal visual abilities have been artificially manipulated in different ways.
First, Judge and Bradford (1988), van der Kamp et al. (1999) and Bennett et al. (1999b, 2000) investigated to what extent binocular information is involved in ttc estimation by using a telestereoscope. Two pairs of parallel mirrors in the telestereoscope lead to the illusion that an object is closer to the observer than it really is, without affecting the monocular information from both eyes separately. When wearing this device, a decrease in catching performance under binocular but not under monocular viewing was found (Judge and Bradford 1988; Bennett et al. 1999b, 2000; van der Kamp et al. 1999). These findings indicate that multiple sources of monocular and binocular information may contribute to the timing of a catch. Similar results were obtained by Gray and Regan (1998), who showed in several psychophysical experiments that more accurate ttc judgements were made when both monocular and binocular information was available. Second, Rushton and Wann (1999) applied virtual reality to create a similar misjudgement: participants had to catch virtual balls of which the diameter was computationally scaled, so that the ball appeared to arrive 100 ms earlier or later than specified by retinal disparity. They demonstrated that binocular disparity and monocular tau are both used for the timing of a catch, the relative weight of both cues depending on which of these information sources specifies the earliest arrival. A third paradigm is to compare catching behaviour in monocular and binocular circumstances, since disparity cannot be used under monocular viewing. Van der Kamp et al. (1997) and Savelsbergh and Whiting (1992) both found a superior catching performance when catching binocularly compared with monocular catching, although they did not agree whether the weaker monocular performance was a result of temporal or spatial errors. A limitation of these monocular-binocular experiments is that the monocular condition was created by covering one eye, which results in more than loss of retinal disparity alone: additional binocular information sources such as concordant information and ocular vergence, but also monocular information sources such as the optical variable tau, relative size of familiar objects, image velocity and motion parallax from the second eye are no longer available (Lee 1976; Collewijn and Erkelens 1990; Davson 1990).
The general conclusion from these experiments is that, depending on the specific circumstances, monocular (e.g. tau) and binocular (e.g. disparity) information sources, or a combination of both, are relied on to accomplish successful interception.
However, in all these paradigms the visual condition of healthy participants was artificially manipulated, creating an unnatural and new situation in which they had minimal experience. Since retinal disparity is the primary cue for binocular depth perception or stereopsis (Collewijn and Erkelens 1990; Patterson and Martin 1992), studying individuals suffering from detrimental stereoscopic vision would entail a unique population to assess the contribution of stereopsis and retinal disparity in the timing of interceptive actions. Moreover, in what way a person with poor stereoscopic vision (disparity) acts in an inexperienced monocular situation has not yet been examined. This could be interesting to find out whether people suffering from weak stereopsis have adapted to their particular visual condition by relying on other monocular information sources in making their ttc estimation. It is plausible that they somehow have compensated for their lack of stereoscopic vision.
Lenoir et al. (1999) examined one-handed binocular catching of individuals with either good stereopsis or congenital/early-onset deficiency in stereopsis. It appeared that those who had weak stereopsis made significant more temporal errors, that is, they closed the hand too late compared to the control group with good steropsis. So, a deficiency in stereopsis turned out to be detrimental for interceptive performance, in particular the temporal aspect of it. These results corroborate the notion that tau and disparity both play a part in the timing of a catch.
The aim of the present experiment was to examine both monocular and binocular performance in a natural catching task in people with good and weak stereopsis. Since all participants were inexperienced in the monocular test condition, they are expected to perform better binocularly than in the monocular test situation, which would be in line with the findings of Savelsbergh and Whiting (1992). However, if stereopsis or disparity is important in the timing of a catch, it is expected that catchers with good stereopsis will benefit more from binocular vision than participants with poor stereopsis, unless the latter have developed some kind of compensatory mechanism for dealing with their weaker stereoacuity. This unique set-up might bring further insight onto the issue of what binocular and monocular information sources are used in the timing of interceptive movements. In addition, having the participants catch balls at different velocities could also clear up whether stereopsis and/or disparity is more important in real-life situations or if it rather relates to conditions with high temporal constraints.
Methods
Participants
Two groups of nine students in physical education (three males and six females in each group) between 18 and 23 years of age with normal or corrected-to-normal vision participated in this experiment after giving informed consent. Participants were selected from a large group of students according to their performance on the Graded Circle Test (from the “Random Dot Stereo Butterfly” test battery—Stereo Optical Company Inc., Chicago, IL, USA). In the StereoN group, all participants had very good stereoacuity (40 arc s or better); the participants of the StereoW group all had very weak stereopsis (five participants scored 400 arc s, four could not see any of the images at all, i.e. worse than 800 arc s). Stereoacuity measures of 30–40 arc s were reported as normal by Fielder and Moseley (1996). All but one of the participants were right-handed catchers. As students in physical education, all participants had experience in ball games such as basketball, volleyball, handball, soccer and tennis. All participants were naive to the purpose or hypotheses of the experiment and were paid a small fee for their collaboration. The study was approved by the ethics committee of the University Hospital Ghent.
Apparatus
A Singly Promatch ball-projection machine (Promatch / Mubo B.V., Gorinchem, The Netherlands) with adjustable launching speed and angle projected yellow tennis balls toward the upright-standing participant. The balls were projected at three different velocities: 8.4, 11.6 and 14.6 m/s, from a distance of 8.40 m, resulting in flight times of the ball of 1001 (±11 ms), 722 (±4 ms) and 573 ms (±5 ms), respectively. Since the focus of this experiment was on the timing aspects of the catch, the spatial complexity of the task was reduced by a) having the balls arrive within an imaginary circle (30 cm in diameter) with the centre of it approximately 10 cm above the catcher’s shoulder, and b) having the participants start with the catching hand near this centre: in front of the shoulder, thumb and index finger together.
All trials were recorded with a Sony CCD-TRV94E Hi8 camera (25 Hz) from the medial side of the catching arm. Three-dimensional kinematics of the first ten trials from each condition were also recorded by means of a 7-camera infrared recording system (Proreflex MCU240 Qualisys, Sweden) at 240 Hz. Therefore, reflective markers were attached with double-sided adhesive cloth tape to the external face of the distal phalanx of thumb and index finger, caput metacarpale of pollux, index and digitus minimus, and to the processus styloideus of radius and ulna. The obtained 3D data were subsequently filtered at a cut-off frequency of 10 Hz with a second-order recursive Butterworth filter.
Design and procedure
All participants were tested in two sessions with a 1-week interval. One session was executed under a normal binocular viewing condition, while in the other session the balls had to be caught monocularly. The monocular-binocular order was randomised over participants. In the monocular condition, they caught with their preferred eye, while the other eye was covered with an eye patch.
In each session, the participants had to catch 90 tennis balls with their preferred hand, in three blocks of 30 for each velocity condition. The sequence of ball speeds was randomised for all sessions to exclude effects of presentation order. Before each block of 30 balls, ten acclimatisation trials were provided. The first ten catches in every session were recorded with both the Qualisys infrared motion capture system and a 25-Hz Sony Hi8 camera. The subsequent 20 trials were captured with only the 25-Hz Sony Hi8 camera. After a successful catch, the ball was dropped into a basket and the hand returned to the initial position. In case of a failure, the hand was returned into the initial position straight away. The outcome of each trial was immediately registered by one of the researchers as a catch or miss.
Dependent variables
In order to examine general catching behaviour, the percentage of successful catches was analysed. Additionally, three sets of kinematic variables were included in the analysis: temporal variables, spatial variables and velocity variables. Ball-hand contact was determined by the sudden backward acceleration of the metacarpal and finger markers at the time of impact. Hand aperture and velocity, displacement and velocity of the wrist were computed from the positional data, and the required variables extracted. All of the temporal variables were defined with respect to ball-hand contact (Tc).
Temporal variables
-
Movement time (MT, millliseconds): time elapsing from the occurrence of the first hand movement until ball-hand contact
-
Latency time (LT, millliseconds): time elapsing between the launching of the ball from the ball machine and the first hand movement, calculated as flight time of the ball minus the movement time
-
Moment of hand closure (Tcl, millliseconds): time at which hand aperture reached its maximum and the hand started to close (i.e. the moment the hand opening velocity turns negative)
Spatial variables
-
Peak hand aperture (PeakHa, millimetres): since the catch is initiated with thumb and index finger united, this variable specifies also the amount of hand opening
-
Hand aperture at the moment of ball-hand contact (HaTc, millimetres)
-
Amount of hand closure (Hcl, millimetres): hand aperture on Tcl minus hand aperture on Tc
-
Coefficient of straightness (CoStr, %): defined as the actual travelled path of the wrist divided by the distance of the shortest linear path × 100, specifying the rectilinearity of the wrist trajectory. Note: this variable is the inverse of the coefficient of straightness defined by Laurent et al. (1994), i.e. shortest path divided by real path ×100
-
Forward displacement of the wrist (DxWrist, millimetres): linear distance between the position of the wrist at the initiation of the catching movement and ball-hand contact along the sagittal axis (X-axis)
Dynamic variables
-
Peak hand opening velocity (PeakHoV, millimetres per second)
-
Peak hand closing velocity (PeakHcV, millimetres per second)
Data analysis
First a 3 (ball speed: low (8.4 m/s) vs. medium (11.6 m/s) vs. high speed (14.6 m/s)) × 2 (visual conditions: monocular vs. binocular) × 2 (stereoacuity group: StereoW vs. StereoN) × 2 (session order: mono-bino vs. bino-mono) ANOVA analysis of variance with repeated measures on the first and second factor was executed on catching performance. Because presentation order of monocular and binocular conditions did not show any significant main effect or interaction, this factor was eliminated from any further analysis.
A 2 (stereoacuity group: StereoW vs. StereoN) × 2 (visual conditions: monocular vs. binocular) × 3 (velocities: low vs. medium vs. high speed) ANOVA analysis of variance with repeated measures on the last two factors was carried out on the success percentages and on the kinematic variables. All Qualisys recorded trials, successful catches as well as failures, were included in the analysis of kinematic data.
Post hoc comparisons were conducted with LSD test in order to examine the nature of the main and interaction effects. An alpha level of .05 was used for all statistical tests and the size of the effect was provided by the partial Eta squared (ηp 2).
Results
General catching performance
A main velocity effect on percentage of successful catches was found (F (2,32)=62.258, p<.05, ηp 2=.796). Catching performance decreased with increasing ball speed. At the lowest speed 91.36% of all balls were caught, while 79.62% were caught at moderate speed and 57.59% at the highest speed. Post hoc analysis revealed significant differences between all three ball speeds (p<.05). Participants performed better binocularly than in the monocular viewing condition (F (1,16)=34.959, p<.05, ηp 2=.686). While participants succeeded in 83.48% of 90 trials in the binocular viewing condition, under monocular conditions the success rate was 68.90%. No main effect of stereoacuity on catching performance was found (F (1,16)=1.139, ns). However, important interactions with other factors deserve attention.
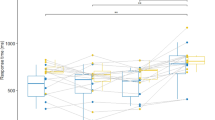
First, a significant interaction between stereoacuity and visual condition (F (1,16)=11.997, p<.05, ηp 2=.429) occurred. Further analysis of this interaction showed an increase in performance for the StereoN group for the binocular condition in comparison with the monocular catching task (23.12% gain, p<.05). In the StereoW group success rate was similar in monocular and binocular conditions (a non-significant gain of 6.04%). Put in an alternative way, both groups performed equally in the monocular viewing condition (69.24% for StereoN vs. 68.56% for StereoW, ns), while a very strong tendency was present for the StereoN group to outperform the StereoW group under binocular vision (92.35% for StereoN vs. 74.60% for StereoW, p=.057). Second, a significant interaction effect between ball speed and visual condition was present. The difference between monocular and binocular performance grew significantly larger with increasing ball speed (F (2,32)=4.750, p<.05, ηp 2=.229). The gain in performance when catching binocularly compared with the monocular condition was 7.56%, 13.94% and 22.23% for the low, moderate and high ball speed, respectively. Finally, the significant speed × group × visual condition-interaction (Fig. 1) indicated that the increasing benefit of binocular vision with increasing ball speed only holds for participants with good stereoscopic vision (F (2,32)=4.470, p<.05, ηp 2=.218). The monocular-binocular gain for the StereoW group remains small and non-significant: 5.9%, 6.0% and 6.3% at low, moderate and high ball speeds, while a benefit of 9.3% (ns), 21.9% (p<.05) and 38.2% (p<.05), respectively, for the StereoN group was found. See Table 1.

Effect of ball speed × stereoacuity × visual condition on % of successful catches
Kinematic variables (Table 1)
Temporal variables
No effects for moment of hand closure were found, i.e. duration of the grasping phase (grasping time) was equal for all conditions, irrespective of ball speed (F (2,32)=1.383, ns), visual condition (F (1,16)=2.383, ns) or stereoacuity (F (1,16)=0.274, ns). A main speed effect for movement time and latency time occurred (F (2,32)=89.784, p<.05, ηp 2=.849 for MT and F (2,32)=39.893, p<.05, ηp 2=.827 for LT). Both MT and LT decreased with increasing ball speed. Post hoc comparisons revealed significant differences between all ball speed conditions (p<.05).
Spatial variables
Participants with good stereopsis tended to execute a larger grasp (i.e. the hand closed more) compared with the participants with weak stereopsis, though this effect failed to attain statistical significance (F (1,16)=3.926, p=.065, ηp 2=.197). A main effect of visual condition on DxWrist was found (F (1,16)=5.556, p<.05, ηp 2=.258), i.e. when catching monocularly the participant intercepted the ball closer to the body compared with binocular catching.
An interaction effect between stereoacuity and visual condition on the coefficient of straightness was found (F (1,16)=4.362, p<.05, ηp 2=.214). For the StereoN group, the wrist appeared to travel less straight toward the point of ball-hand contact when catching binocularly as compared with the monocular catching condition (CoStr of 109.0 vs. 106.7% in binocular and monocular visual conditions, respectively). For StereoW, no difference between both conditions seems to occur (CoStr of 105.3 vs. 106.3% for binocular and monocular vision, respectively).
Peak hand aperture increased with increasing ball speed (F (2,32)=4.375, p<.05, ηp 2=.215), PeakHa being significantly larger at the highest velocity as compared with the two lower velocities. A main speed effect for hand aperture on moment of ball-hand contact occurred as well; HaTc was larger with increasing ball speed (F (2,32)=21.372, p<.05, ηp 2=.572). Post hoc analysis revealed significant differences among all three speed conditions (p<.05). A significant speed effect on the amount of hand closure was found (F (2,32)=16.346, p<.05, ηp 2=.505); the hand closed significantly less as ball speed increased.
Dynamic variables
Analysis of the velocity profile of the catch showed a significant difference between both stereo groups for peak hand closing velocity (F (1,16)=5.219, p<.05, ηp 2=.246); participants with good stereoacuity showed significantly higher PeakHcV values than participants of the StereoW group (p<.05). In addition, a speed effect was found for PeakHcV (F (2,32)=17.658, p<.05, ηp 2=.525). PeakHcV was significantly higher in the fastest speed condition compared with the slower ball speed conditions (p<.05). Also for peak hand opening velocity a speed effect was present (F (2,32)=62.804, p>.05, ηp 2=.797). PeakHoV increased significantly with increasing ball speed (p<.05). No interactions with visual condition occurred for PeakHoV or for PeakHcV.
Discussion
The aim of this study was to examine catching behaviour under monocular and binocular viewing in people with good and weak stereopsis. Catchers with good stereopsis showed an increase in catching performance when catching binocularly compared with monocular viewing, a phenomenon that was not observed in the StereoW group. Participants with good stereoacuity closed their hand more during the grasp, and showed larger peak closing velocities. Larger coefficients of straightness were observed in the StereoN group under binocular vision than under monocular viewing, while no differences in the StereoW group were found.
General catching performance
The main finding in this study, the significant interaction between visual condition and stereoacuity group on number of successful catches, provides an answer to several questions concerning both the information sources in catching and implications for daily life.
The addition of a second viewpoint does not add much to the quality of the catch in people with very weak stereopsis. Under normal binocular viewing, individuals with poor stereoscopic vision and inadequate retinal disparity could also use concordant information or convergence next to tau (present in both eyes) to attune their catch (Jones and Lee 1981). Given that binocular performance did not exceed monocular catching, they did not take advantage of this additional available information. Whatever experimental condition, participants with weak stereopsis seemed to depend only on monocularly available information (e.g. tau) to time their catch. By consequence, the advantage of binocular vision apart from stereopsis or disparity is very limited if not negligible, which confirms earlier statements that the major advantage of having two eyes seems to be stereoscopic depth discrimination (Fielder and Moseley 1996) and/or retinal disparity. This is, however, in contrast with a series of experiments by Jones and Lee (1981), which showed that concordant or matching information from both eyes is a significant advantage of binocular vision. In this respect it has to be noticed that Jones and Lee (1981) did not use interceptive actions in their experiments, which could explain the discrepancy in results. Other work by Lee et al. (1991, 1992), Heuer (1993) and Laurent et al. (1996) suggested that a tau function (i.e. x/(dx/dt) of some binocular variable x, and more specific the tau function of not only disparity, but vergence as well, could also play a role in interceptive timing. Although our participants from the StereoW group did dispose of two clear monocular views, the available concordant or vergence information did not add significantly (about 6%) to their performance. Therefore, it might be stated that the tau function of disparity is a valuable binocular information source for the temporal control of the catch.
The relationship between static stereoacuity and a dynamic task like catching confirms earlier findings (Lenoir et al. 1999). Nevertheless, this is somewhat surprising since different cortical mechanisms have been identified for the processing of static and dynamic disparity (Cynader and Regan 1978; Regan and Cynader 1982; Regan 1997). A person with normal static stereoacuity can be blind to a rate of change of relative disparity or vice versa. What could explain the weaker binocular catching performance if dynamic disparity were still available in the StereoW group remains obscure. In fact, this might be a redundant question. Support for these different mechanisms mainly stems from psychophysical experiments, in which no motor response guided by the available information is given. However, there is evidence for separate information processing channels for visual perception on the one hand, and for visual control of goal-directed action on the other hand (Milner and Goodale 1995). It is therefore not clear at this time to what extent results from psychophysical experiments may be generalized to visually guided motor tasks. Although this issue deserves further elaboration in the future, it seems feasible that the static stereotest correlates with the dynamic catching performance, regardless of the sensitivity of the participant for dynamic disparity.
The significant difference between the StereoW and StereoN group in binocular circumstances, which was not present when catching monocularly (69.24% for StereoN vs. 68.56% for StereoW in monocular catching), clearly demonstrates that people with a lifelong experience with virtually no binocular depth vision are not able to compensate for this deficiency by depending more on other information sources (e.g. vergence, concordant information or tau from both monocular views), as was suggested by Lenoir et al. (1999). In binocular catching, they keep lagging behind the StereoN group by about 18%, which is not dramatic in daily life, but might become a problem when severe temporal constraints are imposed, such as in fast ball sports or in traffic situations. Similarly Steeves et al. (2000) showed that enucleated observers had not developed compensatory mechanisms for lacking binocular vision.
Jones and Lee (1981) suggested, however, that binocular stereopsis should only play a part when the head is stationary. Is the head free to move, depth information could arise from the optical flow in one eye. In the telestereoscope studies (Judge and Bradford 1988; Bennett et al. 2000) the head was stationary, while participants in our study were free to move the head. In spite of this freedom, stereopsis still accounted for almost 18% of the success rate for the StereoN group compared with StereoW, in binocular conditions.
The abovementioned equal monocular success rates for the StereoW and the StereoN group shows that monocular information is sufficient to obtain a score of nearly 70%. Monocular sources, such as the relative rate of optical dilatation of the approaching ball (tau), specifying time to contact, is sufficient for reasonable but not for maximal performance in the timing of the catch. The monocular success rates reveal that anyhow tau or other monocular information sources are used for the timing of the catch. The slowest ball velocity task even shows a pretty good performance in spite of the lack of any kind of binocular information for both groups (85.51% for StereoW and 89.64% for StereoN). When temporal constraints get rather severe, still 47.78 and 45.17% of all trails for StereoW and StereoN, respectively, are accomplished successfully with monocular information only. However, the ball speed × visual condition × stereoacuity interaction may lead to the suggestion that as the temporal constraints become more stringent, the importance of monocular information decreases in favour of binocular sources when the latter are available, as is the case in the StereoN group. This is expressed by the increasing monocular-binocular difference with increasing ball velocity (Fig. 1 and Table 1). An alternative hypothesis could be that when the temporal window decreases, the availability of redundant monocular and binocular sources, all specifying the same information, is favourable to reach a high level of performance. In this case the importance of combining several sources of information is emphasized for the control of fast interceptive movements.
The fact that the gain in successful catches for participants with good stereopsis (and thus good retinal disparity) under binocular viewing grows remarkably higher with increasing ball speed, suggests that a binocular source, in particular the tau function of disparity, is to be held responsible for this. This effect is in fact predicted by the motion in depth model put forward by Regan and Beverley (1979), which suggests that changing disparity as a stimulus for motion-in-depth sensation becomes more effective as velocity increases.
Kinematic analysis
Before examining the kinematics of the catching movement more profoundly, it is important to emphasize the fact that successful catches as well as failures were considered in the kinematic analysis. By consequence, relatively more failures are processed in these catching conditions where catching performance was weaker, e.g. in monocular catching vs. binocular or at the highest ball speed vs. slowest. But in this way, a representation of the general catching behaviour of both groups is reflected.
The differences in catching performance between both stereo groups were partially encountered again in the kinematics of the catching movement, more specifically in the grasping phase. The initial part of the catch was performed in a similar way by both stereo groups: movement time (time of onset of the movement prior to ball-hand contact), the magnitude of the maximal hand aperture and moment of the initiation of the grasp were equal. The grasp, however, showed important differences: participants with poor stereopsis not only closed their catching hand less compared with the participants with good stereopsis, they also reached a lower peak closing velocity of the hand, resulting in a weaker performance. These findings imply that the weaker catching performance of StereoW is not necessarily due to a too late initiation of the grasp (timing error), but rather a too slow execution of the closing movement of the fingers around the ball, i.e. the dynamical aspect of the catch. Apparently, the fine tuning of the hand closure to the approaching ball is not as adequate in the StereoW group as in the StereoN group. Corrections to the ongoing grasp are possible until the very last millisecond before ball-hand contact, based on the available information one visuo-motor delay earlier (funnel-like type of control by Bootsma et al. 1991). Since the StereoW participants have less redundant information available, the quality of the ongoing information stream for the StereoW group is probably lower, and therefore they might fail to make the meticulous adjustments essential for successful catching.
An important finding concerning the spatial aspect of the catch is the interaction between stereoacuity and visual condition on the coefficient of straightness. The stereo group × visual condition interaction that occurred in the overall catching performance was encountered again in the linearity of the trajectory of the wrist. Since participants with good stereoscopic abilities, when catching binocularly, had higher coefficients of straightness, they seem to implement more spatial corrections during their catching movement. A more precise orientation of the hand in space to obtain a successful catch is in this way a spatial advantage.
Contrary to the results of Laurent et al. (1994), in the present study the point of ball interception did not show a shift backwards. However, such an effect was present between visual conditions: when catching monocularly, participants intercepted the ball closer to the body. Apparently participants buy themselves extra time to cope with the perceptual restrictions of performing under the less familiar monocular vision.
The impact of the changing temporal constraints imposed to the catcher is also revealed in the kinematic data. First of all, a raise in peak hand aperture was found as ball speed increased. If the ball approaches faster, participants opened their catching hand wider. This could be a safety margin due to the fact that there is less time to execute potential necessary adjustments before initiating the grasp. This is in line with a reaching movement experiment by Wing et al. (1986), where a wider hand aperture was found when reaching faster than normal. Another interesting finding was that no speed effect occurred on the initiation of the grasp, the grasp being initiated at a constant time to contact. This is in line with the results of several studies (e.g. Savelsbergh et al. 1992; Laurent et al. 1994; Watson and Jakobson 1997; Bennett et al. (1999a). For peak closing velocity, however, a speed effect occurred, i.e. PeakHcV decreased with increasing ball speed. This is not surprising, given the facts that on the one hand participants initiated their grasp at a constant time to contact irrespective of the ball speed (invariant grasping time), and on the other hand the magnitude of the hand closure reduced. Since a raise in PeakHa occurred with increasing ball speed together with a reverse speed effect on PeakHcV, the hand does not close enough to get a firm grip on the ball. Thus, the ball bounces away from the area of the metacarpo-phalangeal joints when the fingers are still closing, resulting in a catching failure. The present experiment revealed that not really discrete temporal landmarks such as a late initiation of the hand closure are involved, but rather that the dynamical aspects of the grasp are responsible for the outcome of the catching movement, i.e. a successful catch or a failure.
In addition, spatial inaccuracy might be involved in the outcome of the catching movement. In spite of the fact that the experimental set-up was designed to more or less exclude spatial errors, they were still possible. The presence of such spatial inaccuracy may be reflected by the interaction effect between visual condition and stereoacuity on the coefficient of straightness. The fact that a difference in stereoacuity elicits both spatial and temporal kinetic differences suggests that the spatial and temporal aspects in the control of interceptive action are somewhat intertwined—as was the case in the work of Peper et al. (1994) and Montagne and Laurent (1994) where spatiotemporal aspects of the catching movement are discussed—and should not be approached as two separate entities.
It remains to be noticed that the natural task used in the present experiment is probably responsible for the large inter- and intra-personal variability that was present in the kinematic data. In that way, the experimental set-up might not be powerful enough to reveal distinct kinematics differences between both stereo groups to explain the observed differences in catching performance.
Conclusions
An important conclusion from this experiment is that people with weak stereopsis do not seem to have adapted for their lack of binocular depth vision by developing compensatory mechanisms. Whether they catch in normal binocular circumstances or with the vision of one eye only hardly affects catching success. In addition, it is suggested that poor stereopsis is not really disadvantageous in daily life, but only restricted to highly temporal constrained situations, such as in fast ball games. Kinematic analysis of the unrestricted natural catching movements revealed both spatial and temporal changes, i.e. differences in coefficient of straightness and grasping dynamics, respectively, in participants with weak stereopsis compared with those with good stereoscopic vision.
Another main finding of this study is that disparity is probably used in combination with a monocular time-to-contact information source, and that the importance of monocular and binocular sources depends on the conditions of execution (see also van der Kamp et al. 1997; Rushton and Wann 1999). It is suggested that in “normal circumstances” the perceptual process entails the use of several information sources at the same time and consequently the implementation of some kind of integrating mechanism, which may lead to a better quality of ttc information.
References
Alderson GJK, Sully DJ, Sully HG (1974) An operational analysis of a one-handed catching task using high speed photography. J Mot Behav 6:217–226
Bennett SJ, Davids K, Craig T (1999a) The effect of temporal and informational constraints on one-handed catching performance. Res Q Exerc Sport 70:206–211
Bennett SJ, van der Kamp J, Savelsbergh GJP, Davids K (1999b) Timing a one-handed catch. I. Effects of telestereoscopic viewing. Exp Brain Res 129:362–368
Bennett SJ, van der Kamp J, Savelsbergh GJP, Davids K (2000) Discriminating the role of binocular information in the timing of a one-handed catch. The effects of telestereoscopic viewing and ball size. Exp Brain Res 135:341–347
Bootsma RJ, Houbiers MHJ, Whiting HTA, van Wieringen PCW (1991) Acquiring an attacking fore-hand drive: the effects of static and dynamic environmental conditions. Res Q Exerc Sport 62:276–284
Collewijn H, Erkelens CJ (1990) Binocular eye movements and the perception of depth. In: Kowler E (ed) Eye movements and their role in visual and cognitive processes. Elsevier Science, Amsterdam, pp 213–261
Cynader M, Regan D (1978) Neurons in cat para-striate cortex sensitive to the direction of motion in three-dimensional space. J Physiol 274:549–569
Davson H (1990) Visual perception: introduction. In: Davson H (ed) Physiology of the eye. MacMillan, London, pp 449–485
Fielder AR, Moseley MJ (1996) Does stereopsis matter in humans? Eye 10:233–238
Gray R, Regan D (1998) Accuracy of estimating time to collision using binocular and monocular information. Vision Res 38:499–512
Heuer H (1993) Estimates of time to contact based on changing size and changing target vergence. Perception 22:549–563
Jones RK, Lee DN (1981) Why two eyes are better than one: the two views of binocular vision. J Exp Psychol Hum Percept Perform 7:30–40
Judge SJ, Bradford CM (1988) Adaptation to telestereoscopic viewing measured by one-handed ball-catching performance. Perception 17:783–802
Laurent M, Montagne G, Savelsbergh GJP (1994) The control and coordination of one-handed catching: the effect of temporal constraints. Exp Brain Res 101:314–322
Laurent M, Montagne G, Durey A (1996) Binocular invariants in interceptive tasks: a directed perception approach. Perception 25:1437–1450
Lee DN (1976) A theory of visual control of braking based on information about time-to-collision. Perception 5:437–459
Lee DN, Reddish PE, Rand DT (1991) Aerial docking by hummingbirds. Naturwissenschaften 78:526–527
Lee DN, Young DS, Rewt D (1992) How do somersaulters land on their feet? J Exp Psychol Hum Percept Perform 18:1195–1202
Lenoir M, Musch E, La Grange N (1999) Ecological relevance of stereopsis in one-handed ball-catching. Percept Mot Skills 89:495–508
Milner AD, Goodale MA (1995) The visual brain in action. Oxford University Press, Oxford
Montagne G, Laurent M (1994) The effects of environmental changes on one-handed catching. J Mot Behav 26:237–246
Patterson R, Martin WL (1992) Human stereopsis. Hum Factors 34:669–692
Peper L, Bootsma RJ, Mestre DR, Bakker FC (1994) Catching balls: how to get the hand to the right place at the right time. J Exp Psychol Hum Percept Perform 20:591–612
Regan D (1997) Visual factors in hitting and catching. J Sports Sci 15:533–558
Regan D, Beverley KI (1979) Binocular and monocular stimuli for motion in depth: changing-disparity and changing-size feed the same motion-in-depth stage. Vision Res 19:1331–1342
Regan D, Cynader M (1982) Neurons in cat visual cortex tuned to the direction of motion in depth: effect of stimulus speed. Invest Ophthalmol Vis Sci 22:535–550
Rushton SK, Wann JP (1999) Weighted combination of size and disparity: a computational model for timing a ball catch. Nat Neurosci 2:186–190
Savelsbergh GJP, Whiting HTA (1992) The acquisition of catching under monocular and binocular conditions. J Mot Behav 24:320–328
Savelsbergh GJP, Whiting HTA, Bootsma RJ (1991) Grasping tau. J Exp Psychol Hum Percept Perform 17:315–322
Savelsbergh GJP, Whiting HTA, Burden AM, Bartlett RM (1992) The role of predictive visual temporal information in the coordination of muscle-activity in catching. Exp Brain Res 89:223–228
Steeves JKE, Gray R, Steinbach MJ, Regan D (2000) Accuracy of estimating time to collision using only monocular information in unilaterally enucleated observers and monocularly viewing normal controls. Vision Res 40:3783–3789
van der Kamp J, Savelsbergh G, Smeets J (1997) Multiple information sources in interceptive timing. Hum Mov Sci 16:787–821
van der Kamp J, Bennett SJ, Savelsbergh GJP, Davids K (1999) Timing a one-handed catch II. Adaptation to telestereoscopic viewing. Exp Brain Res 129:369–377
Watson MK, Jakobson LS (1997) Time to contact and the control of manual prehension. Exp Brain Res 117:273–280
Wing AM, Turton A, Fraser C (1986) Grasp size and accuracy of approach in reaching. J Mot Behav 18:245–260
Acknowledgement
The authors are grateful to R. van Ee for helpful comments on the manuscript.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Mazyn, L.I.N., Lenoir, M., Montagne, G. et al. The contribution of stereo vision to one-handed catching. Exp Brain Res 157, 383–390 (2004). https://doi.org/10.1007/s00221-004-1926-x
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00221-004-1926-x