
Abstract
Rationale
The novel antipsychotic aripiprazole in use for treatment of schizophrenia is a partial agonist at dopamine D2 receptors with actions at a variety of other receptors as well. Cocaine is believed to exert an important part of its rewarding effect by increasing extracellular levels of dopamine that subsequently act at dopamine D2 receptors.
Objectives
As a partial agonist, aripiprazole may antagonize effects at D2 receptors and we accordingly tested whether aripiprazole could antagonize self-administration of cocaine. Because D2-like receptor agonists are self-administered, a D2 receptor partial agonist like aripiprazole might itself be reinforcing. Thus, we also assessed whether mice would acquire self-administration of aripiprazole.
Materials and methods
A single session, mouse self-administration procedure was used.
Results
Oral pretreatment with aripiprazole dose-dependently decreased cocaine self-administration under a fixed ratio 1 schedule at the peak cocaine dose (0.03 mg/kg/infusion), reaching significance at 0.2 and 0.4 mg/kg of aripiprazole. Using 0.4 mg/kg, aripiprazole decreased rates of cocaine self-administration without shifting the peak of the dose-response function. There was no effect of aripiprazole per se, suggesting that its inhibitory action was due to effects on cocaine self-administration rather than non-specific motor effects. Aripiprazole was not found to be self-administered in the tested dose range (0.0003–0.3 mg/kg/infusion). The three highest doses (0.03, 0.1, and 0.3 mg/kg/infusion) even caused significant decreases in nose-poking activity, possibly due to extrapyramidal side effects.
Conclusions
These data are consistent with a potential role for aripiprazole in treatment of cocaine addiction without abuse potential per se.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
Cocaine addiction is a serious health problem for modern society with no effective treatment available. Cocaine is believed to cause a substantial part of its rewarding effect by inhibiting the reuptake of dopamine (Ritz et al. 1987; Bannon 2005; Riddle et al. 2005), thereby increasing dopamine binding to postsynaptic dopamine receptors. In addition, there is also evidence for a role of the serotonin system in mediating the rewarding effects of cocaine (see Müller and Huston 2006 and Müller et al. 2007 for reviews). As for the dopamine system, several studies indicate that the D2 receptor is centrally involved in mediating the rewarding effect of cocaine. For instance, D2-like receptor agonists are self-administered in animals trained to self-administer cocaine (Woolverton et al. 1984; Wise et al. 1990; Caine and Koob 1993; Grech et al. 1996; Nader and Mach 1996; Sinnott et al. 1999; Ranaldi et al. 2001) and pretreatment with D2-like agonists cause a leftward shift in cocaine self-administration, dose-effect functions (Caine and Koob 1995; Caine et al. 1999, 2000; Barrett et al. 2004). D2-like receptor agonists also facilitate reinstatement of responding previously reinforced by cocaine (Self et al. 1996; Khroyan et al. 2000). In contrast, pretreatment with D2-like receptor antagonists shift the dose-effect curve to the right in animals self-administering cocaine, suggesting surmountable antagonism of cocaine’s reinforcing effects (Bergman et al. 1990; Mello and Negus 1996 for review).
Another line of evidence implicating D2 receptors comes from studies showing that chronic cocaine self-administration can induce prolonged reduction in the density of striatal D2-like binding sites in monkeys (Moore et al. 1998; Nader et al. 2002). Similarly, positron emission tomography data have shown reduced striatal D2-like binding sites in cocaine abusers (Volkow et al. 1993). Perhaps, paradoxically, there is evidence that low D2 receptor density is associated with an increased propensity to self-administer cocaine in rats and monkeys, and an increased “liking” of methylphenidate in humans (Volkow et al. 1999, 2002; Morgan et al. 2002; Nader et al. 2006; Dalley et al. 2007).
Aripiprazole, a novel atypical antipsychotic, is primarily known as a partial agonist at dopamine D2 receptors with low propensity for extrapyramidal side effects (Kikuchi et al. 1995; Semba et al. 1995; Lawler et al. 1999; Burris et al. 2002; Nakai et al. 2003). However, aripiprazole may also exert effects at a variety of other non-D2 receptors, including partial agonist actions at 5-HT1a and 5-HT2c receptors and antagonist actions at 5-HT2a serotonin receptors (Lawler et al. 1999; Shapiro et al. 2003; Marona-Lewicka and Nicholas 2004; Kleven et al. 2005; Auclair et al. 2006; Stark et al. 2007). Dopamine D2 partial agonists have received some attention as potential medication for cocaine abuse and dependence as they can act as an antagonist in states of high D2 receptor stimulation, such as when cocaine is present, while acting as an agonist in states of low dopaminergic tone, such as in a cocaine withdrawal state (see Pulvirenti and Koob 2002 for review). Consistent with this concept, aripiprazole has been shown to reduce cocaine craving (Brown et al. 2005) and abuse in schizophrenic patients (Beresford et al. 2005) and to decrease subject-rated effect of d-amphetamine (Lile et al. 2005; Stoops et al. 2006). To further explore this therapeutic potential of aripiprazole, we examined the effect of aripiprazole on cocaine self-administration in a well-characterized, single-session procedure in mice (Criswell and Ridings, 1983; Kuzmin et al. 1992; Rasmussen and Swedberg, 1998; Rasmussen et al. 2000; Fink-Jensen et al. 2003; Paterson et al. 2003; Blokhina et al. 2005). Because D2-like receptor agonists are self-administered (Caine and Koob 1993; Grech et al. 1996; Sinnott et al. 1999; Ranaldi et al. 2001), a D2 partial agonist like aripiprazole might, in addition to blocking drug abuse, have rewarding effects per se. To address this issue, we also assessed whether drug-naïve mice would acquire self-administration of aripiprazole.
Materials and methods
Compounds
Methyl (2R,3S)-3-benzoyloxy-8-methyl-8-azabicyclo[3.2.1]octane-2-carboxylate hydrochloride (cocaine; synthesized at Danish Hospital Pharmacies and H. Lundbeck A/S) and methyl-alpha-phenyl-2-piperidineacetate hydrochloride (methylphenidate; synthesized at H. Lundbeck A/S) were dissolved in 0.9% isotonic saline. 7-[4-[4-(2,3-Dichlorophenyl)-1-piperazinyl]butoxy]-3,4-dihydrocarbostyril (Aripiprazole; synthesized at H. Lundbeck A/S) was suspended in vehicle consisting of isotonic 10% 2-hydroxy-propyl-beta-cyclodextrin (HPBC) + 1 M CH3SO3H (methanesulfuric acid) for both oral (p.o.) and intravenous (i.v.) administrations.
Animals
Male NMRI mice (Taconic, DK or Charles River, Germany) were used for all experiments. The animals were group-housed under standard laboratory conditions in boxes of up to ten mice per cage, on a 12 h light–dark cycle with free access to standard rodent chow and tap water. All experiments were performed between 900 h and 1600 h. Animals were only used for one type of experiment listed below. The Guidelines for the Care and Use of Mammals in Neuroscience and Behavioral Research as well as the guidelines of the Animal Experimentation Inspectorate of Denmark were followed in all experiments.
Acute self-administration
The self-administration procedure was described previously (Fink-Jensen et al. 2003). Mice weighing 20–22 g were used. The self-administration apparatus consisted of transparent plastic boxes (8 × 8 × 8 cm) with a centered frontal nose-poke hole (12 mm diameter) 1 cm above the floor and a centered posterior vertical opening (width 5 mm) through which the tail extended. Dual photocells projected an infrared beam 1 mm in front of the nose-poke hole. Eight mice were tested at the same time, with interposed shields, preventing the mice from seeing each other during the experiment.
Immediately before being placed in the test boxes, the mice were left for approximately 3 min 30–35 cm below a 150 W infrared light bulb to induce vasodilatation of their tails, thus, facilitating the insertion of the cannula into the tail vein. The tail was fixed onto a stripe of double-sided adhesive tape (Tesafix® 50X19 4964, Lynge Papir & Kontor, Silkeborg, Denmark). The tape also fixed the tail at its base as well as the vein cannula at the cannula insertion point. A nose-poke interrupted the infrared beam, thus, activating the photocell connected to an interface (SG 502, Med Associates Inc., East Fairfield, VT, USA) and, thereafter, a syringe pump (PHM-100A, Med Associates Inc.) holding a 2 ml syringe (Omnifix® Solo, B. Braun, Copenhagen, Denmark) connected by a 75 cm PVC tubing (0.9 mm inner diameter, Original-Perfusor Leitung MR, B. Braun; Germany) to a 27 G infusion needle (0.4 × 40 mm, Sterican®, B. Braun). A backcheck valve (Rüchschlagsventile, B. Braun) between needle and tubing prevented reflux of blood. A fixed ratio 1, FR-1, schedule was used, with no time-out between nose-poke and infusion so that each nose-poke induced the i.v. infusion of 1.4 μl of drug. A computer and self-administration software (Ellegaard Systems, Denmark) were used to control infusions and to record nose-poking behavior.
After placing the mouse in the self-administration box for a 10-min habituation period during which nose-poking did not induce infusions, one priming infusion was given by the experimenter immediately before starting a 30-min session. Immediately after the session, the correct placement of the infusion needle was verified by manual infusion of the tested drug by an experimenter blind as to the number of nose-pokes produced, and the animals were quickly killed. Mice were excluded from further analysis if they had not produced at least five nose-pokes during the self-administration session or if correct placement of the infusion needle could not be verified.
Three self-administration experiments were performed. In the first experiment, the mice received p.o. administration of aripiprazole (0.05, 0.1, 0.2 or 0.4 mg/kg) or corresponding vehicle. One hour later, the animals were subjected to the self-administration procedure described above with access to i.v. administration of vehicle or cocaine at 0.03 mg/kg/infusion, previously shown to be the optimal dose for self-administration (Fink-Jensen et al. 2003). In the second experiment, the mice received p.o. administration of 0.4 mg/kg aripiprazole or corresponding vehicle. One hour later, the animals were subjected to the self-administration procedure described above with access to i.v. administration of cocaine (0.003, 0.01, 0.03, 0.1 or 0.3 mg/kg/infusion) or vehicle. In the third experiment, the mice were subjected to the self-administration procedure without prior administration of drug. The animals had access to i.v. self-administer cocaine (0.003, 0.03, 0.3 mg/kg/infusion), aripiprazole (0.0003, 0.001, 0.003, 0.01, 0.03, 0.1, 0.3 mg/kg/infusion), or corresponding vehicles.
Methylphenidate antagonism
Lower doses of the indirect dopamine agonist methylphenidate causes hyperactivity that is considered to involve predominant activation of mesolimbic dopamine pathways. At higher doses, methylphenidate induces stereotyped gnawing behavior believed to selectively involve the nigrostriatal dopamine pathway (Nielsen et al. 1997). Antipsychotics block both methylphenidate-induced hyperactivity and gnawing; however, the ratio between ED 50 for blocking gnawing and ED 50 for blocking hyperactivity (i.e., the mesolimbic ratio) is considered a useful ratio for predicting selectivity of a compound to antagonize mesolimbic dopamine neurotransmission without causing extrapyramidal side effects (Nielsen et al. 1997). The ability of aripiprazole to antagonize effects of methylphenidate was measured both in locomotor and gnawing tests in male NMRI mice (20–25 g). The experiments were performed in a well-lit and undisturbed room with white-noise background. The mice were habituated to the test room at least overnight.
Methylphenidate-induced hyperactivity
Test cages (20 cm × 32 cm) were equipped with 5 × 8 infrared light sources and photocells spaced by 4 cm. The light beams crossed the cage 1.8 cm above the bottom of the cage. Mice received one dose of aripiprazole (0.04, 0.16, 0.31, or 1.3 mg/kg) or vehicle p.o. 60 min before activity recordings (n = 8–16/group). After oral administration, the mice were allowed to habituate to the activity cage for 60 min. Then methylphenidate (7.5 mg/kg s.c., 10 ml/kg) or vehicle was injected, the mice were returned to the activity cage, and locomotor activity recording continued for 60 min. Locomotor activity was thus recorded for a total of 2 h, in 5-min bins (i.e., a total of 24 bins). Only consecutive beam breaks (i.e., not repeated breaks of a same beam) were recorded as locomotor activity.
Methylphenidate-induced gnawing
Perspex observation cages without bottom or lid were used for evaluating methylphenidate-induced gnawing behavior. The cages consisted of five sections, each measuring 11 × 25 cm and were placed on corrugated paper, the corrugations facing upwards. Mice received one oral dose of aripiprazole (0.08, 0.31, 1.3, 5, or 10 mg/kg; n = 8–16/group) or vehicle (n = 24) 60 min prior to evaluation. Then methylphenidate (60 mg/kg s.c., 10 ml/kg) or vehicle was injected, and the mice were placed in individual observation boxes for 60 min; after which, the gnawing score was determined according to a method modified from Nielsen et al. (1997). A transparent Plexiglas plate of the same size as the test box and divided into 12 squares (2 × 6) was used for scoring of gnawing. The degree of gnawing was measured as the number of squares with bite marks. Only a total number of bite marks above ten was considered as a gnawing response. Squares with bite marks (even only one) was then counted and the gnawing degree expressed as fractions of 12, e.g., 1/12 or 4/12, where 1 and 4 indicate the number of squares with bite marks.
In vivo binding to D2 receptors ([3H]-raclopride)
Mice (20–25 g) were treated with aripiprazole p.o. 60 min before measuring radioactivity in the brain. At 50 min after drug administration, the animals received 3.0 μCi [3H]-raclopride (specific activity 60 Ci/mmol; #NET-975, PerkinElmer, Inc., Denmark) i.v. via the tail vein. In vivo binding was carried out according to Andersen et al. (1986) with a few modifications (Kapur et al. 2003). Ten minutes after radioligand injection, the animals were killed by cervical dislocation, their brains were quickly removed, and striatum dissected out and homogenized in 3.5 ml ice-cold buffer (50 mM KPO4, pH 7.4). Homogenate (0.5 ml) was filtered through 0.1% polyethyleneimine-soaked Whatman GF/C filters. This entire process was completed within a time-span of only 60 s after decapitation. Filters were washed twice with 5 ml ice-cold buffer and counted in a scintillation counter. A group of saline-treated animals was used to determine total and non-specific [3H]-raclopride binding, measuring radioactivity in the striatum and cerebellum, respectively. The homogenate was measured for protein content by the bicinchoninic acid protein determination assay (Smith et al. 1985).
Statistics
Data were analyzed using Statistical Package for the Social Sciences or GraphPad Prism computer software. One animal from the first experiment was excluded from the experiment as an extreme value defined as more than three interquartile ranges above or below the 25th or 75th percentile, respectively (SPSS Inc. 2004). A high variance was seen at effective cocaine doses (0.03, 0.1 mg/kg/infusion) as compared to lower variance in saline and non-self-administered cocaine doses. The higher variance at effective cocaine doses was probably due to the fact that some animals failed to learn the self-administration paradigm during the single session. To counter this variance inhomogeneity, the number of nose-pokes was square root transformed before statistical analyses. Logarithmic transformation was also tried, confirming the results. In the first and third self-administration experiments, data were analyzed using one-way analysis of variance (ANOVA) followed by post-hoc least significant difference (LSD) t-tests. In the second experiment, two-way ANOVA was used. Subsequently, after significant treatment effect of cocaine, data were analyzed using one-way ANOVA followed by post-hoc LSD t-tests. After significant aripiprazole treatment effect, we used post-hoc t-tests to determine the cocaine doses at which the treatment effect occurred. In addition, for the second self-administration experiment, polynomial regression of second order was performed on logarithm-transformed cocaine doses to further analyze the inverted U-shaped cocaine dose-response curve. The dose at which the nose-poking activity was at its maximum (vertex) was x max = −b/(2a). To assess whether aripiprazole shifted the cocaine dose-response curve to the right or left, a likelihood ratio test was carried out of the hypothesis that x max did not depend on whether aripiprazole pretreatment was given (mle, software package R, version 2.4; R Development Core Team 2006). The in vivo ED 50 values of the methylphenidate experiments were estimated based on a sigmoid dose-response curve using non-linear regression. D2 receptor occupancy (%) was calculated as: occupancy = [compound]/(ED 50 + [compound]). Data were presented as means ± standard errors of the means. The level of significance was p < 0.05.
Results
Acute self-administration
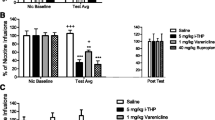
In the first experiment (Fig. 1), oral administration of aripiprazole significantly and dose-dependently decreased self-administration of the 0.03 mg/kg/infusion cocaine dose (F(5,84)=2.72, P = 0.025), reaching significance at the two highest aripiprazole doses (0.2 and 0.4 mg/kg; P = 0.046 and P = 0.024, respectively).

Aripiprazole pretreatment p.o. dose-dependently decreased nose-poking for cocaine at the optimal i.v. self-administration dose of cocaine (0.03 mg/kg/infusion) in mice, reaching significance at 0.2 and 0.4 mg/kg. Numbers on columns indicate the number of animals in the group. **P < 0.01, *P < 0.05 vs. saline i.v. preceded by vehicle p.o., ***P < 0.05 vs. cocaine i.v. preceded by vehicle p.o.; post-hoc LSD t-tests following significant one-way ANOVA
In the second experiment (Fig. 2), two-way ANOVA revealed a significant effect of treatment with cocaine (F(5,155)=3.06, P = 0.012) as well as aripiprazole (F(1,155)=20.11, P < 0.001) without interaction (P = 0.47). In vehicle p.o.-pretreated mice, cocaine appeared to be self-administered i.v. according to an inverted U-shaped curve, reaching maximum at 0.03 mg/kg/infusion consistent with previous work (Fink-Jensen et al, 2003). Post-hoc LSD t-test following significant one-way ANOVA (F(5,77)=2.73, P = 0.025) confirmed that doses of both 0.03 and 0.1 mg/kg/infusion were self-administered as compared to saline i.v. (P = 0.002 and P = 0.02, respectively). Polynomial regression of second order confirmed the inverted U-shaped form of the curve (F(1,80)=6.38, P = 0.014; y = −2.0 x 2 + −5.9 x + 7.6). One-way ANOVA for aripiprazole-pretreated animals administered various doses of cocaine i.v. was far from significant (F(5,73)=0.89, P = 0.492), indicating that cocaine was not self-administered as compared to saline in aripiprazole-pretreated mice. Consistent with this interpretation, polynomial regression showed no evidence that the data of aripiprazole-pretreated mice could be fitted to a parabola (F(1,76)=0.42, P = 0.52). In accordance with the above-mentioned treatment effect of aripiprazole in the two-way ANOVA, the 0.4-mg/kg aripiprazole dose significantly reduced the nose-poking frequency of i.v.-administered cocaine at the two cocaine doses self-administered significantly as compared to saline i.v. (0.03 and 0.1 mg/kg/infusion; t(33)=2.57, P = 0.015 and t(22)=2.09, P = 0.049, respectively) as well as at the 0.003 mg/kg/infusion (t(25)=2.49, P = 0.020) cocaine dose (Fig. 2). However, aripiprazole p.o., in comparison with vehicle p.o., did not significantly decrease nose-poking when saline was present as the i.v. reinforcer.

Inverted U-shaped i.v. cocaine self-administration curves in mice pretreated with aripiprazole at 0.4 mg/kg or vehicle p.o. Cocaine’s dose-effect curve was shifted down. There was no effect of aripiprazole per se, suggesting that its inhibitory action is due to an effect on cocaine self-administration rather than a non-specific effect on motor behaviour. Number on columns indicates the number of animals in the group. **P < 0.01, *P < 0.05 vs. saline i.v. pretreated with vehicle p.o., post-hoc LSD t-tests following significant one-way ANOVA after significant cocaine treatment effect in two-way ANOVA. ***P < 0.05 vs. corresponding p.o. vehicle-pretreated group, post-hoc t-test following significant aripiprazole treatment effect in two-way ANOVA
Like in vehicle p.o.-pretreated mice, the highest nose-poking rate was observed at 0.03 mg/kg/infusion cocaine in aripiprazole p.o.-pretreated animals. To confirm that aripiprazole did not change the potency of cocaine, the vertices of the dose-response curves for cocaine with or without aripiprazole at 0.4 mg/kg were compared after fitting to parabolas. A likelihood ratio test showed that the vertex of the cocaine dose-response curve of aripiprazole treated mice did not shift significantly to the right or left (P = 0.62), indicating that aripiprazole did not change the potency of cocaine.
In the third experiment (Fig. 3), the 0.03 mg/kg/infusion dose of cocaine was self-administered i.v. at a significantly higher frequency than saline (P = 0.001, LSD post-hoc t-test after significant one-way ANOVA; F(3,43)=5.19, P = 0.004), consistent with the previous experiments. In contrast, i.v. aripiprazole did not cause higher nose-poking frequencies as compared to vehicle at any dose in the interval tested. However, at the three highest doses, i.v. administration of aripiprazole was associated with a significant decrease in nose-poking behavior as compared to vehicle i.v.

Nose-poking rates for i.v. self-administration of cocaine (n = 10–14) or aripiprazole (n = 5–9) as compared to corresponding vehicle groups (n cocaine = 11, n aripiprazole = 14). Unlike cocaine, aripiprazole was not self-administered i.v. in the tested dose range but showed a significant decrease in nose-poking at the three highest doses tested as compared to vehicle. ***P < 0.001, **P < 0.01, *P < 0.05 vs. vehicle i.v., LSD post-hoc t-test following significant one-way ANOVA
Methylphenidate antagonism
Aripiprazole reduced the hyperactivity induced by the dopamine-stimulating compound methylphenidate (ED 50 = 0.31 mg/kg). The efficacy of aripiprazole at reversing methylphenidate-induced gnawing was lower (ED 50 = 1.6 mg/kg), with a mesolimbic efficacy ratio of 5.2.
In vivo binding to D2 receptors ([3H]-raclopride)
The ED 50 for D2 receptor occupancy at 60 min after oral administration of aripiprazole was 0.68 mg/kg (95% confidence interval, 0.52–0.89 mg/kg; Fig. 4). Based on the fitted sigmoid dose-response curve, the 0.4 mg/kg dose of aripiprazole was associated with approximately 30% D2 receptor occupancy.

D2 dopamine receptor occupancy (%) in the mouse striatum as measured by [3H]-raclopride binding following oral administration of aripiprazole 60 min before decapitation of the animals
Discussion
The present study shows that aripiprazole dose-dependently reduced acute cocaine self-administration at the dose of cocaine resulting in maximal nose-poking frequency (0.03 mg/kg/infusion) in drug-naïve mice. In addition, the dose-response curve for cocaine self-administration was shifted down in animals pretreated with 0.4 mg/kg aripiprazole, without indication of a leftward or rightward shift. This suggests that aripiprazole reduced the efficacy of cocaine as a reinforcer, rather than producing surmountable antagonism (i.e., reducing its potency), which would be manifested as a rightward shift of the dose-effect curve. Indeed, in animals treated with aripiprazole, no dose of cocaine was self-administered significantly above saline levels, and because the highest nose-poking frequency occurred at the 0.03 mg/kg/infusion cocaine dose, it seems unlikely that higher doses of cocaine than the ones tested would have maintained self-administration. This profile supports the concept that aripiprazole could be useful as an abuse medication in humans, as it suggests that its blockade of cocaine’s effect may not be easily overcome by administering higher doses of cocaine. These results are in seeming contrast with a previous report using another D2 receptor agonist, terguride, which shifted cocaine’s self-administration dose-effect curve to the right (Pulvirenti et al. 1998). In partial agreement with our results, it was recently reported that pretreatment with aripiprazole in rats shifted the dose-response function for self-administration of methamphetamine down and to the right (Wee et al. 2007b). The reason for this partial discrepancy remains unclear except for the obvious mouse vs. rat, single session vs. chronic procedure, and cocaine vs. methamphetamine differences in the two studies involving aripiprazole.
Our data are consistent with previous investigations suggesting that aripiprazole may block reinforcing effects of psychostimulants. A recent study showed that aripiprazole blocked cocaine’s ability to reinstate lever pressing previously associated with cocaine reinforcement in rats (Feltenstein et al. 2007). The presently used acute cocaine self-administration model might not reflect the abuse situation in subjects with chronic cocaine abuse. Indeed, Feltenstein et al. (2007) reported that pretreatment with aripiprazole i.p. failed to affect i.v. cocaine self-administration in their chronic rat model. Further studies are needed to address this issue. One reason for the lack of effect in the paradigm of Feltenstein et al. (2007) could be that they used only one dose of cocaine. The testing of different doses of cocaine might have revealed an effect of aripiprazole. In line with our results, clinical studies have reported that aripiprazole reduced cocaine craving and decreased the subject-rated effect of d-amphetamine (Beresford et al. 2005; Brown et al. 2005; Lile et al. 2005; Stoops et al. 2006). Taken together, these findings support the potential usefulness of aripiprazole in the management of psychostimulant abuse.
Aripiprazole at a dose of 0.4 mg/kg per se did not significantly affect nose-poking for saline, indicating that it did not produce strong non-specific rate-decreasing effects that may account for the reduction in cocaine self-administration. Consistent with this interpretation, previous studies indicate that aripiprazole may disrupt psychostimulant self-administration with a greater potency than for food-maintained behavior. For example, aripiprazole prolonged the time required for rats to earn 20 casein pellet reinforcers only at a relatively high dose, 2.5 mg/kg, but not at doses up to 0.75 mg/kg (Schwabe and Koch 2007). Doses up to 0.75 mg/kg aripiprazole i.p. also did not reduce the breaking point in a progressive ratio schedule of food reinforcement in the same study, while decreased responding in rat self-administration of saline using 1–10 mg/kg i.p. aripiprazole was reported in another rat study (Wee et al. 2007b). Similarly, aripiprazole pretreatment failed to alter food-primed reinstatement of lever pressing previously associated with food reinforcement at doses (0.25–1 mg/kg, i.p.) that blocked cocaine-primed reinstatement in rats trained to self-administer cocaine (Feltenstein et al. 2007). In this respect, aripiprazole may be superior to the previously tested D2 receptor partial agonists, terguride and SDZ 208-911. Thus, terguride was found to decrease food-maintained behavior more potently than cocaine self-administration in squirrel monkeys, and SDZ 208-911 actually facilitated reinstatement of cocaine-associated but not food-associated, lever pressing (Weissenborn et al. 1996; Platt et al. 2003). Further comparisons of drug-and non-drug-maintained behaviors are needed to substantiate the apparent superiority of aripiprazole relative to other D2 receptor partial agonists. Perhaps, functional selectivity of aripiprazole (Urban et al. 2007) and other partial D2 agonists at activating different D2 receptor-mediated signalling pathways could be involved.
The methylphenidate hyperactivity model of the present study is believed to reflect activation of mesolimbic regions while methylphenidate-induced gnawing is a model related to action on the nigrostriatal dopamine system (Nielsen et al. 1997). The fact that aripiprazole required five times higher doses for antagonizing methylphenidate-induced gnawing (ED 50 = 1.6) than for reversing methylphenidate-induced hyperactivity (ED 50 = 0.31), i.e., the mesolimbic efficacy ratio, indicates a preferential action of aripiprazole in the mesolimbic dopamine system at doses used in the self-administration study. For comparison, the atypical antipsychotic risperidone and clozapine have mesolimbic efficacy ratios of 1.3 and 9.0, respectively (unpublished data). However, surprisingly, the 0.4 mg/kg aripiprazole dose effective at blocking cocaine self-administration of the present study was associated with only approximately 30% D2 receptor occupancy. Nonetheless, the ED 50 for D2 occupancy (0.68 mg/kg) of the present study was practically identical with that reported in a rat study (0.7 mg/kg) after subcutaneous administration of aripiprazole (Natesan et al. 2006). The fact that blocking of cocaine self-administration by aripiprazole in the present study appeared to be associated with relatively low D2 occupancy suggests that treatment for cocaine abuse could be achieved without significant extrapyramidal side effects.
In order to address the possibility that aripiprazole may have abuse potential in itself, we also evaluated acquisition of i.v. aripiprazole self-administration in drug-naïve mice. We found that aripiprazole was not self-administered i.v., as compared to vehicle, over a wide dose range. This suggests that aripiprazole does not possess intrinsic abuse potential. This result is consistent with the lack of self-administration of other D2 receptor partial agonists in monkeys trained to self-administer cocaine (Pulvirenti et al. 1998; Ranaldi et al. 2001). D2 receptor partial agonists also did not substitute for cocaine in discrimination assays, suggesting that they may not mimic cocaine’s subjective effects (Callahan and Cunningham 1993; Tidey and Bergman 1998). In fact, we saw a decrease in nose-poking relative to vehicle at the three highest doses of aripiprazole tested. This could be due to sedative-like, aversive, or cataleptogenic effects. The lowest dose of aripiprazole associated with a significant decrease in nose-poking (0.03 mg/kg/infusion) during the 30-min test period would roughly correspond to administering a total dose of 2.3 mg/kg. This self-administered dose was clearly above the ED 50 (ED 50 = 1.6 mg/kg) shown in the present study for reversal of methylphenidate-induced gnawing. Because antagonism of methylphenidate-induced gnawing is believed to reflect propensity to cause extrapyramidal side effects (Nielsen et al. 1997), this indicates that significant decreases in nose-poking at the three highest doses of aripiprazole self-administered i.v. could be due to extrapyramidal side effects mediated by blockade of D2 receptors. This is also consistent with two previous studies reporting cataleptogenic effects of aripiprazole at doses from 3 to 10 mg/kg in mice (Nakai et al. 2003; Bardin et al. 2005). It is noteworthy that this seems to represent a species difference between rats and mice because several investigations have failed to induce catalepsy in rats at doses up to 30 or 40 mg/kg i.p. (Semba et al. 1995; Kleven et al. 2005; Natesan et al. 2006). Thus, aripiprazole appears to have very low abuse liability, including at higher doses than might be used clinically (which may actually have aversive properties).
In addition to being a partial agonist at dopamine D2 receptors, aripiprazole appears to act as a partial agonist at dopamine D3 receptors with an affinity similar to that at D2 receptors (DeLeon et al. 2004; Burstein et al. 2005). However, D3 receptor partial agonists did not alter cocaine self-administration in rats (Pilla et al. 1999; Gyertyán et al. 2007) or apes (Beardsley et al. 2001), and D3 receptor partial agonists are not self-administered (Pilla et al. 1999). In general, the D3 receptor seems to be involved in other aspects of addiction than the direct reinforcing effects of i.v. self-administration of drugs (Le Foll et al. 2005). Thus, it is not likely that activity at D3 receptors mediated effects observed in the present study.
Aripiprazole also targets serotonin receptors, 5-HT1a, 5-HT2a, and 5-HT2c as partial agonist and/or antagonist (Lawler et al. 1999; Shapiro et al. 2003; Marona-Lewicka and Nichols 2004; Kleven et al. 2005; Auclair et al. 2006; Stark et al. 2007), and the serotonin system has also been implicated in mediating the psychostimulant effects of cocaine via actions at these receptors (Müller and Huston 2006; Müller et al. 2007). Aripiprazole has approximately ten times lower affinity for 5-HT1a receptors than for D2 receptors (DeLeon et al. 2004). Because the 0.4 mg/kg aripiprazole dose of the present study resulted in approximately 30% D2 receptor occupancy, occupancy at 5-HT1a receptors could be considerably lower. Thus, although a role for 5-HT1a receptors cannot be ruled out, it is less likely that aripiprazole exerted significant effect in the present study via 5-HT1a receptors. A previous study in rats also showed that aripiprazole displayed substantially lower occupancy at 5-HT2a receptors than at D2 receptors at the effective dose used in the present experiment (Natesan et al. 2006), suggesting that 5-HT2a receptors did not contribute importantly to mediating effects of aripiprazole in the present study. Aripiprazole has lower affinity to 5-HT2c receptors than to 5-HT2a (Shapiro et al. 2003; Stark et al. 2007), suggesting that effects at 5-HT2c receptors are probably also of minor importance. Effects at other receptors cannot completely be ruled out at present, for instance, aripiprazole has moderate affinity to alpha-1 adrenergic receptors (Shapiro et al. 2003), and alpha-1 adrenergic receptors were recently implicated in motivation for cocaine self-administration in rats (Wee et al. 2007a). However, again, the substantially lower affinity to alpha-1 adrenergic receptors than to D2 receptors suggests that the former receptors play only a minor role in the presently observed effects.
In conclusion, although actions of aripiprazole at D2 receptors are the most likely explanation for the present results, the potential role of other receptors remains to be determined. The present study is consistent with the concept that aripiprazole, by reducing the reinforcing effect of cocaine, could be used for treatment of cocaine addiction.
References
Andersen PH, Nielsen EB, Grønvald FC, Bræstrup C (1986) Some atypical neuroleptics inhibit [3H]SCH 23390 binding in vivo. Eur J Phamacol 120:143–144
Auclair AL, Kleven MS, Besnard J, Depoortère R, Newman-Tancredi A (2006) Actions of novel antipsychotic agents on apomorphine-induced PPI disruption: influence of combined serotonin 5-HT1a receptor activation and dopamine D2 receptor blockade. Neuropsychopharmacology 31:1900–1909
Bannon MJ (2005) The dopamine transporter: role in neurotoxicity and human disease. Toxicol Appl Pharmacol 204:355–360
Bardin L, Kleven MS, Barret-Grévoz C, Depoortère R, Newman-Tancredi A (2006) Antipsychotic-like vs cataleptogenic actions in mice of novel antipsychotics having D2 antagonist and 5-HT1a agonist properties. Neuropsychopharmacology 31:1869–1879
Barrett AC, Miller JR, Dohrmann JM, Caine SB (2004) Effects of dopamine indirect agonists and selective D1-like and D2-like agonists and antagonists on cocaine self-administration and food maintained responding in rats. Neuropharmacology 47:256–273
Beardsley PM, Sokoloff P, Balster RL, Schwartz JC (2001) The D3R partial agonist, BP 897, attenuates the discrimitive stimulus effects of cocaine and d-amphetamine and is not self-administered. Behav Pharmacol 12:1–11
Beresford TP, Clapp L, Martin B, Wiberg JL, Alfers J, Beresford HF (2005) Aripiprazole in schizophrenia with cocaine dependence; a pilot study. J Clin Psychopharmacol 25:363–366
Bergman J, Kamien JB, Spealman RD (1990) Antagonism of cocaine self-administration by selective dopamine D(1) and D(2) antagonists. Behav Pharmacol 1:355–363
Blokhina EA, Kashkin VA, Zvartau EE, Danysz W, Bespalov AY (2005) Effects of nicotinic and NMDA receptor channel blockers on intravenous cocaine and nicotine self-administration in mice. Eur Neuropsychopharmacol 15:219–225
Brown ES, Jeffres J, Liggin JD, Garza M, Beard L (2005) Switching outpatients with bipolar or schizoaffective disorders and substance abuse from their current antipsychotics to aripiprazole. J Clin Psychiatry 66:756–760
Burris KD, Molski TF, Xu C, Ryan E, Tottori K, Kikuchi T, Yocca FD, Molinoff PB (2002) Aripiprazole, a novel antipsychotic, is a high-affinity partial agonist at human dopamine D2 receptors. J Pharmacol Exp Ther 302:381–389
Burstein ES, Ma J, Wong S, Gao Y, Pham E, Knapp AE, Nash NR, Olsson R, Davis RE, Hacksell U, Weiner DM, Brann MR (2005) Intrinsic efficacy of antipsychotics at human D2, D3, and D4 dopamine receptors: identification of the clozapine metabolite N-desmethylclozapine as a D2/D3 partial agonist. J Pharmacol Exp Ther 315:1278–1287
Caine SB, Koob GF (1993) Modulation of cocaine self-administration in the rat through D3 dopamine receptors. Science 260:1814–1816
Caine SB, Koob GF (1995) Pretreatment with the dopamine agonist 7-OH-DPAT shifts the cocaine self-administration dose-effect function to the left under different schedules in the rat. Behav Pharmacol 6:333–347
Caine SB, Negus SS, Mello NK, Bergman J (1999) Effects of dopamine D1-like and D2-like agonists in rats that self-administer cocaine. J Pharmacol Exp Ther 291:353–360
Caine SB, Negus SS, Melle NK (2000) Effects of dopamine D1-like and D2-like agonists on cocaine self-administration in rhesus monkeys: rapid assessment of cocaine dose-effect functions. Psychopharmacology 148:41–51
Callahan PM, Cunningham KA (1993) Discriminative stimulus properties of cocaine in relation to dopamine D2 receptor function in rats. J Pharmacol Exp Ther 266:585–592
Criswell HE, Ridings A (1983) Intravenous self-administration of morphine by naïve mice. Pharmacol Biochem Behav 18:467–470
Dalley JW, Fryer TD, Brichard L, Robinson ES, Theobald DE, Laane K, Pena Y, Murphy ER, Shah Y, Probst K, Abakumova I, Aigbirhio FI, Richards HK, Hong Y, Baron JC, Everitt BJ, Robbins TW (2007) Nucleus accumbens D2/3 receptors predict trait impulsivity and cocaine reinforcement. Science 315:1267–1270
DeLeon A, Patel NC, Crismon ML (2004) Aripiprazole: a comprehensive review of its pharmacology, clinical efficacy, and tolerability. Clin Ther 26:649–666
Feltenstein MW, Altar CA, See RE (2007) Aripiprazole blocks reinstatement of cocaine seeking in an animal model of relapse. Biol Psychiatry 61:582–590
Fink-Jensen A, Fedorova I, Wörtwein G, Woldbye DPD, Rasmussen T, Thomsen M, Bolwig TG, Knitowski KM, McKinzie DL, Yamada M, Wess J, Basile A (2003) Role for M5 muscarinic acetylcholine receptors in cocaine addiction. J Neurosci Res 74:91–96
Grech DM, Spealman RD, Bergman J (1996) Self-administration of D1 receptor agonists by squirrel monkeys. Psychopharmacology 125:97–104
Gyertyán I, Kiss B, Gál K, Laszlovszky I, Horváth A, Hémesi LI, Sághy K, Pásztor G, Zájer M, Kapás M, Csongor ÉÁ, Domány G, Tihanyu K, Szombathelyi Z (2007) Effects of RGH-237 [N-{4-[4-(3-Aminocarbonyl-phenyl)-piperazin-1-yl]-butyl}-4-bromo-benzamide], an orally active selective dopamine D3 receptor partial agonist in animal models of cocaine abuse. J Pharm Exp Ther 320:1268–1278
Kapur S, Vanderspek SC, Brownlee BA, Nobrega JN (2003) Antipsychotic dosing in preclinical models is often unrepresentative of the clinical condition: a suggested solution based on in vivo occupancy. J Pharmacol Exp Ther 305:625–631
Khroyan TV, Barret-Laramore RL, Rowlett JK, Spealman RD (2000) Dopamine D1- and D2-like receptor mechanisms in relapse to cocaine-seeking behavior: effects of selective antagonists and agonists. J Pharmacol Exp Ther 294:680–687
Kikuchi T, Tottori K, Uwahodo Y, Hirose T, Miwa T, Oshiro Y, Morita S (1995) 7-(4-[4-(2,3-Dichlorophenyl)-1-piperazinyl]butyloxy)-3,4-dihydro-2(1H)-quinolinone (OPC-14597), a new putative antipsychotic drug with both presynaptic dopamine autoreceptor agonistic activity and postsynaptic D2 receptor antagonistic activity. J Pharmacol Exp Ther 274:329–336
Kleven MS, Barret-Grévoz C, Slot LB, Newman-Tancredi A (2005) Novel antipsychotic agents with 5-HT1a agonist properties: Role of 5-HT1a receptor activation in attenuation of catalepsy induction in rats. Neuropharmacology 49:135–143
Kuzmin A, Zvartau E, Gessa GL, Martellotta MC, Fratta W (1992) Calcium antagonists isradipine and nimodipine suppress cocaine and morphine intravenous self-administration in drug-naïve mice. Pharmacol Biochem Behav 41:497–500
Lawler CP, Prioleau C, Lewis MM, Mak C, Jiang D, Schetz JA, Gonzalez AM, Sibley DR, Mailman RB (1999) Interactions of the novel antipsychotic aripiprazole (OPC-14597) with dopamine and serotonin receptor subtypes. Neuropsychopharmacology 20:612–627
Le Foll B, Goldberg SR, Sokoloff P (2005) The dopamine D3 receptor and drug dependence: effects on reward or beyond? Neuropharmacology 49:525–541
Lile JA, Stoops WW, Vansickel AR, Glaser PEA, Hays LR, Rush CR (2005) Aripiprazole attenuates the discriminative-stimulus and subject-rated effects of d-amphetamine in humans. Neuropsychopharmacology 30:2103–2114
Marona-Lewicka D, Nichols DE (2004) Aripiprazole (OPC-14597) fully substitutes for the 5-HT1a receptor agonist LY293284 in the drug discrimination assay in rats. Psychopharmacology 172:415–421
Mello NL, Negus SS (1996) Preclinical evaluation of pharmacotherapies for treatment of cocaine and opioid abuse using drug self-administration procedures. Neuropsychopharmacology 14:375–424
Moore RJ, Vinsant SL, Nader MA, Porrino LJ, Friedman DP (1998) Effect of cocaine self-administration on dopamine D2 receptors in rhesus monkeys. Synapse 30:88–96
Morgan D, Grant KA, Gage HD, Mach RH, Kaplan JR, Prioleau O, Nader SH, Buchheimer N, Ehrenkaufer RL, Nader MA (2002) Social dominance in monkeys: dopamine D2 receptors and cocaine self-administration. Nat Neurosci 5:169–174
Müller CP, Huston JP (2006) Determining the region-specific contributions of the 5-HT receptors to the psychostimulant effects of cocaine. Trends Pharmacol Sci 27:105–112
Müller CP, Carey RJ, Huston JP, Silva MADS (2007) Serotonin and psychostimulant addiction: focus on 5-HT1a-receptors. Prog Neurobiol 81:133–178
Nader MA, Mach RH (1996) Self-administration of the dopamine D3 agonist 7-OH-DPAT in rhesus monkeys is modified by prior cocaine exposure. Psychopharmacology 125:13–22
Nader MA, Daunais JB, Moore T, Nader SH, Moore RJ, Smith HR, Friedman DP, Porrino LJ (2002) Effects of cocaine self-administration on striatal dopamine systems in rhesus monkeys: initial and chronic exposure. Neuropsychopharmacology 27:35–46
Nader MA, Morgan D, Gage HD, Nader SH, Calhoun TL, Buchheimer N, Ehrenkaufer R, Mach RH (2006) PET imaging of dopamine D2 receptors during chronic cocaine self-administration in monkeys. Nat Neurosci 9:1050–1056
Nakai S, Hirose T, Uwahodo Y, Imaoka T, Okazaki H, Miwa T, Nakai M, Yamada S, Dunn B, Burris KD, Molinoff PB, Tottori K, Altar CA, Kikuchi T (2003) Diminished catalepsy and dopamine metabolism distinguish aripiprazole from haloperidol or risperidone. Eur J Pharmacol 472:89–97
Natesan S, Reckless GE, Nobrega JN, Fletcher PJ, Kapur S (2006) Dissociation between in vivo occupancy and functional antagonism of dopamine D2 receptors: comparing aripiprazole to other antipsychotics in animal models. Neuropsychopharmacology 31:1854–1863
Nielsen EB, Hansen JB, Grønvald FC, Swedberg MDB, Scheideler M (1997) NNC 19-1228 and NNC 22-0031, novel neuroleptics with a “mesolimbic-selective” behavioral profile. Psychopharmacology 129:168–178
Paterson NE, Semenova S, Gasparini F, Markou A (2003) The mGluR5 antagonist MPEP decreased nicotine self-administration in rats and mice. Psychopharmacology 167:257–264
Pilla M, Perachon S, Sautel F, Garrido F, Mann A, Wermuth CG, Schwartz JC, Everitt BJ, Sokoloff P (1999) Selective inhibition of cocaine-seeking behaviour by a partial dopamine D3 receptor agonist. Nature 400:371–375
Platt DM, Rodefer JS, Rowlett JK, Spealman RD (2003) Suppression of cocaine- and food-maintained behavior by the D2-like receptor partial agonist terguride in squirrel monkeys. Psychopharmacology 166:298–305
Pulvirenti L, Koob GF (2002) Being partial to psychostimulant addiction therapy. Trends Pharmacol Sci 23:151–153
Pulvirenti L, Balducci C, Piercy M, Koob GF (1998) Characterization of the effects of the partial dopamine agonist terguride on cocaine self-administration in the rat. J Pharmacol Exp Ther 286:1231–1238
R Development Core Team (2006) R: a language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria http://www.R-project.org
Ranaldi R, Wang Z, Woolverton WL (2001) Reinforcing effects of D2 dopamine receptor agonists and partial agonists in rhesus monkeys. Drug Alcohol Depend 64:209–217
Rasmussen T, Swedberg MDB (1998) Reinforcing effects of nicotinic compounds: intravenous self-administration in drug-naïve mice. Pharmacol Biochem Behav 60:567–573
Rasmussen T, Sauerberg P, Nielsen EB, Swedberg MDB, Thomsen C, Sheardown MJ, Jeppesen L, Calligaro DO, DeLapp NW, Whitesitt C, Ward JS, Shannon HE, Bymaster FP, Fink-Jensen A (2000) Muscarinic receptor agonists decrease cocaine self-administration rates in drug-naïve mice. Eur J Pharmacol 402:241–246
Riddle EL, Fleckenstein AE, Hanson GL (2005) Role of monoamine transporters in mediating psychostimulant effects. APPS Journal 7:E847–E851
Ritz MC, Lamb RJ, Goldberg SR, Kuhar MJ (1987) Cocaine receptors on dopamine transporters are related to self-administration of cocaine. Science 237:1219–1223
Schwabe K, Koch M (2007) Effects of Aripiprazole on operant responding for a natural reward after psychostimulant withdrawal in rats. Psychopharmacology 191:759–765
Self DW, Barnhart WJ, Lehman DA, Nestler EJ (1996) Opposite modulation of cocaine seeking behavior by D1- and D2-like dopamine receptor agonists. Science 271:1586–1589
Semba J, Watanabe A, Kito S, Toru M (1995) Behavioural and neurochemical effects of OPC-14597, a novel antipsychotic drug, on dopaminergic mechanisms in rat brain. Neuropharmacology 34:785–791
Shapiro DA, Renock S, Arrington E, Chiodo LA, Liu L, Sibley DR, Roth BL, Mailman R (2003) Aripiprazole, a novel atypical antipsychotic drug with a unique and robust pharmacology. Neuropsychopharmacology 28:1400–1411
Sinnott RS, Mach RH, Nader MA (1999) Dopamine D2/D3 receptors modulate cocaine’s reinforcing and discriminative stimulus effects in rhesus monkeys. Drug Alcohol Depend 54:97–110
Smith PK, Krohn RI, Hermanson GT, Mallia AK, Gartner FH, Provenzano MD, Fujimoto EK, Goeke NM, Olson BJ, Klenk DC (1985) Measurement of protein using bicinchoninic acid. Anal Biochem 150:76–85
SPSS Inc. (2004) SPSS Base version 13.0 User Manual. SPSS Inc., Chicago
Stark AD, Jordan S, Allers KA, Bertekap RL, Chen R, Kannan TM, Molski TF, Yocca FD, Sharp T, Kikuchi T, Burris KD (2007) Interaction of the novel antipsychotic aripiprazole with 5-HT1a and 5-HT2a receptors: functional receptor-binding and in vivo electrophysiological studies. Psychopharmacology 190:373–382
Stoops WW, Lile JA, Glaser PEA, Rush CR (2006) A low dose of aripiprazole attenuates the subject-rated effects of d-amphetamine. Drug Alcohol Depend 84:206–209
Tidey JW, Bergman J (1998) Drug discrimination in methamphetamine-trained monkeys: agonist and antagonist effects of dopaminergic drugs. J Pharmacol Exp Ther 285:1163–1174
Urban JD, Vargas GA, von Zastrow M, Mailman RB (2007) Aripiprazole has functionally selective actions at dopamine D2 receptor-mediated signaling pathways. Neuropsychopharmacology 32:67–77
Volkow ND, Fowler JS, Wang GJ, Hitzemann R, Logan J, Schlyer DJ, Dewey SL, Wolf AP (1993) Decreased dopamine D2 receptor availability is associated with reduced frontal metabolism in cocaine abusers. Synapse 14:169–177
Volkow ND, Wang GJ, Fowler JS, Logan J, Gatley SJ, Gifford A, Hitzemann R, Ding YS, Pappas N (1999) Prediction of reinforcing responses to psychostimulants in humans by brain dopamine D2 receptor levels. Am J Psychiatry 156:1440–1443
Volkow ND, Wang GJ, Fowler JS, Thanos PP, Logan J, Gatley SJ, Gifford A, Ding YS, Wong C, Pappas N (2002) Brain DA D2 receptors predict reinforcing effects of stimulants in humans: replication study. Synapse 46:79–82
Wee S, Mandyam CD, Lekic DM, Koob GF (2007a) a1-Noradrenergic system role in increased motivation for cocaine intake in rats with prolonged access. Eur Neuropsychopharm (in press)
Wee S, Wang Z, Woolverton WL, Pulvirenti L, Koob GF (2007b) Effect of aripiprazole, a partial dopamine D2 receptor agonist, on increased rate of methamphetamine self-administration in rats with prolonged session duration. Neuropsychopharmacology 32:2238–2247
Weissenborn R, Deroche V, Koob GF, Weiss F (1996) Effects of dopamine agonists and antagonists on cocaine-induced operant responding for a cocaine-associated stimulus. Psychopharmacology 16:311–322
Wise RA, Murray A, Bozarth MA (1990) Bromocriptine self-administration and bromocriptine-reinstatement of cocaine-trained and heroin-trained lever pressing in rats. Psychopharmacology 100:355–360
Woolverton WL, Goldberg LI, Ginos J (1984) Intravenous self-administration of dopamine receptor agonists by rhesus monkeys. J Pharmacol Exp Ther 230:678–683
Acknowledgements
The authors wish to thank Dr. Rene Holm for the expert assistance on formulation of aripiprazole. In addition, excellent technical assistance by Mrs. Lisbet Petri, Mrs. Helle Porsdal, Mr. Leslie Griffiths, and Mrs. Rikke Madsen is highly appreciated.
The authors certify that there is no actual or potential conflict of interest in relation to this article.
Author information
Authors and Affiliations
Corresponding author
Additional information
This work was supported by grants from the Augustinus Foundation, Ivan Nielsen Foundation, Lundbeck Foundation, Psychiatric Basic Research Foundation and Theodore and Vada Stanley Foundation.
Rights and permissions
About this article
Cite this article
Sørensen, G., Sager, T.N., Petersen, J.H. et al. Aripiprazole blocks acute self-administration of cocaine and is not self-administered in mice. Psychopharmacology 199, 37–46 (2008). https://doi.org/10.1007/s00213-008-1069-z
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00213-008-1069-z