
Abstract
We investigated the neural correlates of dual task performance using EEG coherence as a measure of the functional coupling between cortical regions. Nine healthy participants performed a rhythmical movement with the right hand and an isometric contraction with the left hand, either initiated simultaneously or successively. EEG data revealed that dual task performance was associated with stronger coherence in left hemispheric and mesial areas than the sum of the tasks performed separately in the beta (>12–30 Hz), but not alpha (8–12 Hz), band. This effect was more pronounced when the two assignments were initiated simultaneously, as opposed to successively. The data demonstrate that the pattern of cortico-cortical coupling during bimanual actions is not just the sum of that associated with its component parts, but is increased according to coordinative demands and processing load.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Dual task performances during which assignments are carried out concurrently are part of our daily life activities. Despite the ease with which these acts can be executed, the overall processing demands may be high. This is particularly true when qualitatively distinctive functions are coordinated between the upper limbs. Accordingly, the study of bimanual dual task performance provides insights into how complex behavior is realized and how the coupling between cortical areas is modified as the activities of the hands contribute to the coordinative goal. Such coupling or functional connectivity can be followed through the coherence between scalp EEG signals from different regions (Gerloff et al. 1998; Serrien et al. 2003a). In particular, motor behavior is associated with EEG changes in the alpha (8–12 Hz) and beta (>12–30 Hz) bands (Andres et al. 1999; Brown 2000; Gerloff et al. 1998; Manganotti et al 1998; Serrien and Brown 2002; Serrien et al. 2003a).
However, cortico-cortical coupling is not merely dependent on the nature of the task per se but is also modulated by contextual factors. The latter may be related to environmental relationships, such as the anticipation of changes in the appropriate motor response (Serrien et al. 2003b, 2004a), or intrinsic factors more pertinent to task organization and execution, as when the same task is executed with or without the benefit of a representation held in working memory (Serrien et al. 2004b). To date this broad categorization has also been reflected in qualitative changes in the frequency of interregional coupling. Manipulations of the information provided by environmental cues and of the expectancy of imperative changes in the task effector both lead to changes in cortico-cortical coupling that predominantly affect the alpha band (Serrien et al. 2003b, 2004a). These observations are consonant with the supposition that EEG changes in the alpha band characterize cognitive-related processes (González-Hernández et al. 2002; Klimesch 1999). The association between alpha activity and cognitively driven operations is highlighted by a prominent effect of attention and behavioral preparation upon alpha activity (Deiber et al. 2001; Petsche et al. 1997; Ray and Cole 1985; Shaw 1996; Shibata et al. 1998).
The corollary of the above is that contextual factors that directly impinge on motor processing may be more overt in the beta band. The importance of cortico-cortical coupling in this band to motor planning and execution has been repeatedly demonstrated (Gerloff et al. 1998; Serrien and Brown 2002; Serrien et al. 2003a) and even finds reflection in the pattern of corticomuscular drive (Brown 2000). Thus in one study in which working memory was manipulated as an intrinsic contextual factor by varying the length of the delay interval after which motion had to be reproduced, the cortico-cortical coupling associated with movement production was changed in the beta, but not the alpha band, even though task performance remained the same (Serrien et al. 2004b). Here we further test whether internal contextual factors that directly impinge on motor organization are more likely to shape task-related cortico-cortical coupling in the beta than alpha band. In particular, we tested whether the extra coordinative demands of simultaneous performance of a task by each hand are associated with greater cortico-cortical coupling in the beta band than the sum of the coupling associated with the independently performed unimanual tasks. To emphasize the coordination requirements we used a continuous bimanual assignment that involved asymmetrical functions of the hands, characteristic of various daily-life activities. In addition, we predicted that a delay in the initiation of one of the two bimanually performed tasks would enlist a relatively lower degree of functional interregional connectivity during steady-state performance than immediate simultaneous bimanual performance. This is due to the potentially lower processing load in the former compared to the latter performance condition, as the delayed task can be interleaved with the control requirements of the already established task. We confirmed that changes in internal motor context preferentially modulated cortico-cortical coupling in the beta band, without changes in the behavioral performance of component tasks.
Materials and methods
Task and procedure
Right-handed subjects (n=9) gave informed consent to participate in the study, which was approved by the local ethics committee. Subjects were seated in front of a desk with custom-built manipulanda, the forearms secured in a neutral position. They were asked to carry out dual task performances that involved a rhythmical movement with the right hand and an isometric contraction with the left hand. For the rhythmical movement, subjects executed flexion/extension actions around the wrist joint in the horizontal plane paced by an auditory metronome with a cycle duration of 900 ms. For the isometric contraction, subjects were required to produce a power grip with a force level that corresponded to 20% of maximum force. The U-shaped manipulandum included strain gauges from which the signals were fed into an amplifier. The output was recorded and also connected to an oscilloscope which provided on-line visual feedback to the subjects. There were two different dual task conditions: simultaneous vs. successive initiation of the two acts (Fig. 1). One condition consisted of performing both tasks simultaneously from the start (dual task start), whereas in the second condition the isometric task was started about 20 s after the rhythmical movement had been initiated (dual task midway). The dual task start condition lasted 20 s whereas the dual task midway condition lasted 40 s but only the last 20 s involved the simultaneous performance of both tasks. Single task conditions were also included and involved either a rhythmical movement with the right hand paced by the metronome at 900 ms/cycle for 40 s, or an isometric contraction with the left hand at 20% of maximum grip force for 20 s. Subjects were told beforehand that dual or single task performances would be required and five trials of each condition were executed. Subjects practiced before the actual recording started so as to minimize modulations in the adopted movement amplitude. Rest (baseline) trials that included passive listening to the metronome tone were also recorded. The order of the conditions was randomized.

The dual task conditions. The right hand (rhythmical) and left hand (isometric) tasks were initiated simultaneously (dual task start) or successively (dual task midway). In the latter condition the isometric task was applied after the rhythmical movement had been initiated. The last 10 s of the actual 20 s of the unimanual rhythmical performance in the dual task midway condition are shown
Before the experiment started, ten silver-silver chloride electrodes were fixed with collodion over the subject’s scalp according to the International 10–20 System, and referenced to linked ears. The electrode positions of interest were F3/4, FC3/4, C3/4, P3/4 and Cz/FCz, which are likely to overlie the dorsal prefrontal, premotor, primary sensorimotor, superior parietal areas, and mesial motor areas, respectively (Homan et al. 1987; Steinmetz et al. 1989). The signals were amplified (Digitimer Ltd., Herts., UK), band pass filtered (0.5–100 Hz), digitized by a 1401 analogue to digital converter (Cambridge Electronic Design, Cambridge, UK) and sampled at a rate of 200 Hz (Spike 2, Cambridge Electronic Design, Cambridge, UK). Instances containing eye movement artifacts, main spikes or electromyographic artifacts were rejected.
Analysis
The behavioral measurements of the rhythmical right hand movement were amplitude and cycle duration. EEG coherence was used to assess functional coupling between the cortical areas in the frequency domain, and was estimated using the discrete Fourier transform and parameters derived from it. The coherence |R ab(λ)|2 was calculated by using the formula: |R ab(λ)|2=|f ab(λ)|2/f aa(λ)f bb(λ). In this equation, f characterizes the spectral estimate of two EEG signals a and b for a given frequency (λ). The numerator includes the cross-spectrum for a and b (f ab), whereas the denominator includes the autospectra for a (f aa) and b (f bb). Figure 2 depicts auto-spectra from the electrodes FCz and C3 and their cross-spectrum when performing the dual task start condition. A peak in the alpha (8–12 Hz) as well as beta (>12–30 Hz) band is evident. Coherences were transformed using the inverse hyperbolic tangent and evaluated in both frequency bands. Transformation was necessary to stabilize variances and was an essential step in the comparison of the dual task with the sum of its parts, as it increased the dynamic range of coupling strengths (which, otherwise, are capped at 1). In order to separate the task-related coherence (TRCoh) from the background coherence, and to eliminate as much as possible that due to volume conduction, the values of the resting state were subtracted from those of the active state. TRCoh values were divided into the following connectivity groupings: 6 left lateral hemisphere (F3-FC3, F3-C3, F3-P3, FC3-C3, FC3-P3, C3-P3), 6 right lateral hemisphere (F4-FC4, F4-C4, F4-P4, FC4-C4, FC4-P4, C4-P4), 16 interhemispheric (F3-F4, F3-FC4, F3-C4, F3-P4, FC3-F4, FC3-FC4, FC3-C4, FC3-P4, C3-F4, C3-FC4, C3-C4, C3-P4, P3-F4, P3-FC4, P3-C4, P3-P4), and 17 mesial (FCz-F3, FCz-F4, FCz-FC3, FCz-FC4, FCz-C3, FCz-C4, FCz-P3, FCz-P4, FCz-Cz, Cz-F3, Cz-F4, Cz-FC3, Cz-FC4, Cz-C3, Cz-C4, Cz-P3, Cz-P4). Thereafter, average TRCoh scores were calculated per connectivity grouping. In addition, EEG power was measured in the alpha and beta bands at the individual electrodes, and stabilized by logarithmic transformation. Task-related power (TRPow) was calculated by subtracting the values at rest from those of the active state. Averages were estimated across the electrodes for the different conditions.

Auto-spectra from the electrodes FCz and C3 and their cross-spectrum when performing the dual task start condition. The auto-spectra are scaled on the left axis and the cross-spectrum is scaled on the right axis (arbitrary units). Peaks are noted in the alpha (8–12 Hz) and beta (>12–30 Hz) bands at around 10 and 18 Hz, respectively
The analyses compared TRCoh differences from rest during dual tasks to the sum of the TRCoh differences from rest during the composite single tasks, for any given connection. We sought super-additive effects that could not be due to the simple linear summation of coupling strengths when two tasks are performed simultaneously. Super-additive effects were taken to reflect the extra costs of simultaneous task performance. This likely offers a conservative view of the extra costs as it assumes the worst-case scenario that TRCoh changes are additive with no instances of sharing of coupling elements between tasks. For the dual task conditions (dual task start and dual task midway) the first and last 2 s of the recordings were eliminated to limit motion onset and offset transients, resulting in epochs of 16 s/trial. For the single task conditions, equivalent time intervals of 16 s/trial were used. Accordingly, the right hand movement was evaluated at the start and midway to match the recordings of the dual tasks and to avoid changes due to fatigue, as the duration of movement prior to analysis was precisely matched between paired conditions. The number of evaluated epochs was the same for each condition, and measurements were concatenated to give 80 s per condition in each subject. The statistics of TRCoh included separate 2×4 (condition × region) ANOVAs for the start and midway paradigms. The first factor included the coherence scores of the dual task vs. sum of the single tasks, whereas the second factor referred to the four connectivity groupings: left hemisphere, right hemisphere, interhemispheric, and mesial. TRPow was evaluated by means of 2×3 (condition × region) ANOVAs. The first factor indicated the dual and corresponding single task conditions whereas the second factor represented the electrode groupings: left hemisphere, right hemisphere, and mesial.
The statistics of the behavioral data comprising the amplitude and cycle duration of the rhythmical right hand movement involved paired t-tests that compared dual vs. matching single tasks. Those of the isometric task included the percentage scores of maximum grip force and the root mean square of the force profiles by means of separate one-way (single, dual task start, dual task midway) ANOVAs. Data were transformed for normalization purposes. Data were corrected for multiple comparisons.
Results
Behavioral data
The amplitude and cycle duration of the right hand movement were not significantly different in the dual task and single task executions (P>0.05, for both). Amplitudes and cycle durations were 41±5° and 892±9 ms for the dual task start, 44±7° and 895±12 ms for the dual task midway, 43±3° and 897±7 ms for the single task start, 45±5° and 898±10 ms for the single task midway (mean ± SD). This indicates that the overall behavioral characteristics of the right hand movement were not significantly modulated in the different conditions. In correspondence with the instructions, the applied torque of the isometric contraction was close to the 20% of maximum grip force and was equivalent in the dual and single task performances (P>0.05). The average scores were 23±3%, 22±4%, 21±2% in the dual task start, dual task midway and single task, respectively. The root mean square of the force profiles did not differ significantly across the performance conditions (P>0.05). The average scores were 0.89±0.19, 0.87±0.14 and 0.85±0.10 in the dual task start, dual task midway and single task, respectively.
EEG data
TRCoh: The statistics in the alpha band showed no significant effects, P>0.05. The average TRCoh scores for the start paradigm were for the dual task vs. sum of the single tasks 0.05±0.03 and 0.03±0.02 (left hemisphere), 0.04±0.03 and 0.04±0.02 (right hemisphere), 0.05±0.05 and 0.04±0.03 (interhemispheric) and 0.04±0.04 and 0.02±0.01 (mesial). The average TRCoh scores for the midway paradigm were for the dual task vs. sum of the single tasks 0.06±0.03 and 0.04±0.02 (left hemisphere), 0.05±0.02 and 0.04±0.01 (right hemisphere), 0.06±0.04 and 0.05±0.05 (interhemispheric) and 0.05±0.03 and 0.03±0.04 (mesial).
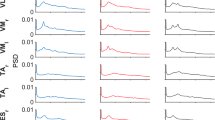
The statistics in the beta band showed a significant condition × region interaction, for the start paradigm (F (3,24)=11.5, P<0.01) and midway paradigm (F (3,24)=4.1, P<0.02). For both paradigms post hoc analyses revealed significant increases of TRCoh in the dual task vs. sum of the single tasks in the left hemisphere and mesial connections (P<0.05) whereas the functional couplings comprising the right hemisphere and interhemispheric connections were not significantly affected (P>0.05). Figure 3 illustrates TRCoh in the beta band for the different cortical regions in the start (A) and midway (B) paradigms, and shows that superior values during dual task execution were obtained for the left hemisphere and mesial connections. In particular, for the left hemisphere the highest TRCoh scores in the dual task conditions involved those of F3-C3 (0.13±0.06 for dual task start and 0.05±0.03 for dual task midway), and for the mesial connectivity grouping the highest scores involved Cz-C3 (0.11±0.04 for dual task start and 0.10±0.05 for dual task midway) and Cz-F3 (0.10±0.04 for dual task start and 0.08±0.03 for dual task midway). Figure 3C, D also illustrates the relative contribution of the isometric and rhythmic pattern to the sum of the single tasks.

TRCoh scores in the beta band associated with the dual task vs. sum of the single tasks conditions for the different cortical areas in the start (A) and midway (B) paradigms. The relative TRCoh scores of the isometric and rhythmic assignment that comprise the sum of the single tasks are also indicated for the start (C) and midway (D) paradigms. The cortical areas involved the left hemisphere, right hemisphere, interhemispheric and mesial connections. Averaged transformed data (mean ± SD). Significant differences between dual tasks vs. sum of the single tasks were noted for the left hemisphere and mesial connections (as indicated by asterisk)
The combined TRCoh scores of the left hemisphere and mesial connections were further analyzed to evaluate differences in functional interactions due to the context, in this case order, in which the dual task was executed. By means of a paired t-test, t (18)=3.5, P<0.01) a higher degree of TRCoh was noticed in the dual task start (0.08±0.05) compared to dual task midway (0.05±0.03). Figure 4 further illustrates this difference between the dual task conditions by means of TRCoh spectra for the F3-C3 connection in a representative subject. Note that the differences are apparent in the beta, but not alpha, band.

TRCoh spectra of the F3-C3 connection for both dual task conditions. An increased degree of coherence can be observed in the beta frequency band (>12–30 Hz) when both tasks are initiated simultaneously vs. successively. Individual transformed data
TRPow: The statistics revealed no significant effects (P>0.05). In the alpha band power decreased when performing the movement compared to rest. Expressed as percentage scores, the average power decreases for all electrodes were −17±8% and −14±7% for dual and single task start, −16±6% and −12±3% for dual and single task midway. In the beta band power also decreased when executing the task in comparison to rest. Expressed as percentage scores, the average power decreases for F3, FC3, C3 and P3 (for the left hemisphere grouping) were −12±7% and −9±5% for dual and single task start, −11±4% and −10±5% for dual and single task midway. The average power decreases for electrodes in the mesial grouping were −13±7% and −10±3% for dual and single task start, −12±6% and −8±3% for dual and single task midway. Therefore, although changes in power could contribute to increases in TRCoh with movement through non-linear effects (Florian et al. 1998), they could not explain the major differences in TRCoh between single and dual task conditions.
Discussion
Bimanual coordinated activity represents an exceptional ability of the human motor system and generally consists of subtasks carried out by each hand. Our experimental paradigm included dual task performances during which the upper limbs executed dissimilar acts, in contrast to previous studies (Serrien et al. 2003a). We focused on functional interregional connectivity as measured by EEG coherence, and hypothesized that the behavioral context related to the movement performance would modify the cortical dynamics. It was observed that neural interactions associated with dual task processing were increased over and above the summed contribution of the single tasks, even though no differences in behavioral characteristics were observed. Cortico-cortical coherence preferentially changed in the beta band. That no performance costs were associated with multitasking suggests that this increased interregional connectivity acted to limit the effect of bimanual interference.
The surplus of EEG-EEG coherence in the dual tasks suggests that extra functional communication was required to achieve coordination of the two components. It also implies that cortical interactions as expressed by EEG coherence reflect processing operations that encapsulate the regulation of complex functions such as multitasking, in the present case the coordination of the manual features into a bimanual activity. That more pronounced coherence increments occurred in the left than right hemisphere indicate an increased hemispheric asymmetry in movement control when both hands are concurrently active. Although we did not examine bimanual performance with the hand reverse, our findings are consistent with previous functional imaging data that have demonstrated that bimanual movements more strongly implicate the left than right hemisphere (Jäncke et al. 1998, 2000; Serrien et al. 2003a), and clinical work that has shown that left hemisphere damage impairs the ability to learn bimanual coordination (Wyke 1971) and produces bilateral motor deficits whereas right hemisphere damage is more prone to produce only contralateral deficits (Haaland and Harrington 1994). Overall, the present data are in line with a left hemispheric dominance for the representation of skilled movement (Haaland et al. 2000). Increased functional couplings for dual task performance were also observed between mesial and hemispheric connections, which denote the prominent involvement of midline structures, including the SMA, in complex acts such as bimanual movements (Andres et al. 1999; Sadato et al. 1997; Serrien et al. 2002; Stephan et al. 1999). In addition, it has been suggested that the SMA is involved in higher-order aspects of sensorimotor functions, including ideation of motor tasks (Shibasaki et al. 1993).
The results revealed that cortical dynamics were adapted in a context specific manner (Gerloff et al. 1998). In particular, the simultaneous compared to successive initiation of the component tasks in the dual task paradigm necessitated a relatively higher degree of functional interaction between cortical regions. Coherence increases as a function of task complexity (Manganotti et al. 1998; Serrien and Brown 2002), but this was an unlikely factor in the present experiment as the combination of the manual activities was similar in both dual task conditions, except in one important regard. With simultaneous initiation of the two component tasks there was the possibility of interference between the processing pathways used to select and/or produce the manual actions at the same time, whereas the successive retrieval of the tasks would reduce conflict as one action was already stably performed before the other was initiated, hence lowering the parallel processing load and permitting a more efficient unification of the responses into a coherent synergy. The intensification of the interregional connectivity could be viewed as a means of resisting the interference between processing pathways when the two component tasks were performed simultaneously. Along a similar line it has been shown that training of bimanual polyrhythmic sequences is more effective when the component tasks are learned according to an integrated timing control which merges the intervals into a common reference frame compared to a parallel motor organization (Summers and Kennedy 1992), stressing the importance of the strategy with which components are assimilated into the bimanual activity.
In conclusion, many behaviors require the integration of multiple actions into a synergy for successful achievement of the action goal. The present data illustrate that dual tasks necessitate increased functional connectivity compared to the sum of the single tasks, which suggests that extra information processing is required to manage the coordination of the components. This is particularly true when component tasks are initiated simultaneously.
The general significance of our findings is that they provide another example of how the cortical processing associated with a particular task is shaped by contextual factors rather than just the task itself. Contextual factors may be related to environmental relationships, such as the anticipation of changes in appropriate motor response (Serrien et al. 2003b, 2004a), or more pertinent to task organization and execution per se, as when the same task is executed with or without the benefit of a representation held in working memory (Serrien et al. 2004b) or a given task must be integrated with another executed with the other hand. The latter seem to preferentially involve modulations of functional cortico-cortical connectivity in the beta band.
References
Andres GG, Mima T, Schulman AE, Dichgans J, Hallett M, Gerloff C (1999) Functional coupling of human cortical sensorimotor areas during bimanual skill acquisition. Brain 122:855–870
Brown P (2000) Cortical drives to human muscle: the Piper and related rhythms. Prog Neurobiol 60:97–108
Deiber MP, Caldara R, Ibanez V, Hauert CA (2001) Alpha band power changes in unimanual and bimanual sequential movements, and during motor transitions. Clin Neurophysiol 8:1419–1435
Florian G, Andrew C, Pfurtscheller G (1998) Do changes in coherence always reflect changes in functional coupling. Electroencephalogr Clin Neurophysiol 106:87–91
Gerloff C, Hadley J, Richard J, Uenischi N, Honda M, Hallett M (1998) Functional coupling and regional activation of human cortical motor areas during simple, self-paced and metronome-paced finger movements. Brain 121:1513–1531
González-Hernández JA, Pita-Alcorta C, Cedeňo I, Bosch-Bayard J, Galán-Garcia L, Scherbaum WA, Figueredo-Rodriguez P (2002) Wisconsin Card Sorting Test synchronizes the prefrontal, temporal and posterior association cortex in different frequency ranges and extensions. Hum Brain Mapp 17:37–47
Haaland KY, Harrington DL (1994) Limb-sequencing deficits after left but not right hemisphere damage. Brain Cogn 24:104–122
Haaland KY, Harrington DL, Knight RT (2000) Neural representations of skilled movement. Brain 123:2306–2313
Homan RW, Herman J, Purdy P (1987) Cerebral location of international 10–20 system electrode placement. Electroencephalogr Clin Neurophysiol 66:376–382
Jäncke L, Peters M, Schlaug G, Posse S, Steinmetz H, Müller-Gärtner H-W (1998) Differential magnetic resonance signal change in human sensorimotor cortex to finger movements of different rate of the dominant and subdominant hand. Cogn Brain Res 6:279–284
Jäncke L, Peters M, Himmelbach M, Nosselt T, Shah J, Steinmetz H (2000) fMRI study of bimanual coordination. Neuropsychologia 38:164–174
Klimesch W (1999) EEG alpha and theta oscillations reflect cognitive and memory performance: a review and analysis. Brain Res Rev 29:169–195
Manganotti P, Gerloff C, Toro C, Katsuta H, Sadato N, Zhuang P, Leocani L, Hallett M (1998) Task-related coherence and task-related spectral power changes during sequential finger movements. Electroencephalogr Clin Neurophysiol 109:50–62
Ray WJ, Cole HW (1985) EEG alpha activity reflects attentional demands, and beta activity reflects emotional and cognitive processes. Science 228:750–752
Petsche H, Kaplan S, von Stein A, Filz O (1997) The possible meaning of the upper and lower alpha frequency for cognitive and creative tasks: a probability mapping study. Int J Psychophysiol 26:77–97
Sadato N, Yonekura Y, Waki A, Yamada H, Ishii Y (1997) Role of the supplementary motor area and the right premotor cortex in the coordination of bimanual finger movements. J Neurosci 17:9667–9674
Serrien DJ, Brown P (2002) The functional role of interhemispheric synchronization in the control of bimanual timing tasks. Exp Brain Res 147:268–272
Serrien DJ, Strens LHA, Oliviero A, Brown P (2002) Repetitive transcranial magnetic stimulation of the supplementary motor area (SMA) degrades bimanual movement control in humans. Neurosci Lett 328:89–92
Serrien DJ, Cassidy MJ, Brown P (2003a) The importance of the dominant hemisphere in the organization of bimanual movements. Hum Brain Mapp 18:296–305
Serrien DJ, Fisher RJ, Brown P (2003b) Transient increases of synchronized neural activity during movement preparation: influence of cognitive constraints. Exp Brain Res 153:27–34
Serrien DJ, Pogosyan AH, Cassidy MJ, Brown P (2004a) Anticipatory cortico-cortical interactions: switching the task configuration between effectors. Exp Brain Res 154:359–367
Serrien DJ, Pogosyan AH, Brown P (2004b) Influence of working memory on patterns of motor related cortico-cortical coupling. Exp Brain Res (in press)
Shaw JC (1996) Intention as a component of the alpha-rhythm response to mental activity. Int J Psychophysiol 24:7–23
Shibasaki H, Sadato N, Lyshkow H, Yonekura Y, Honda M, Nagamine T, Suwazono S, Magata Y, Ikeda A, Miyazaki M et al. (1993) Both primary motor cortex and supplementary motor area play an important role in complex finger movement. Brain 116:1387–1398
Shibata T, Shimoyama I, Ito T, Abla D, Iwasa H, Koseki K, Yamanouchi N, Sato T, Nakajima Y (1998) The synchronization between brain areas under motor inhibition process in humans estimated by event-related EEG coherence. Neurosci Res 31:265–271
Steinmetz H, Furst G, Meyer B-U (1989) Craniocerebral topography within the international 10-20 system. Electroencephalogr Clin Neurophysiol 72:499–506
Stephan KM, Binkofski FU, Halsband U, Dohle C, Wunderlich G, Schnitzler A, Tass P, Posse S, Herzog H, Sturm V, Zilles K, Seitz RJ, Freund H-J (1999) The role of ventral medial wall motor areas in bimanual co-ordination: a combined lesion and activation study. Brain 122:351–368
Summers JJ, Kennedy TM (1992) Strategies in the production of a 5:3 polyrhythm. Hum Mov Sci 11:101–112
Wyke M (1971) The effects of brain lesions on the learning performance of a bimanual co-ordination task. Cortex 7:59–72
Acknowledgements
The research was supported by the Medical Research Council of Great Britain and GlaxoSmithKline.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Serrien, D.J., Pogosyan, A.H. & Brown, P. Cortico-cortical coupling patterns during dual task performance. Exp Brain Res 157, 79–84 (2004). https://doi.org/10.1007/s00221-003-1822-9
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00221-003-1822-9