
Abstract
Rationale
The range of cognitive and psychotomimetic effects produced by antagonists of the N-methyl-D-aspartate (NMDA) receptor has lead to widespread usage of these molecules as pharmacological models of cognitive impairment for drug discovery. Historically, NMDA receptor antagonists have been used interchangeably on the assumption that they produce analogous effects.
Objectives
To profile a subset of these antagonists across a novel within-subject cognitive battery in the rat.
Methods
Naïve male Lister Hooded rats were subjected to a series of tests in which they were required to learn a simple visuo-auditory conditional discrimination. They then underwent testing in a delayed discrimination test followed by rule reversal and rule extinction tests.
Results
All NMDA receptor antagonists tested impaired acquisition performance and, with the exception of ketamine and the GluN2A preferring antagonist, NVP-AAM077, impaired consolidation of extinction. GluN2B antagonism produced a singular profile with potentially enhanced delayed discrimination performance and reduced hit rates in the reversal phase. Only PCP (phencyclidine) and ketamine disrupted performance in the delay phase but did so in a delay-independent manner. MK-801, PCP and memantine all increased the hit rate in the reversal phase; whilst only MK-801 and PCP impaired extinction per se.
Conclusions
NMDA receptor-dependent mechanisms are requisite in the acquisition of a simple conditional discrimination and consolidation of extinction. Their role in working memory and reversal tasks appear to be less critical and potentially specific to the paradigm and NMDA receptor antagonist used. It is clearly misleading to generalise across NMDA antagonists with respect to their preclinical cognitive profile.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
The N-methyl-D-aspartate (NMDA) receptor is an ionotropic glutamate receptor that has ubiquitous expression throughout the mammalian central nervous system (CNS; for review, see Dingledine et al. 1999). It has attracted a great deal of scientific attention as it has been implicated in a wide variety of neurological and pathological disorders such as schizophrenia, Alzheimer's disease and stroke. The observation that non-competitive NMDA receptor antagonists such as phencyclindine (PCP) and ketamine may induce or exacerbate schizophrenic-like symptoms in man (Krystal et al. 1994; Lahti et al. 1995) has lead to the hypothesis that NMDA-receptor dysfunction is a core deficit in the schizophrenic syndrome. In rodents, NMDA receptor antagonists induce a wide spectrum of behavioural responses including hyperlocomotion, stereotypy and loss of sensorimotor gating (Geyer et al. 2001; Mansbach and Geyer 1989; Tricklebank et al. 1989).
In addition to their psychotomimetic effects, NMDA receptor antagonists may induce performance deficits across a number of cognitive domains following either intra-cerebral or systemic administration. Attention has been particularly focused on hippocampal-dependent tasks due to the importance of this substrate in human episodic memory (Milner 2005). A wide variety of studies has shown that blockade of NMDA receptors in the hippocampus by direct infusion of the NMDA receptor antagonist, 2-amino-5-phosphopentanoate will prevent spatial learning in tasks such as non-matching to place in a T maze (Tonkiss and Rawlins 1991), Morris watermaze (Morris 1989; Morris et al. 1986), or the encoding of a paired associate task (Day et al. 2003). It is not just spatial memory tasks that have been shown to be dependent on NMDA receptors. For example, blockade of NMDA receptors in perirhinal cortex will produce deficits in the acquisition and/or consolidation of long-term novel object memories as assessed in a spontaneous object recognition test (Barker et al. 2006a, b; Winters and Bussey 2005; Abe et al. 2004). Effects on learning and memory are not surprising given that activation of these receptors underlies basal excitatory synaptic transmission, long-term potentiation and long-term depression (Bliss and Collingridge 1993; Morris 1989).
The behavioural consequences of NMDA receptor antagonism have led to the use of NMDA receptor antagonists not only as models of psychosis but also as models of cognitive dysfunction in disorders such as schizophrenia and Alzheimer's disease. Some of these behavioural deficits may be reversed by atypical antipsychotics; hence these models have become widely used as screening assays in drug discovery in the search for novel antipsychotic and procognitive compounds. However, the NMDA receptor is a complex heterogeneous structure comprised of GluN1, GluN2 and GluN3 (NR1, NR2, NR3) subunits with GluN2 itself being represented by four different genes, GluN2A, GluN2B, GluN2C and GluN2D (Alexander et al. 2008). A range of NMDA receptor antagonists have been described with varying degrees of subtype selectivity and mechanism of action: (i) phencyclidine, MK-801 and ketamine are the prototypical, non-competitive open channel blockers, devoid of subunit selectivity (Lodge and Johnson 1990); (ii) memantine acts similarly but with lower affinity (Johnson and Kotermanski 2006); (iii) the non-competitive antagonists, CP 101,606 and Ro 25-6981 have substantial selectivity for GluN2B-containing receptors (Chenard et al. 1995; Menniti et al. 1997; Fischer et al. 1997) and (iv) NVP-AAM007 which has some, albeit limited, selectivity for GluN2A-containing receptors (Auberson et al. 2002). Historically, NMDA receptor antagonists such as PCP, MK-801 and ketamine have been used interchangeably preclinically on the assumption that they produce analogous effects on a variety of cognitive and non-cognitive behaviours. Quite clearly, this is over-simplistic when consideration is given to the different biochemical profile of the drugs and their pharmacological cross-reactivity. Furthermore, despite the similarity of memantine to ketamine in terms of its mechanism of action and biophysical properties (Johnson and Kotermanski 2006), the use of memantine as a treatment for Alzheimer's disease is clearly at odds with the clinical experience of ketamine: the latter being used to model psychosis and cognitive disruption. Indeed, only ketamine of the NMDA receptor antagonists has been extensively tested in the clinical setting. In contrast, MK-801 and PCP are far more commonly used as research tools within the preclinical literature.
It is therefore somewhat surprising that there has been no systematic comparison of the range of compounds available on measures of cognitive and non-cognitive behaviours. This gap has been recently addressed with the profiling of a number of NMDA receptor antagonists, including GluN2A and GluN2B subtype-selective antagonists, across a broad battery of behavioural tests. In the first body of work, the effect of a range of NMDA receptor antagonists on the ability of animals to respond under various variable interval (VI) schedules of reinforcement was examined. Not only were large differences in effect size observed in this study, but the different antagonists tested produced qualitatively distinct effects on instrumental responding (Gilmour et al. 2009). In this second study, NMDA receptor antagonists were profiled through four consecutive cognitive tests designed to access the neuronal systems underlying learning, memory and cognitive flexibility. Rats were initially trained to press levers before entering phase 1 (rule acquisition) in which animals had to learn a simple conditional discrimination to a visual and auditory cue over five daily training sessions with drugs given each day immediately beforehand. Both dopaminergic and glutamatergic processes have been shown to be important in the acquisition of conditional discriminations (Dunn and Killcross 2006; Murray et al. 1995). It is therefore not surprising that schizophrenic patients have been reported to show equivalent deficits (Hofer et al. 2001). In phase 2 (delayed discrimination), a delay of 1, 4 or 8 s was inserted between presentation of the stimuli and access to the choice levers. Working memory paradigms are intrinsically different in rodents and humans, as is the definition of working memory itself. However, delayed (non) match-to-sample procedures in rodents are dependent on medial prefrontal cortex (Sloan et al. 2006) and it is accepted that the prefrontal cortex is critical for working memory in humans (Petrides 2000a, b; Goldman-Rakic 1987). Furthermore, solution of the delayed discrimination task requires the holding of information during the delay period and thus is consistent with human definitions of working memory. Deficits in working memory are one of the core cognitive symptoms of schizophrenia (Barch et al. 2009). The aim of phase 2 of the battery was therefore to assess whether NMDA receptor antagonists produce a delay-dependent deficit in this novel rodent working memory task. NMDA receptor antagonists typically produce delay-independent deficits in delayed (non) matching paradigms (Han et al. 2000; Stephens and Cole 1996). In phase 3, the contingency between the conditional stimulus presented and the rewarded lever was reversed. Schizophrenic patients show deficient cognitive flexibility evident as poor performance in both extra-dimensional set shifting and reversal learning paradigms (e.g. McKirdy et al. 2009). Both acute and chronic treatment with PCP can produce reversal learning deficits in a lever-press reversal paradigm (Abdul-Monim et al. 2003; Abdul-Monim et al. 2006). The aim of this phase was, therefore, to examine the effect of NMDA receptor blockade in this novel reversal task. The final phase of the battery, phase 4, was designed to assess extinction learning and its consolidation. The animals continued to be presented with two levers but only one of these was now rewarded. Drugs were again administered immediately before testing, as in phases 1–3. On the following day, the animals were retested in a drug-free state hence both the effect of NMDA receptor blockade on extinction itself and then on the consolidation of this learning could be assessed. Whilst there are a number of reports showing that consolidation of fear extinction is NMDA receptor-dependent, appetitive extinction is rarely tested.
To summarise, the aim of the present study was to further understand (a) the role of the NMDA receptor across a range of cognitive domains and (b) to test the functional equivalence of antagonists both within and between test phases and cognitive domains. The work further builds on the body of data accumulated by the Lilly laboratories in an attempt to elucidate the behavioural phenotype produced by a broad spectrum of NMDA receptor antagonists. This database will enhance our understanding of the translational potential of both our preclinical tests and pharmacological models of CNS disorders such as schizophrenia. The results demonstrate that the processes of acquisition of an operant visuo-auditory conditional discrimination task and the consolidation of its extinction are generally prevented by NMDA receptor antagonists regardless of mechanism of action or subtype selectivity, whilst effects on measures associated with working memory and behavioural flexibility are very much more compound specific and bidirectional.
Methods
Subjects
Forty-eight naïve male Lister Hooded (250–350 g) rats were used as subjects for each experiment. The only exception was Memantine II in which 64 rats were used to allow for an additional ketamine control group. Animals were housed in groups of four, receiving only water ad libitum. Body weights were maintained at no less than 85% of their free-feeding weights (including allowance for growth) by food pellets presented during experimental sessions and by post-session feeding. All animals were supplied by Harlan, UK and housed in individually ventilated caging where they were provided with sawdust and environmental enrichment (tunnels and balls). They were maintained on a 12-h/12-h light/dark cycle (lights on at 0700 h) and at a room temperature of 21 ± 2°C. All procedures complied with regulations laid down in the United Kingdom Animals (Scientific Procedures) Act, 1986, and were approved by the Ethics Committee of the Lilly Research Centre.
Materials
Each experiment was performed in one of two sets of 16 standard operant chambers (Med Associates Inc. US). Each chamber was housed in a sound and light attenuating cabinet and comprised a house light, two retractable levers; each with a stimulus light above it. The levers were located either side of a recessed magazine where food pellets (Noyes, 45 mg, Formula P) were delivered from an automatic pellet dispenser. The start of a session was signalled by onset of the house light, and its permanent offset indicated the end of a session. Each chamber contained standard grid floors that were wiped with disinfectant wipes (Excel 30, Shiloh Healthcare) between each subject. Auditory signals using a continuous tone and a clicker were also available. Experimental sessions were controlled and data recorded using programmes written in-house using MedPC-IV software. Data were prepared for analysis using an in-house Microsoft Excel macro designed for the experiment.
Protocol
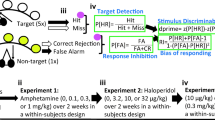
An overview of the procedure can be found in Fig. 1. Animals were initially trained to lever press for an appetitive reward. They then received seven sessions in which they were trained to acquire a simple visuo-auditory conditional discrimination. Animals received drug treatments prior to each discrimination acquisition session but not prior to lever-press acquisition sessions. The rats then underwent further ‘baseline’ drug-free training sessions to ensure all animals had reached criterion; this was followed by the delayed conditional discrimination session. Once again animals had a drug-free baseline session before entering the two sessions of the reversal phase. Animals were dosed prior to the delayed discrimination test and prior to each session of the reversal tests. Finally, animals received a drug-free lever-press session followed by the two extinction sessions. Animals were dosed prior to the first extinction session but not the second. It is important to note that in order to reduce the impact of carry-over effects animals were pseudorandomly reassigned to a different drug treatment prior to each test phase. Groups were balanced to ensure they were matched for performance in the baseline session preceding each phase and that there was a reasonable distribution reflecting each animal's drug history.

Summary of cognitive battery testing protocol. The schematic represents the standard within-subject training and testing protocol that all animals were subjected to in this series of studies. During initial lever training, rats learned to associate the pressing of either lever in an operant box with the delivery of food reward. Phase 1—acquisition: the first stage of the test battery involved learning to acquire a conditional visuo-auditory discrimination, where one cue (either clicker or light) signalled reinforcement availability on pressing one lever while the other cue signalled reinforcement from the alternative. Animals were dosed during the entire acquisition phase. Phase 2—delay: during the delay phase of the test battery, a variable time interval was imposed between cue presentation and lever availability during performance of the conditional discrimination to tax working memory. Drug was administered only once during this phase. Phase 3—reversal: during the reversal phase the animals performed the no-delay conditional discrimination, but in this phase the original cue-lever contingencies were reversed. Drug was administered during two reversal test sessions. Phase 4—Extinction: during the final extinction phase, both levers were presented to animals, but only one was rewarded. No cues were presented during this phase. There were two extinction sessions, and drug was administered prior to the first of these. When moving to the next phase of the cognitive battery, animals were trained to criterion on the previous phase as necessary, and data from these training sessions was used to rank performance and re-allocate drug treatments to animals to maintain matched groups throughout the entirety of the study
Lever-press training
In the first lever-press training session rats received ten pellets on a VI-18 schedule of delivery (variable interval; approximately every 18 s; range 10–25 s). The delivery of a food reward was signalled by a 0.5 s tone. This was followed by a 1-minute timeout period (lights out). They then entered the lever-press section in which there was pseudorandom presentation of the levers. Each trial comprised insertion of one or other of the levers. The rat was rewarded with a food pellet (signalled with a 0.5 s tone) when it pressed the lever and the levers were then retracted. Levers were presented on a VI-18 schedule and rewarded on a FR-1 schedule (fixed ratio; i.e. each lever press was rewarded). The session ended after 30 min. The second session was identical to the first except that there was no free delivery of pellets. If more than eight animals did not reach the inclusion criterion of greater than ten presses in 30 min a third day of lever-press training was included. This session was identical to the previous session. Data from the final training session were used to create matched treatment groups.
Phase 1: visuo-auditory conditional discrimination acquisition
Animals were trained to acquire a simple visuo-auditory conditional discrimination between a clicker and signal lights. There were 90 trials in each session (45 light and 45 clicker). Each trial comprised a 4-s signal (illumination of either signal lights or a 10 Hz clicker) followed by insertion of both levers. If an auditory (clicker) signal was given, pressing on the left lever was rewarded. If a visual (lights) signal was given, pressing on the right lever was rewarded. Lever-signal contingency was counterbalanced between animals. If an animal made a correct response it was counted as a hit and reinforced (pellet delivery signalled by tone). An incorrect response had no programmed consequences. The levers retracted when the rat had made a lever press. If a rat failed to respond within 10 s it was counted as an omission and the levers were retracted. There were no correction trials. Animals underwent seven sessions on this programme; there was a two day break between sessions 3 and 4. This was the only phase completed by the Memantine II cohort; a second cohort of animals was tested at a higher dose range as no significant effects were revealed in the first study (memantine I). Ketamine (10 mg/kg) was included as a positive control in this second cohort.
Phase 2: visuo-auditory delayed conditional discrimination
This was identical to the above programme except that three delays were imposed between the offset of the signal and the insertion of the levers. There were 30 trials each of 1-, 4- and 8-s delays. Animals had one day of training on the standard discrimination programme followed by a number of remedial sessions for those animals that had not acquired the discrimination (hit rates of 0.70 or less). This was followed by a baseline session prior to the delayed discrimination test. Data from this session were used to create matched treatment groups for the delay test; i.e. animals were reassigned to a treatment group that was independent to their assignment in the preceding phase. Animals that had a hit rate of less than 0.65 during this baseline session were excluded from the analyses.
Phase 3: reversal of visuo-auditory conditional discrimination
This was identical to the discrimination programme above except that the lever-signal contingency was reversed for all animals. Animals had one day of training on the standard discrimination programme prior to the reversal test. Data from this session were used to create matched treatment groups for the delay test; i.e. animals were reassigned to a treatment group that was independent to their assignment in the preceding phases. Animals that had a hit rate of less than 0.65 were excluded from the analyses. In the MK-801, PCP, CP 101,606 and ketamine studies, the reversal tests was performed over two sessions. For the memantine study, only one session of reversal was performed. The reversal phase was excluded from the Ro 25-6981 experiment.
Phase 4: extinction test
In this test, both levers were presented simultaneously, however only one of these levers was rewarded. There were 90 trials in three blocks of 30. Animals had one day of training prior to the extinction test which was identical to the second session of lever-press training (above). Data from this session were used to create matched treatment groups for the delay test; i.e. animals were reassigned to a treatment group that was independent to their assignment in the preceding phases. No animals were excluded. The left/right assignment of the rewarded lever was counter balanced across animals. The following day, the test was repeated but no drug was administered.
Measurements and statistical analysis
Two measures are presented: the omission rate (OR) and the hit rate (HR; proportion of correct trials of responses made). During the extinction phase the measure was the proportion of responses on the non-rewarded (extinguished) lever. High doses of the NMDA receptor antagonists induced high levels of omissions in some animals. Subjects were excluded from the HR analyses if they omitted more than 75% of their trials. If this resulted in fewer than four animals remaining in a treatment group then the whole treatment group was excluded from the HR analysis. Data from all animals were included in the omission rate analyses. ORs were not normally distributed as groups tended to the extremes of the range (0–1). Analyses were therefore performed on the square root of the OR. If all groups made fewer than five omissions (OR < 0.056; arbitrarily determined) no statistical analysis was performed as it was unlikely to be of biological relevance.
All statistics were calculated using Statistica v.8 (Statsoft, UK). A general linear model, a priori approach was used for all Analyses of Variance (ANOVA) conducted for measured parameters. The HR and OR in each phase was submitted to an appropriate mixed ANOVA with the between-subject factor of drug (i.e. dose of the test compound); the within-subject factors varied dependent on the phase. For the acquisition phase OR data from all seven sessions were analysed with the within-subject measure of session. In no experiment were significant effects of the drug revealed in the first three sessions, indeed no measurable learning was evident in these sessions hence they simply add variance to the data. The HR ANOVA was therefore performed over sessions 4–7 only. In the delay phase, the within-subject factor of delay (1, 4 or 8 s) was included in the analysis. The reversal phase comprised six blocks of 30 trials across two sessions except for the memantine study in which only one session was performed. The OR ANOVA included the within-subject factor of session (1–2) whilst the HR ANOVA included the factor of block (1–6). The OR data for the extinction phase included the factor of session (1–2). Each extinction session was split into three blocks of 30 trials. The sessions were analysed separately as they aimed to test distinct hypotheses. In each analysis block (1–3) was the within-subject factor. Significant main effects and interactions were followed with planned comparisons against the appropriate vehicle control group. In each analysis, statistical significance was defined as p < 0.05.
Forty-one additional analyses were performed in order to test for carry-over effects of each animals' drug history. The HR data were submitted to separate ANOVAs comprising the additional between-subject of ‘previous treatment’—separate analyses were performed for each preceding phase (i.e. the extinction HR data were submitted to three further ANOVAs with the factors: (1) AQ treatment group, (2) delay treatment group and (3) reversal treatment group).
Drugs
PCP (Phencyclidine hydrochloride, Sigma–Aldrich, UK), (S)-(+)-ketamine hydrochloride (Sigma–Aldrich, UK), MK-801 hydrogen maleate (dizocilpine, Sigma–Aldrich, UK), Memantine hydrochloride (Tocris, UK), Ro 25-6981 ([(R:(+), S:(-))-α-(4-hydroxyphenyl)-β-methyl-4-(phenylmethyl)-1-piperidinepropanol, Lilly Research Labs]), CP 101,606 ((1S,2S)-1-(4-Hydroxyphenyl)-2-(4-hydroxy-4-phenylpiperidino)-1-propanol, Lilly Research Labs) and NVP AAM-077 ([(R)-[(S)-1-(4-bromo-phenyl)-ethylamino]-(2,3-dioxo-1,2,3,4-tetrahydroquinoxalin-5-yl)-methyl]-phosphonic acid, Lilly Research Labs) were used in this study. Memantine, MK-801, PCP and ketamine were all dissolved in 5% (w/v) glucose vehicle. CP 101,606 was initially dissolved in a small volume of 10% lactic acid before being made to volume with 5% (w/v) glucose. Ro 25-6981 was dissolved in 0.9% (w/v) saline. NVP AAM-077 was dissolved in 5% (v/v) Pharmasolve (N-methylpyrrolidone) then brought to volume with 5% (w/v) glucose solution. Where necessary, 1 M sodium hydroxide was used to adjust the pH towards neutral. All drugs were formulated daily and administered in a volume of 1 ml/kg via the subcutaneous route with the exception of NVPAAM-077 which was administered by the intraperitoneal route. All doses refer to free base or acid weights of compounds. Drug administration was 30 min prior to test sessions for all compounds with the exception of CP 101,606 which was administered 60 min prior to test sessions (i.e. acquisition sessions 1–7, the delay test, the two reversal sessions and day 1 of the extinction phase).
Results
Full statistical analyses can be found in Tables 1, 2, 3, 4 and 5 of the online supplementary material. ORs of less than 0.056 are not reported nor were they submitted to statistical analysis. The highest dose of MK-801 (0.25 mg/kg) produced severe ataxia and motor deficits such that most animals made fewer than ten responses. This group was excluded from the hit rate analyses for each phase. Similarly, the highest dose of PCP (5 mg/kg) induced a high level of omissions in the acquisition phase and its data were removed from the analyses. In this study, the decision was made to remove this dose from the subsequent phases and include a lower dose of 0.5 mg/kg. Animals treated with Ro 25-6981 developed local skin lesions at the site of injection. They had a 3-week recovery period between the end of the acquisition phase and the start of the delay phase. They did not undergo testing in the reversal phase. In the Memantine study, only one session of the reversal phase was performed.
Data from baseline sessions preceding each phase of the battery were analysed. In all studies, baseline performance (drug free) was not statistically significant between the treatment groups and has not been presented. Baseline lever-press data in the final lever-press session preceding the acquisition phase (number of lever presses in the 30-min session) can be found in Table 1 of the online supplementary material. It was necessary to include a third day of lever-press training in two of the studies (CP 101,606 and ketamine) as a high number of animals did not reach the inclusion criterion of greater than ten presses within the 30-min session.
Phase 1: visuo-auditory conditional discrimination acquisition
Vehicle-treated animals from all studies learned to discriminate between the two stimuli resulting in HR performance levels with group means between 72% and 81%. All seven compounds produced a significant disruption of performance such that the animals at the highest dose tested failed to acquire the discrimination (see Fig. 2, 3). All compounds except NVP-AAM077 produced a concomitant increase in the OR (see Fig. 2). It should be noted that in each experiment the lowest dose that produced a blockade of acquisition did so in the absence of a marked effect on the OR. This distinction was particularly striking in the case of the GluNR2B antagonists (Ro 25-6981 and CP 101,606) which produced mild effects evident in session 1 only. Animals dosed with high doses of MK-801 (0.25 mg/kg), PCP (5 mg/kg) and ketamine (25 mg/kg) were severely motorically impaired and unable to respond. With the exception of MK-801 (0.25 mg/kg) and PCP (5 mg/kg) animals habituated to the adverse effects of the treatments across the seven days resulting in a decrease in the OR across the sessions (Fig. 2).

Phase 1: rule acquisition. The effect of NMDA receptor antagonists on the omission rate during the acquisition of a simple visuo-auditory conditional discrimination. The figures show the mean omission rate (OR; ±SEM) calculated for each treatment group across seven sessions of testing. The asterisks represent significant effects following planned comparisons against the vehicle group. Asterisks along the x-axis refer to significant main effects; asterisks marked on the graph refer to significant effects from the interaction term: * p < 0.05, **p < 0.01, ***p < 0.001

Phase 1: rule acquisition. The effect of NMDA receptor antagonists on the acquisition of a simple visuo-auditory conditional discrimination. The figures show the mean hit rate (HR; ±SEM) calculated for each treatment group across seven sessions of testing. The asterisks represent significant effects following planned comparisons against the vehicle group. Asterisks along the x-axis refer to significant main effects; asterisks marked on the graph refer to significant effects from the interaction term: * p < 0.05, **p < 0.01, ***p < 0.001
Phase 2: visuo-auditory delayed conditional discrimination
Introducing a delay between the conditioned stimulus and the choice phase induced a delay-dependent disruption in performance regardless of drug treatment (see Fig. 4). MK-801, Ro 25-6981, NVP-AAM077 and memantine did not disrupt performance in this paradigm. The ANOVA on the HR for MK-801 revealed a significant interaction between delay and dose; however planned comparisons found no significant effects against the vehicle control group at any of the delays. PCP (2.5 mg/kg) produced a delay-independent disruption in performance; this effect was non-selective as there was a small, but significant, increase in the OR (ORs at 1, 4 and 8 s, respectively: 0.15 ± 0.08, 0.16 ± 0.09, 0.16 ± 0.09). Ketamine (25 mg/kg) also produced a deficit in performance which was evident at the short (1 s) delay only. It also induced an increase in the OR (ORs at 1, 4 and 8 s, respectively: 0.31 ± 0.1, 0.24 ± 0.09, 0.30 ± 0.09). Of particular note was the significant delay-independent enhancement in performance produced by CP 101,606. There was no evidence of dose dependency in this effect. MK-801 induced a significant increase in the OR at the highest dose such that this group was excluded from analysis (ORs at 1, 4 and 8 s, respectively: 0.87 ± 0.1, 0.88 ± 0.1, 0.86 ± 0.1).

Phase 2: Delayed Discrimination. The effect of NMDA receptor antagonists on performance of delayed conditional visuo-auditory discrimination in which a delay was inserted between presentation of the stimuli and presentation of the levers. The figures show the mean hit rate (HR ± SEM) calculated for each treatment group at three delay periods: 1, 4 and 8 s. The asterisks represent significant effects following planned comparisons against the vehicle group. Asterisks along the x-axis refer to significant main effects; asterisks marked on the graph refer to significant effects from the interaction term: * p < 0.05, **p < 0.01, ***p < 0.001
Phase 3: reversal of visuo-auditory conditional discrimination
Reversing the contingency between the signal and the lever clearly severely disrupted performance in all treatment groups across experiments (see Fig. 5). Vehicle-treated animals showed some limited evidence of acquiring the new contingency in the MK-801, CP 101,606 and NVP-AAM077 studies (planned comparisons of B1 vs B6: p < 0.05), but not in the PCP or ketamine study (B1 v B6: both p > 0.1). In the memantine experiment, planned comparisons between block 1 and block 3 in the vehicle-treated animals revealed no significant learning within this group (p < 0.05). The effect of the NMDA antagonists on the HR produced a heterogeneous pattern of data with four profiles: (1) MK-801 (0.1 mg/kg), PCP (2.5 mg/kg) and memantine all increased the hit rate relative to control animals however there was no evidence of learning within any of these treatment groups (B1 v B6: all p > 0.1); (2) NVP-AAM077 (10 mg/kg) produced a significant elevation in the HR in block 3 (day 1) relative to the vehicle control animals; in contrast (3) CP 101,606 significantly reduced the HR compared to control animals; whilst (4) ketamine failed to produce any significant effects on the HR regardless of a significant elevation in the OR at 10 and 25 mg/kg (session 1 and 2: 10 mg/kg −0.19 ± 0.09, 0.1 ± 0.07; 25 mg/kg −0.58 ± 0.11, 0.62 ± 0.09). Concomitant significant increases in the OR were observed for MK-801 (session 1 and 2: 0.1 mg/kg −0.11 ± 0.05, 0.02 ± 0.01; 0.25 mg/kg −0.95 ± 0.2, 0.87 ± 0.02) and PCP (session 1 and 2: 2.5 mg/kg −0.7 ± 0.1, 0.54 ± 0.11) but not for memantine, CP 101,606 or NVP-AAM077.

Phase 3: reversal. The effect of NMDA receptor antagonists on reversal performance in which the contingency between signal and reward was reversed. The figures show the mean hit rate (HR ± SEM) calculated for each treatment group across two consecutive sessions of testing (memantine was tested in one session only). Each session is presented into three blocks of 30 trials. The asterisks represent significant effects following planned comparisons against the vehicle group. Asterisks along the x-axis refer to significant main effects; asterisks marked on the graph refer to significant effects from the interaction term: * p < 0.05, **p < 0.01, ***p < 0.001
Phase 4: extinction
All control animals acquired a preference for the rewarded lever and extinguished responding on the non-rewarded lever within the first session (planned comparisons between B1 and B3: all p < 0.05). Only MK-801 (0.1 mg/kg) and PCP (2.5 mg/k) produced significant disruption such that animals continued to respond on the non-rewarded lever (see Fig. 6). PCP (2.5 mg/kg) also produced a non-specific effect evident as an increase in omissions in session 1 only (0.65 ± 0.1). The second and drug-free session of the extinction phase aimed to assess whether animals remembered the preceding extinction phase. All compounds tested, with the exception of ketamine and NVP-AAM077, produced a selective disruption in performance. This was evident as an increase in responding on the non-rewarded lever during session 2 at a dose that had produced no significant effects on the HR or OR during session 1. Ketamine (25 mg/kg) also induced an increase in omissions (0.22 ± 0.09).

Phase 4: extinction. The effect of NMDA receptor antagonists on extinction. The figures show the mean proportion of responses on the non-rewarded lever (±SEM) calculated for each treatment group across two consecutive sessions of testing. Drugs were administered prior to session 1 only. Each session is presented into three blocks of 30 trials. The asterisks represent significant effects following planned comparisons against the vehicle group. Asterisks along the x-axis refer to significant main effects; asterisks marked on the graph refer to significant effects from the interaction term: * p < 0.05, **p < 0.01, ***p < 0.001
Carry-over effects
Forty-one additional ANOVAs were performed to assess whether prior drug treatment affected subsequent performance. One significant effect was revealed: the delay phase performance of animals treated with CP 101,606 was significantly affected by the drug treatment that they received during the acquisition phase (F3, 37 = 3.2, p < 0.05) such that animals treated with either 3 or 10 mg/kg CP 101,606 in the AQ phase were impaired relative to the vehicle control (both p < 0.05). Comparisons at each delay showed effects of 10 mg at both 1 and 4 s, and the effect of 3 mg at 4 s only. There was no interaction with the delay phase drug treatment such that CP 101,606 remained significant (F3,25 = 5.4, p < 0.01).
General discussion
In this study seven NMDA receptor antagonists were profiled across four sequential cognitive operant tests to examine the equivalence of these preclinical tools. The data demonstrated that whilst there was some clear commonality of effect, qualitatively different performance patterns were revealed across the four different test phases. A summary of the effect size within each phase is provided in Table 1. There are two primary implications of these findings: firstly, as intended by design, different phases of the battery show differential selectivity to pharmacological treatment and hence may have different and potentially dissociable neural substrates; and secondly, NMDA receptor antagonists produce performance impairments across different cognitive domains that are maybe both compound and/or domain specific.
NMDA receptor antagonist profile across the battery
Phase 1: rule acquisition
All seven compounds disrupted the ability of animals to acquire the conditional discrimination. MK-801, PCP, the two GluN2B-selective antagonists (Ro 25-256981 and CP 101,606) and the GluN2A-preferring NVP-AAM077 showed selective cognitive disruption at doses that did not produce effects on omissions. In contrast, the minimal dose of ketamine and memantine required to produce a deficit in acquisition also concomitantly produced an increase in omissions. Despite tolerance to the non-specific effects of the compounds (as evident in the decrease in omissions with repeated dosing) there was very little evidence of learning in the highest dose group of each compound with the exception of NVP-AAM077 (10 mg/kg). The effect of the latter compound was modest in comparison to the other antagonists. Whilst a higher dose may have produced a more severe learning deficit it is likely to have also induced a very large increase in omissions. In our previous work (Gilmour et al. 2009), 10 mg/kg NVP-AAM077 was essentially equivalent in effect to 0.25 mg/kg MK-801, 2.5 mg/kg PCP and 10 mg/kg ketamine on lever pressing on a VI schedule demonstrating a clear dissociation in this task. Our data are consistent with previous reports that MK-801 impairs acquisition but not performance of a simple visual conditional discrimination in a Y maze (Murray et al. 1995). The NMDA receptor is considered to be critical for hippocampus-dependent learning and memory, Pavlovian-conditioned approach behaviour and also in the acquisition but not expression of appetitive instrumental learning (i.e. lever-press acquisition; Baldwin et al. 2000; Bannerman et al. 2006; Di Ciano et al. 2001; Kelley et al. 1997; Morris et al. 1986; Pallares et al. 1995; van der Meulen et al. 2003). These data demonstrate that NMDA receptor-dependent mechanisms are also important in the acquisition of appetitive conditional discrimination learning, and that these effects are more likely GluN2B than GluN2A mediated. Furthermore, conditional learning deficits reported in schizophrenia (Hofer et al. 2001) may indeed reflect glutamatergic deficits.
Phase 2: delayed discrimination
Animals showed a clear delay-dependent disruption in performance during delayed conditional discrimination consistent with working memory being engaged during this paradigm. Dissociation in task sensitivity was found such that doses that impaired acquisition did not impair delayed discrimination performance, i.e. the NMDA receptor is critical for acquisition but not performance of this paradigm. Furthermore, no evidence was found supporting a critical role for the NMDA receptor in working memory in this paradigm as no delay-dependent deficit was revealed. Working memory impairment manifests as a deficit at longer delays whilst performance at the shortest delays remains intact. Indeed, only PCP and ketamine produced any disruption in this paradigm, apparent only at doses that induced concurrent increases in omissions. Working memory deficits are a core deficit evident in schizophrenia (Barch et al. 2009) these data suggest that either working memory paradigms in rodents are not equivalent to those in humans or that glutamatergic deficits do not cause the working memory deficits evident in schizophrenia. As the animals were performing above criterion prior to the test phase the impairment must reflect an inability to either remember the rule, an inability to apply the rule (presumably due to motor or sensory deficits) or perseverative responding prevailing over correct responding. Previous studies have demonstrated that MK-801 can produce delay-independent deficits in spatial operant tasks such as delayed matching to position (DMTP) at doses of 0.1 and 0.2 mg/kg (Han et al. 2000; Stephens and Cole 1996). The lack of an effect in the present study at 0.1 mg/kg may reflect task differences such as the type of discrimination performed or the naivety of the animals. Whilst this phase shared features of spatial or non-spatial delayed discrimination tasks there is an important distinction: animals in the contemporary design are naïve to the delay. Accordingly, different cognitive strategies and hence domains may be important as animals do not have the opportunity to learn that they need to maintain the representation or to learn a mediating strategy (Chudasama and Muir 1997). This does not of course preclude the use of an unintentional mediation strategy, for example the animals remaining in position by the correct lever whilst waiting for lever presentation.
Of particular interest in the delayed discrimination phase of the battery was the finding that CP 101,606 significantly enhanced performance. Whilst this effect was not dose dependent it corroborates a previous report in which CP 101,606 improved accuracy in DMTP (Higgins et al. 2005). Whilst the ANOVA failed to reach significance, there was also a trend for enhanced performance with planned comparisons following treatment with Ro 25-6981 at 3 mg/kg (p = 0.04) and 5 mg/kg (p = 0.07). These data strongly suggest that selective GluN2B antagonism may indeed improve accuracy in trained animals performing a conditional discrimination. However, the effect was not delay-dependent and so is unlikely to reflect a specific effect directly on working memory: perhaps attentional mechanisms are more directly influenced. Whatever the reason, it is difficult to reconcile enhanced accuracy in the performance of a task at doses that impair acquisition and consolidation of extinction of that same task other than to assume that it illustrates the mechanistic separation between encoding and consolidation of information over time with the information processing needed to successfully complete one individual trial. Enhanced performance could be an artefact of disruption of another behavioural process tending to impair performance, for example by reducing interference.
Phase 3: reversal
Vehicle-treated animals showed very poor performance and were slow to show improvement when the discrimination rule was suddenly reversed and the animals were required to inhibit the response–outcome relationship that had been rewarded over many sessions. MK-801 (0.1 mg/kg), PCP (2.5 mg/kg) and memantine (5 mg/kg) all increased hit rate from the start of the reversal session. Whilst this could represent rapid acquisition of the new discrimination it is more likely to represent a performance artefact resulting from reduced interference from the learned discrimination or, alternatively, an inability to apply the rule such that their performance was simply at, or approaching chance level. Alternatively, animals perseverating on one lever would also lead to performance tending towards 50%. A literature search revealed no previous reports of the effect on NMDA receptor antagonism on the reversal of operant conditional discriminations. However MK-801 has previously been demonstrated to impair reversals in a simple lever-press reversal task (van der Meulen et al. 2003).
In contrast, CP 101,606 produced a different pattern of disruption of the reversal phase such that the hit rate was below that of the control group across the two sessions. This could reflect a stronger representation of the previous discrimination, genuine impaired reversal learning or an artefact of a spurious vehicle group as the performance of this group was indeed higher than in the other studies. However, when the enhanced performance seen in the delayed discrimination phase is considered with CP 101,606, it is possible that the findings indeed represent stronger representation and/or perseveration. Intriguingly, NVP-AAM077 (10 mg/kg) also produced a significant enhancement relative to vehicle control animals in block 3 of the first session only, consistent with enhanced acquisition of the reverse discrimination. Further study is needed to determine whether this represents meaningful or anomalous data.
Phase 4: extinction
The final phase of the battery examined appetitive lever-press extinction in the absence of conditioned stimuli. It should be noted that, as this is a discrete-trial choice procedure, it may alternatively be described as spatial lever-press discrimination since one of the levers remained rewarded. This phase of the battery also shares some similarity to the extra-dimensional shift in an attentional set shifting tasks (Dias et al. 1996; Birrell and Brown 2000) in that the previously ignored dimension (spatial) is now the relevant dimension. However in the current paradigm the irrelevant stimuli are no longer present. There were two phases to the extinction phase—day 1, in which the animals received drug treatment, and day 2 which was drug free. Clearly, vehicle-treated animals learned to inhibit responding on the non-rewarded lever in favour of the rewarded lever on day 1. This memory was consolidated and remembered the following day as animals continued to respond preferentially on the rewarded lever from the start of the session (block 1). The effects of the receptor antagonists on these two phases could be clearly dissociated. MK-801 (0.1 mg/kg) and PCP (2.5 mg/kg) blocked extinction such that the animals continued to respond on the non-rewarded lever. This is consistent with previous data that have demonstrated that MK-801 can block extinction in an appetitive lever-press paradigm (van der Meulen et al. 2003). Other compounds though, were without effect on extinction day 1.
With the exception of ketamine and NVP-AAM077, the effect of the NMDA receptor antagonists on day 2 of extinction was quite striking as significant effects were observed at doses below those which affected omissions or, in the case of MK-801 and PCP, extinction on day 1. This effect was therefore clearly NMDA receptor-mediated and probably reflects the blockade of consolidation of learning as deficits were observed at doses that did not affect day 1 extinction. It is widely accepted that extinction involves new learning as opposed to forgetting per se (Myers and Davis 2002). It is also known that consolidation of extinction of conditioned fear involves the transfer from NMDA receptor-independent to NMDA receptor-dependent learning (Santini et al. 2001) which is consistent with the current findings. Indeed the NMDA/glycine site receptor agonist, D-cycloserine may potentiate extinction of fear conditioning (Walker et al. 2002). An alternative explanation: that these data can be explained by state-dependent learning (Poling et al. 2000), seems unlikely since both ketamine and NVP-AAM077 were essentially without effect.
Carry over effects
The aim of this investigation was to examine the differential effects of NMDA receptor antagonism on learning, memory and cognitive flexibility using a novel consecutive series of behavioural tests. The sequential nature of the tests made it imperative to avoid performance changes in one phase affecting performance in a subsequent phase. The potential for this was mitigated by balancing the treatment groups of each phase in each subsequent phase. In addition, all animals were trained to criterion prior to each new phase and received extra training where necessary. Any animals that failed to reach criterion were excluded from the analysis. Whilst this may not have entirely prevented carry-over effects they would at least have been balanced across treatment groups. An alternative protocol in which animals received the same dose of the same compound throughout the test battery would have clearly led to confounded data. For example, it would have been impossible to determine the effect of NMDA receptor antagonism on later phases of the battery relative to vehicle as animals would have been impaired by the drugs on the initial discrimination. A second alternative of testing drug-naïve subjects independently on each phase of the battery would have required very high numbers of animals: 48 for each phase of the battery—quadrupling the number of animals used in the present study. Where more subtle effects are anticipated, for example in the search for nootropic compounds, animals may receive the same treatment prior to each test session or alternatively may be dosed chronically throughout. It is of note that no carry-over effects were found following treatment with either PCP or MK-801 as sub-chronic administration can produce deficits in tasks such as delay-dependent T maze alternation, social interaction, reversal learning and set shifting (Abdul-Monim et al. 2006; Pratt et al. 2008; Seillier and Giuffrida 2008).
Comparison to human studies
A meta-analysis of published clinical data concluded that ketamine produces robust episodic memory impairments in man evident as deficits in both recognition and recall performance (Morgan and Curran 2006). In contrast, the body of literature suggests only limited, if any, effects on attention and executive function (Morgan and Curran 2006). Krystal et al. (1994) examined the error pattern subjects made whilst performing the Wisconsin card sorting test (WCST) and suggested that ketamine infusion-induced subjects to show deficits in rule learning rather than in executive function per se. This analysis would be in accordance with the current data set as animals were impaired on the discrimination acquisition (rule learning) phase. Furthermore, and consistent with our data, (Krystal et al. 2000) noted that ketamine impaired procedural learning on repeated testing using the WCST but not the expression of a learned rule. Ketamine produced no delay-dependent deficits in the delay phase of the battery and studies of working memory in humans have, in general, demonstrated that ketamine does not affect the maintenance of information in working memory (see Morgan and Curran 2006, for review). In this battery, we were unable to separate unambiguously cognitive from non-specific performance deficits which contrasts to the clear cognitive deficits revealed in clinical studies. It is conceivable that the motoric requirements of human clinical tests are not sufficient to reveal a deficit, and/or that humans are sufficiently motivated to overcome them.
The memantine data are also of particular interest. Memantine and ketamine share many biochemical properties but have divergent therapeutic potential and clinical profile (Johnson and Kotermanski 2006). Memantine is licenced in humans for the treatment of moderate to severe Alzheimer's disease (Witt et al. 2004). It is thought to confer therapeutic benefit without cognitive impairment as it demonstrates the requisite separation between pathological and physiological activation of NMDA receptors (Johnson and Kotermanski 2006). It has been proposed that memantine may provide neuroprotection and enhance cognition through the same mechanism, i.e. restoration of homeostasis in the glutamatergic system (Parsons et al. 2007). Cognitive impairment was demonstrated at low doses in the acquisition phase (10 mg/kg) and consolidation of extinction (5 mg/kg) consistent with the data of Creeley et al. (2006). The minimum dose for neuroprotection in the rat is 10–20 mg/kg (Creeley et al. 2006; Parsons et al. 1999). Acute doses of up to 5 mg/kg produce plasma levels similar to those mediating clinical effects in demented patients and may be considered a therapeutic dose (Parsons et al. 1999). However a number of papers have reported positive effects in models of cognitive impairment such as transgenic mice mimicking some of the biochemical consequences of Alzheimer's disease and lesion-induced deficits in rats (e.g. Creeley et al. 2006; Minkeviciene et al. 2004, 2008; More et al. 2008; Yuede et al. 2007). This discrepancy between the positive effects on cognition in deficit models as opposed to the negative effects in normal animals is potentially a consequence of a hypoglutamatergic state in these animals (Parsons et al. 2007). Consistent with this hypothesis are reports of deficits in recognition memory in normal humans given memantine (Rammsayer 2001). Another study demonstrated that memantine produced no effects on mood, attention, or verbal or visuospatial memory but did impair the acquisition of classical eyeblink conditioning—a non-declarative memory task (Schugens et al. 1997). Overall, the data highlight the importance of evaluating the potential of pro-cognitive effects of drugs in conjunction with manipulations that impair cognition. As approved antipsychotics do not confer cognitive improvement, reversal of the cognitive impairment induced by the NMDA receptor antagonists in this task would not be anticipated if these are a valid model of schizophrenia.
Summary and conclusions
This study forms part of larger body of work in which we have profiled a selection of different classes of NMDA receptor antagonists across a broad range of cognitive and non-cognitive tasks. In the first of these papers, the compounds were profiled over a series of VI schedules of increasing complexity and locomotor activity (Gilmour et al. 2009). Three qualitatively distinct patterns of performance were revealed which neither corroborated nor negated the broad classification suggested by the contemporary findings. MK-801 and PCP showed a similar profile across the battery with marked effects on acquisition, reversal and extinction. Their profile only differed in the delay phase where only PCP produced a hit rate deficit. In the initial phase, the GluN2A-preferring antagonist, NVP-AAM077, stood out as producing only very mild deficits on acquisition and reversal. The GluNR2B preferring antagonists CP 101,606 and Ro 25-6981 produced a highly selective effect on the acquisition phase with only a mild increase in omissions evident on the first day. The compounds showed a similar profile during the extinction phase. The delay and reversal phases revealed CP 101,606 as distinct from the other antagonists suggesting dissociation between GluNR2A- and GluNR2B-dependent behavioural processes. There is the need for GluNR2A ligands with higher selectivity to explore these findings further. Clearly, effects on cognitive parameters could be separated from a direct consequence of drug effects on motor, appetitive or goal-directed instrumental behaviour. The novel test battery was designed to present a series of different cognitive challenges and hence gain multiple indices of behaviour including rule learning, cognitive flexibility, working memory, reference memory, extinction and motor and sensory processing. Animals readily acquired each phase of the battery and showed baseline performance at levels suitable for demonstrating either cognitive enhancement or cognitive impairment.
Translational psychopharmacological studies with NMDA receptor antagonists are hampered by a lack of tools that are available for exploratory research in humans. Preclinically, MK-801 and PCP have been used to model psychosis and cognitive disturbances in animals whilst ketamine alone is used clinically and much less frequently preclinically. The assumption has been that these compounds produce equivalent behavioural phenotypes across species. In our studies, to date, we have seen equivalence in the preclinical profile of MK-801 and PCP but divergence with respect to the effects of ketamine. Since it is much easier to use the licenced pharmaceutical, ketamine than PCP in human psychological and biomarker studies and given that the preclinical data are not equivalent to PCP and MK-801, it would make sense to focus on ketamine as the predominant preclinical tool. However, in our hands, ketamine's non-specific effects were more evident than its cognitive impairing properties across the current rat cognitive battery. Ketamine has a short half-life, making it far from ideal as a preclinical tool when given as a systemic bolus. Controlled intravenous infusion, as is used in human studies, could potentially overcome this problem and compound exposure may be able to be manipulated to produce a more selective cognitive deficit. It would also be interesting to devise human tests with ketamine that, similar to rodent studies, are motor demanding and so ascertain whether motor impairment is also a translational measure.
Whilst it is difficult to extrapolate directly from the available clinical data, the profile of memantine and ketamine was nevertheless essentially consistent with data produced from human studies. This battery may therefore have utility when used in conjunction with rodent models of schizophrenia and Alzheimer's disease. Indeed, it would be interesting to systematically compare CP 101,606 with memantine and ketamine in man as the GluN2B antagonist produced a distinct profile across the battery. Both preclinical and clinical cognitive assays and animal models of schizophrenia and Alzheimer's disease are undergoing intensive re-examination and development. The choice of NMDA receptor antagonist used preclinically and clinically needs careful thought and awareness of their differing properties if assay validation across laboratories and the translation of findings to man is to be successful.
Abbreviations
- ANOVA:
-
analyses of variance
- CNS:
-
central nervous system
- DMTP:
-
delayed matching to position
- HR:
-
hit Rate
- NMDA:
-
N-methyl-D-aspartate
- OR:
-
omission rate
- PCP:
-
phencyclidine
- sem:
-
standard error of the mean
- VI:
-
variable interval
References
Abdul-Monim Z, Reynolds GP, Neill JC (2003) The atypical antipsychotic ziprasidone, but not haloperidol, improves phencyclidine-induced cognitive deficits in a reversal learning task in the rat. J Psychopharmacol 17:57–65
Abdul-Monim Z, Reynolds GP, Neill JC (2006) The effect of atypical and classical antipsychotics on sub-chronic PCP-induced cognitive deficits in a reversal-learning paradigm. Behav Brain Res 169(2):263–273
Abe H, Ishida Y, Iwasaki T (2004) Perirhinal N-methyl-D-aspartate and muscarinic systems participate in object recognition in rats. Neurosci Lett 356(3):191–194
Alexander SP, Mathie A, Peters JA (2008) Guide to Receptors and Channels (GRAC), 3rd edition. Br J Pharmacol 153(Suppl 2):S1–S209
Auberson YP, Allgeier H, Bischoff S, Lingenhoehl K, Moretti R, Schmutz M (2002) 5-Phosphonomethylquinoxalinediones as competitive NMDA receptor antagonists with a preference for the human 1A/2A, rather than 1A/2B receptor composition. Bioorg Med Chem Lett 12:1099–1102
Baldwin AE, Holahan MR, Sadeghian K, Kelley AE (2000) N-methyl-D-aspartate receptor-dependent plasticity within a distributed corticostriatal network mediates appetitive instrumental learning. Behav Neurosci 114(1):84–98
Bannerman DM, Rawlins JN, Good MA (2006) The drugs don't work-or do they? Pharmacological and transgenic studies of the contribution of NMDA and GluR-A-containing AMPA receptors to hippocampal-dependent memory. Psychopharm 188(4):552–566
Barch DM, Berman MG, Eagle R, Jones JH, Jonides J, MacDonald A, Nee DE, Redick TS, Sponheim SR (2009) CNTRICS final task selection: working memory. Schiz Bull 35:136–152
Barker GR, Bashir ZI, Brown MW, Warburton EC (2006a) A temporally distinct role for group I and group II metabotropic glutamate receptors in object recognition memory. Learn Mem 13:178–186
Barker GR, Warburton EC, Koder T, Dolman NP, More JC, Aggleton JP, Bashir ZI, Auberson YP, Jane DE, Brown MW (2006b) The different effects on recognition memory of perirhinal kainate and NMDA glutamate receptor antagonism: implications for underlying plasticity mechanisms. J Neurosci 26:3561–3566
Birrell JM, Brown VJ (2000) Medial frontal cortex mediates perceptual attentional set shifting in the rat. J Neurosci 20:4320–4324
Bliss TV, Collingridge GL (1993) A synaptic model of memory: long-term potentiation in the hippocampus. Nature 361:31–39
Chenard BL, Bordner J, Butler TW, Chambers LK, Collins MA, De Costa DL, Ducat MF, Dumont ML, Fox CB, Mena EE (1995) (1 S, 2 S)-1-(4-hydroxyphenyl)-2-(4-hydroxy-4-phenylpiperidino)-1-propanol: a potent new neuroprotectant which blocks N-methyl-D-aspartate responses. J Med Chem 38:3138–3145
Chudasama Y, Muir JL (1997) A behavioural analysis of the delayed non-matching to position task: the effects of scopolamine, lesions of the fornix and of the prelimbic region on mediating behaviours by rats. Psychopharm 134:73–82
Creeley C, Wozniak DF, Labruyere J, Taylor GT, Olney JW (2006) Low doses of memantine disrupt memory in adult rats. J Neurosci 26(15):3923–3932
Day M, Langston R, Morris RGM (2003) Glutamate-receptor-mediated encoding and retrieval of paired-associate learning. Nature 424(6945):205–209
Di Ciano P, Cardinal RN, Cowell RA, Little SJ, Everitt BJ (2001) Differential involvement of NMDA, AMPA/kainate, and dopamine receptors in the nucleus accumbens core in the acquisition and performance of pavlovian approach behavior. J Neurosci 21:9471–9477
Dias R, Robbins TW, Roberts AC (1996) Primate analogue of the Wisconsin card sorting test: effects of excitotoxic lesions of the prefrontal cortex in the marmoset. Behav Neurosci 110:872–886
Dingledine R, Borges K, Bowie D, Traynelis SF (1999) The glutamate receptor ion channels. Pharmacol Rev 51:7–61
Dunn MJ, Killcross S (2006) Differential attenuation of d-amphetamine-induced disruption of conditional discrimination performance by dopamine and serotonin antagonists. Psychopharm 188:183–192
Fischer G, Mutel V, Trube G, Malherbe P, Kew JN, Mohacsi E, Heitz MP, Kemp JA (1997) Ro 25-6981, a highly potent and selective blocker of N-methyl-D-aspartate receptors containing the NR2B subunit. Characterization in vitro. J Pharmacol Exp Ther 283:1285–1292
Geyer MA, Krebs-Thomson K, Braff DL, Swerdlow NR (2001) Pharmacological studies of prepulse inhibition models of sensorimotor gating deficits in schizophrenia: a decade in review. Psychopharm 156:117–154
Gilmour G, Pioli EY, Dix SL, Smith JW, Conway MW, Jones WT, Loomis S, Mason R, Shahabi S, Tricklebank MD (2009) Diverse and often opposite behavioural effects of NMDA receptor antagonists in rats: implications for “NMDA antagonist modelling” of schizophrenia. Psychopharm 205:203–216
Goldman-Rakic PS (1987) Circuitry of the frontal association cortex and its relevance to dementia. Arch Gerontol Geriatr 6:299–309
Han CJ, Pierre-Louis J, Scheff A, Robinson JK (2000) A performance-dependent adjustment of the retention interval in a delayed non-matching-to-position paradigm differentiates effects of amnestic drugs in rats. Eur J Pharmacol 403(1–2):87–93
Higgins GA, Ballard TM, Enderlin M, Haman M, Kemp JA (2005) Evidence for improved performance in cognitive tasks following selective NR2B NMDA receptor antagonist pre-treatment in the rat. Psychopharm 179:85–98
Hofer E, Doby D, Anderer P, Dantendorfer K (2001) Impaired conditional discrimination learning in schizophrenia. Schizophr Res 51:127–136
Johnson JW, Kotermanski SE (2006) Mechanism of action of memantine. Curr Opin Pharmacol 6:61–67
Kelley AE, Smith-Roe SL, Holahan MR (1997) Response-reinforcement learning is dependent on N-methyl-D-aspartate receptor activation in the nucleus accumbens core. Proc Natl Acad Sci USA 94:12174–12179
Krystal JH, Karper LP, Selbyl JP, Freeman GK, Delaney R, Bremmer JD, Heninger GR, Bowers MB, Charney DS (1994) Subanaesthic effects of the non-competitive NMDA antagonist, ketamine, in humans. Arch Gen Psychiat 51:199–214
Krystal JH, Bennett A, Abi-Saab D, Belger A, Karper LP, D'Souza DC, Lipschitz D, Abi-Dargham A, Charney DS (2000) Dissociation of ketamine effects on rule acquisition and rule implementation: possible relevance to NMDA receptor contributions to executive cognitive functions. Biol Psychiat 47(2):137–143
Lahti AC, Koffel B, LaPorte D, Taminga CA (1995) Subanaesthetic doses of ketamine stimulate psychosis in schizophrenia. Neuropsychopharmacology 13:9–19
Lodge D, Johnson KM (1990) Noncompetitive excitatory amino acid receptor antagonists. Trends Pharmacol Sci 11:81–86
Mansbach RS, Geyer MA (1989) Effects of phencyclidine and phencyclidine biologs on sensorimotor gating in the rat. Neuropsychopharmacology 2:299–308
McKirdy J, Sussmann JE, Hall J, Lawrie SM, Johnstone EC, McIntosh AM (2009) Set shifting and reversal learning in patients with bipolar disorder or schizophrenia. Psychol Med 39:1289–1293
Menniti F, Chenard B, Collins M, Ducat M, Shalaby I, White F (1997) CP-101, 606, a potent neuroprotectant selective for forebrain neurons. Eur J Pharmacol 331:117–126
Milner B (2005) The medial temporal-lobe amnesic syndrome. Psychiatr Clin North Am 28(3):599–611
Minkeviciene R, Banerjee P, Tanila H (2004) Memantine improves spatial learning in a transgenic mouse model of Alzheimer's disease. J Pharmacol Exp Ther 311:677–682
Minkeviciene R, Banerjee P, Tanila H (2008) Cognition-enhancing and anxiolytic effects of memantine. Neuropharmacology 54:1079–1085
More L, Gravius A, Nagel J, Valastro B, Greco S, Danysz W (2008) Therapeutically relevant plasma concentrations of memantine produce significant L-N-methyl-D-aspartate receptor occupation and do not impair learning in rats. Behav Pharmacol 19:724–734
Morgan CJA, Curran HV (2006) Acute and chronic effects of ketamine upon human memory: a review. Psychopharm 188(4):408–424
Morris RG (1989) Synaptic plasticity and learning: selective impairment of learning in rats and blockade of long term potentiation in vivo by the N-methyl-D-asparate receptor antagonist, AP5. J Neurosci 9:3040–3057
Morris RG, Anderson E, Lynch GS, Baudry M (1986) Selective impairment of learning and blockade of long-term potentiation by an N-methyl-D-aspartate receptor antagonist, AP5. Nature 319:774–776
Murray TK, Ridley RM, Snape MF, Cross AJ (1995) The effect of dizocilpine (MK-801) on spatial and visual discrimination tasks in the rat. Behav Pharmacol 6(5–6):540–549
Myers KM, Davis M (2002) Behavioral and neural analysis of extinction. Neuron 36:567–584
Pallares MA, Nadal RA, Silvestre JS, Ferre NS (1995) Effects of ketamine, a noncompetitive NMDA antagonist, on the acquisition of the lever-press response in rats. Physiol Behav 57(2):389–392
Parsons CG, Danysz W, Quack G (1999) Memantine is a clinically well tolerated N-methyl-D-aspartate (NMDA) receptor antagonist—a review of preclinical data. Neuropharmacology 38:735–767
Parsons CG, Stoffler A, Danysz W (2007) Memantine: a NMDA receptor antagonist that improves memory by restoration of homeostasis in the glutamatergic system—too little activation is bad, too much is even worse. Neuropharmacology 53(6):699–723
Petrides M (2000a) Impairments in working memory after frontal cortical excisions. Adv Neurol 84:111–118
Petrides M (2000b) The role of the mid-dorsolateral prefrontal cortex in working memory. Exp Brain Res 133:44–54
Poling A, Byrne T, Morgan T (2000) Stimulus properties of drugs. In: Poling A, Byrne T (eds) Introduction to behavioural pharmacology. Context Press, Reno, US, pp 141–166
Pratt JA, Winchester C, Egerton A, Cochran SM, Morris BJ (2008) Modelling prefrontal cortex deficits in schizophrenia: implications for treatment. Br J Pharmacol 153(Suppl 1):S465–S470
Rammsayer TH (2001) Effects of pharmacologically induced changes in NMDA-receptor activity on long-term memory in humans. Learn Mem 8:20–25
Santini E, Muller RU, Quirk GJ (2001) Consolidation of extinction learning involves transfer from NMDA-independent to NMDA-dependent memory. J Neurosci 21(22):9009–9017
Schugens MM, Egerter R, Daum I, Schepelmann K, Klockgether T, Loschmann PA (1997) The NMDA antagonist memantine impairs classical eyeblink conditioning in humans. Neurosci Lett 224:57–60
Seillier S, Giuffrida A (2008) Evaluation of NMDA recoptor models of schizophrenia: divergences in the behavioural effects of sub-chronic PCP and MK-801. Behav Brain Res 140(1–2):1–47
Sloan HL, Good M, Dunnett SB (2006) Double dissociation between hippocampal and prefrontal lesions on an operant delayed matching task and a water maze reference memory task. Behav Brain Res 171:116–126
Stephens DN, Cole BJ (1996) AMPA antagonists differ from NMDA antagonists in their effects on operant DRL and delayed matching to position tasks. Psychopharm 126:249–259
Tonkiss J, Rawlins JN (1991) The competitive NMDA antagonist AP5, but not the non-competitive antagonist MK801, induces a delay-related impairment in spatial working memory in rats. Exp Brain Res 85:349–358
Tricklebank MD, Singh L, Oles RJ, Preston C, Iverson SD (1989) The behavioural effects of MK-801: a comparison with antagonists acting non-competitively and competitively at the NMDA receptor. Eur J Pharmacol 167:127–135
van der Meulen JA, Bilbija L, Joosten RN, de Bruin JP, Feenstra MG (2003) The NMDA-receptor antagonist MK-801 selectively disrupts reversal learning in rats. Neuroreport 14:2225–2228
Walker DL, Ressler KJ, Lu KT, Davis M (2002) Facilitation of conditioned fear extinction by systemic administration or intra-amygdala infusions of D-cycloserine as assessed with fear-potentiated startle in rats. J Neurosci 22:2343–2351
Winters BD, Bussey TJ (2005) Glutamate receptors in perirhinal cortex mediate encoding, retrieval, and consolidation of object recognition memory. J Neurosci 25:4243–4251
Witt A, Macdonald N, Kirkpatrick P (2004) Memantine hydrochloride. Nat Rev Drug Discov 3:109–110
Yuede CM, Dong H, Csernansky JG (2007) Anti-dementia drugs and hippocampal-dependent memory in rodents. Behav Pharmacol 18:347–363
Acknowledgements
The authors would like to gratefully acknowledge the technical assistance of Yvonne Thomas, Laura Cross and Alex Harper.
Author information
Authors and Affiliations
Corresponding author
Electronic supplementary material
Below is the link to the electronic supplementary material.
ESM1
(DOC 233 kb)
Rights and permissions
About this article
Cite this article
Dix, S., Gilmour, G., Potts, S. et al. A within-subject cognitive battery in the rat: differential effects of NMDA receptor antagonists. Psychopharmacology 212, 227–242 (2010). https://doi.org/10.1007/s00213-010-1945-1
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00213-010-1945-1