
Abstract
Rationale
Aripiprazole is an atypical antipsychotic drug with high in vitro affinity for 5-HT1A, 5-HT2A and dopamine (DA) D2 receptors. However, its in vivo actions in the brain are still poorly characterized.
Objective
The aim was to study the in vivo actions of aripiprazole in the rat and mouse brain.
Methods
Brain microdialysis and single-unit extracellular recordings were performed.
Results
The systemic administration of aripiprazole reduced 5-HT output in the medial prefrontal cortex (mPFC) and dorsal raphe nucleus of the rat. Aripiprazole also reduced extracellular 5-HT in the mPFC of wild-type (WT) but not of 5-HT1A (−/−) knockout (KO) mice. Aripiprazole reversed the elevation in extracellular 5-HT output produced by the local application of the 5-HT2A/2C receptor agonist DOI in mPFC. Aripiprazole also increased the DA output in mPFC of WT but not of 5-HT1A KO mice, as observed for atypical antipsychotic drugs, in contrast to haloperidol. Contrary to haloperidol, which increases the firing rate of DA neurons in the ventral tegmental area (VTA), aripiprazole induced a very moderate reduction in dopaminergic activity. Haloperidol fully reversed the inhibition in dopaminergic firing rate induced by apomorphine, whereas aripiprazole evoked a partial reversal that was significantly different from that evoked by haloperidol and from the spontaneous reversal of dopaminergic activity in rats treated with apomorphine.
Conclusions
These results indicate that aripiprazole modulates the in vivo 5-HT and DA release in mPFC through the activation of 5-HT1A receptors. Moreover, aripiprazole behaves as a partial agonist at DA D2 autoreceptors in vivo, an action which clearly distinguishes it from haloperidol.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Schizophrenia afflicts 0.5–1% of the world population (Lewis and Lieberman 2000; Goldner et al. 2002), and its pharmacological treatment is far from optimal. First-generation (neuroleptic) antipsychotic drugs alleviate psychotic symptoms and produce severe motor side effects through the same pharmacological mechanism, blockade of dopamine (DA) D2 receptors (Kapur et al. 2000). Second-generation (atypical) antipsychotic drugs have an improved tolerability and milder motor side effects than classical neuroleptics but produce weight gain and metabolic disturbances (Newcomer 2005). Moreover, cognitive deficits, which are central to the illness (Elvevag and Goldberg 2000), are poorly treated, and there is an urgent need for improved treatments.
Aripiprazole (ARI) is a novel atypical antipsychotic drug that differs from other classical and atypical antipsychotics, improving both positive and negative symptoms of psychosis without producing extrapyramidal side effects or increases in serum prolactin (Tamminga 2002; DeLeon et al. 2004). ARI shows high affinity for a large number of monoaminergic receptors, including DA D2, 5-HT1A and 5-HT2A receptors (Shapiro et al. 2003; Green 2004).
Concerning the dopaminergic system, ARI attenuates apomorphine (APO)-induced stereotypy in mice and reduces dopa synthesis in the forebrain of reserpine-treated mice, suggesting that, unlike classical and other atypical drugs, it exhibits agonistic activity at presynaptic DA autoreceptors and antagonistic activity at postsynaptic DA D2 receptors (Kikuchi et al. 1995; Momiyama et al. 1996). In vitro, ARI activates D2 receptor-mediated inhibition of cyclic adenosine monophosphate accumulation stimulated by forskolin in Chinese hamster ovary cells transfected with the human D2L receptor gene (Burris et al. 2002; Tadori et al. 2005). On the other hand, ARI blocked the actions of DA on D1 and D2 receptors in nucleus accumbens (Amano et al. 1995). Interestingly, ARI occupies more than 80% of striatal DA D2 receptors at therapeutic doses but does not produce extrapyramidal side effects (Yokoi et al. 2002; Grunder et al. 2003), thus deviating from the classical relationship between DA D2 receptor occupancy and occurrence of motor side effects. Overall, these data suggest that ARI may act as a partial DA D2 agonist in vivo.
In parallel to its actions at DA D2 receptors, ARI has affinity for several 5-HT receptors (Lawler et al. 1999; Shapiro et al. 2003). Hence, it behaves as a partial agonist at 5-HT1A receptors and as an antagonist at 5-HT2A receptors as assessed in vitro (Jordan et al. 2002; Tamminga 2002; Shapiro et al. 2003; Green 2004). These activities may also participate in the therapeutic action of ARI. In addition to the affinity for 5-HT2A receptors displayed by atypical antipsychotics (Meltzer 1999), ARI may activate 5-HT1A receptors, which are currently a focus of interest in antipsychotic drug development (Millan 2000). Hence, atypical (but not classical) antipsychotics increase DA release in the medial prefrontal cortex (mPFC; Rollema et al. 1997, 2000; Ichikawa et al. 2001; Assié et al. 2005), an effect that depends on the activation of 5-HT1A receptors localized in mPFC (Díaz-Mataix et al. 2005). Given the crucial role of prefrontal DA in cognitive function (Williams and Goldman-Rakic 1995), the increase in mPFC DA release may underlie the slightly superior effects of clozapine and other atypical antipsychotic drugs on the spectrum of cognitive and negative/affective symptoms (Harvey and Keefe 2001) by normalizing a putative impaired dopaminergic transmission in mPFC (Weinberger et al. 1994; Akil et al. 1999).
In the present study, we examined the in vivo actions of ARI on the serotonergic and dopaminergic systems in rodent brain, with a special emphasis on the possible actions on 5-HT1A receptors and on its putative partial agonist activity on DA D2 receptors.
Materials and methods
Animals and treatments
Male albino Wistar rats weighing 250–320 g and C57BL/6 mice, 10–12 week old at the time of the experiments, were used (Iffa Credo, Lyon, France). 5-HT1A receptor knockout KO(−/−) mice (referred onwards as KO) were generated at Princeton University (Parks et al. 1998) and had the same genetic background as their wild-type (WT) counterparts (C57BL/6). From this initial source, a stable colony was grown in the animal facility of the University of Barcelona School of Medicine. Animals were kept in a controlled environment (12-h light–dark cycle and 22 ± 2°C room temperature) with food and water provided ad libitum. Animal care followed the European Union regulations (O.J. of E.C. L358/1 18/12/1986) and was approved by the Institutional Animal Care and Use Committee.
Stereotaxic coordinates (in mm) were taken from bregma and dura mater according to the atlas of Paxinos and Watson (1998) for rat and Franklin and Paxinos (1997) for mouse.
8-OH-DPAT, APO, DOI (1-[2,5-dimethoxy-4-iodophenyl-2-aminopropane]) and WAY-100635 were from RBI (Natick, MA). BAYx3702 was from BAYER, and ARI was from Bristol Myers Squibb. Concentrated solutions were prepared in artificial CSF (aCSF) or saline and stored at −80°C (pH adjusted to 6.5–7.4 with NaHCO3 when necessary), and working solutions were prepared daily by dilution in aCSF. APO was freshly prepared daily in saline. Drugs were dissolved in vehicle at the appropriate concentrations and injected i.p. or i.v. (femoral vein) as indicated in “Results”. In microdialysis experiments involving local drug administration, the stated drug concentrations were applied by reverse dialysis (uncorrected for drug recovery). For systemic administrations, ARI was dissolved in a vehicle consisting of hydroxypropyl-β-cyclodextrin (1.4 g dissolved in 15 ml distilled water). Haloperidol (HAL) was used as commercial injectable solution (Laboratorios Esteve, Barcelona, Spain) and was injected i.v. in electrophysiological experiments. Control rats received the vehicle (systemic drug administration) or were perfused with aCSF for the whole period (local microdialysis experiments). The bars in the figures show the period of drug application (corrected for the void volume of the system).
After experimental procedures were completed, animals were killed by an overdose of anesthetic, and a careful histological verification of the correct placement of the implants was carried out.
Single-unit recordings
We examined the responses of ventral tegmental area (VTA) DA neurons to the systemic administration of APO. The suppressant action of APO was then counteracted with HAL and ARI. Rats were anesthetized (chloral hydrate 400 mg/kg i.p.) and positioned in a David Kopf stereotaxic frame. Thereafter, chloral hydrate was continuously administered i.p. at a dose of 50–70 mg/kg·h using a perfusion pump (Fa et al. 2003). Body temperature was maintained at 37°C with a heating pad. To minimize pulsation, the atlanto-occipital membrane was punctured to release some CSF. DA neurons were recorded extracellularly with glass micropipettes pulled from 2.0-mm capillary glass (WPI, Sarasota, FL) on a Narishige PE-2 pipette puller (Narishige Sci. Inst., Tokyo, Japan). Microelectrodes were filled with 2 M NaCl. Typically, in vitro impedance was 4–10 MΩ. Single-unit extracellular recordings were amplified with a Neurodata IR283 (Cygnus Technology, Delaware Water Gap, PA), postamplified and filtered with a Cibertec amplifier (Madrid, Spain) and computed on-line using a DAT 1401 plus interface system Spike2 software (Cambridge Electronic Design, Cambridge, UK).
Descents in VTA were carried out at AP −5.0 to −5.6, L −0.5 to −1 and DV −7.5 to −9.0. The identification of DA neurons and burst firing analysis was carried out according to the criteria of Grace and Bunney (1984) as previously used (Celada et al. 1999; Díaz-Mataix et al. 2005). Briefly, neurons were considered dopaminergic if they possessed the following characteristics: (1) action potential duration greater than 2.5 ms, (2) typical bi- or triphasic waveform, often with a notch in the initial rising phase, (3) slow firing rate (recorded neurons fired at 1–6 spikes/s in control rats), and (4) frequent presence of bursts. The structure of bursts was defined as starting with a first interspike interval of <80 ms and ending with an interspike interval of 160 ms or greater (Grace and Bunney 1984).
Microdialysis
Microdialysis procedures in rats and mice were conducted essentially as recently described in Bortolozzi et al. (2003), Bortolozzi and Artigas (2003) and Amargós-Boch et al. (2004). Rats were anesthetized with sodium pentobarbital (60 mg/kg i.p.) and implanted with 4-mm concentric dialysis probes (Cuprophan) in mPFC at AP +3.2, L −0.8 and DV −6.0. Groups of rats were also implanted with probes in the dorsal raphe nucleus (DR) at −7.4, L −3.1 and DV −7.5 with a lateral angle of 30° (probe tip 1.5 mm). Microdialysis experiments were performed in freely moving rats >20 h after surgery. Probes were perfused with aCSF pumped at 1.5 μl/min. After an initial 100-min stabilization period, four baseline samples were collected (20 min each) before local (reverse dialysis) or systemic drug administration and then successive dialysate samples were collected.
For mice, the manufacture of the probes was adapted from that previously described for rats (Bortolozzi et al. 2003). Surgical and microdialysis procedures were identical to those described for rats except for the dose of anesthesia (sodium pentobarbital, 40 mg/kg, i.p.), the length of dialysis membrane (2 mm) and the brain coordinates (in mm) of the mPFC: AP +2.2, L −0.2 and DV −3.4.
The concentration of 5-HT and DA in dialysate samples was determined by high-performance liquid chromatography (HPLC) with amperometric detection, using slight modifications of previously described methods. Updated procedures can be found in Bortolozzi et al. (2003) and Díaz-Mataix et al. (2005). Brain dialysates were collected in microvials and were injected into the HPLC. For DA analysis, microvials contained 5 μl of 10 mM perchloric acid. The amperometric detection (Hewlett Packard 1049 detector) of 5-HT ad DA was carried out, respectively, at +0.6 and +0.75 V. Detection limits were typically 1–1.5 fmol for 5-HT and 3 fmol for DA.
Data analysis
Changes in the electrical activity of DA neurons were assessed using repeated measures analysis of variance (ANOVA) followed by post hoc t tests. Paired t test was also used when appropriate. Firing rate and burst firing were quantified by averaging the values each minute after i.v. drug administration (omitting the first minute). Microdialysis results are expressed as fmol/fraction (uncorrected for recovery) and shown in figures as percentages of basal values (individual means of four pre-drug fractions). Microdialysis data were analyzed using one- or two-way repeated measures ANOVA, with drug as independent factor and time as repeated factor. The area under the curve values of selected periods of drug treatment were also calculated and compared using ANOVA. Data are expressed as the mean ± SEM.
Results
Effects of ARI on spontaneous 5-HT release in rat and mice brain
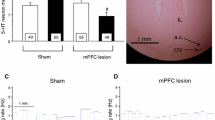
Baseline dialysate 5-HT values in rat brain were 75 ± 16 fmol/fraction (DR, n = 8) and 37 ± 2 fmol/fraction (mPFC, n = 26). In mouse mPFC, 5-HT values were 29 ± 2 fmol/fraction (WT, n = 17) and 29 ± 3 fmol/fraction (5-HT1A KO, n = 18). The administration of ARI (3, 10 and 30 mg/kg i.p.) reduced the 5-HT output in the mPFC and the dorsal raphe (DR) of freely moving rats compared with rats treated with the vehicle (hydroxypropyl-β-cyclodextrin solution). Successive vehicle injections elicited moderate (~20%) increases of extracellular 5-HT, likely due to the injection-associated stress (Adell et al. 1997), whereas ARI induced a significant reduction in 5-HT in DR (p < 0.005 group effect, p < 0.01 time effect and p < 0.005 time × group interaction) and mPFC (p < 0.05 group effect and p < 0.02 time effect; Fig. 1). The maximal reduction in 5-HT release was attained at 3 mg/kg, and subsequent injections of 10 and 30 mg/kg ARI did not reduce dialysate 5-HT further.

Effect of the i.p. administration of vehicle or aripiprazole (ARI, 3, 10 and 30 mg/kg, cumulative doses) on the 5-HT output in the medial prefrontal cortex (mPFC) and dorsal raphe (DR) of rats. Vertical arrows show the injection times (corrected for void volume of the system). Data are means ± SEM of four rats per group. See text for statistical details
To examine the involvement of 5-HT1A receptors in the 5-HT-reducing effect of ARI, we examined its effects on the 5-HT release in mPFC of WT and 5-HT1A KO mice. The i.p. administration of the vehicle did not alter dialysate 5-HT except for a short time increase after the injection, which is likely due to the handling and injection stress and that was slightly greater in 5-HT1A KO mice as previously observed (Bortolozzi et al. 2004; Fig. 2a). The i.p. administration of 3 mg/kg ARI induced a moderate but significant reduction in the 5-HT output, which was not different between WT and KO mice (n = 6 each; p < 0.0001 significant effect of time but not of group or time × group interaction; Fig. 2b). The subsequent injection of the selective 5-HT1A receptor agonist 8-OH-DPAT (0.5 mg/kg i.p.) markedly reduced the 5-HT output in a subgroup of WT but not KO mice within this experiment (p < 0.03 group effect, p = 0.06 time effect and p < 0.001 time × group interaction; n = 3 each; Fig. 2b).

Effect of the i.p. administration of vehicle (a), 3 mg/kg aripiprazole (ARI; b) and 30 mg/kg ARI (c) on the 5-HT output in the mPFC of wild type (WT) and 5-HT1A receptor knockout (KO) mice (n = 6 each except the vehicle/WT group, n = 5). The subsequent administration of the selective 5-HT1A agonist 8-OH-DPAT (0.5 mg/kg i.p.) reduced the 5-HT output in WT but not in 5-HT1A receptor KO mice (n = 3 for the groups treated with 3 mg/kg ARI; n = 6 for the groups treated with 30 mg/kg ARI). See text for statistical details
In contrast, the administration of 30 mg/kg ARI elicited a more marked reduction in the 5-HT output in WT than in 5-HT1A KO mice (p < 0.00001 time effect and p < 0.0005 time × group interaction; n = 6 each; Fig. 2c). As also observed previously, the subsequent i.p. administration of 0.5 mg/kg 8-OH-DPAT reduced the 5-HT output further in WT but not in 5-HT1A receptor KO mice (p < 0.0001 group effect, p < 0.0001 time effect and p < 0.0001 time × group interaction; n = 6 each; Fig. 2c). These results indicate that 30 but not 3 mg/kg ARI reduced 5-HT output in mouse mPFC by a 5-HT1A-dependent mechanism.
Effects of ARI on DOI-stimulated 5-HT release in rat and mice mPFC
Previous reports indicate that the local application of both classical (chlorpromazine, HAL) and atypical (clozapine, olanzapine) antipsychotic drugs reverse the increase in 5-HT levels in mPFC evoked by the hallucinogen DOI, a 5-HT2A/2C receptor agonist (Bortolozzi et al. 2003). We therefore examined the ability of ARI to reverse DOI-stimulated 5-HT release in mPFC.
As previously observed in rat and mouse mPFC (Martín-Ruiz et al. 2001a; Bortolozzi et al. 2003), the application of 100 μM DOI by reverse dialysis doubled the local 5-HT output (p < 0.0001; Fig. 3). The co-perfusion of 100 μM ARI did not counteract the effect of DOI, whereas a higher concentration (300 μM) significantly antagonized DOI’s effect on 5-HT output (p = 0.057 group effect, p < 0.00001 time effect and p < 0.00001 time × group interaction; Fig. 3).

The application of the 5-HT2A/2C receptor agonist DOI (100 μM) by reverse dialysis enhanced the local 5-HT output in rat mPFC (n = 6). The co-perfusion of 300 μM (n = 7) but not 100 μM (n = 5) aripiprazole (ARI) significantly reversed the 5-HT elevation evoked by the application of DOI. Horizontal bars show the period of drug application. See text for statistical analysis
Comparison of the effect of HAL and ARI on the APO-induced suppression of DA cell firing in rat VTA
The systemic administration of ARI (0.4–0.8 mg/kg i.v.) to chloral hydrate-anesthetized rats (n = 6) induced a moderate reduction in the firing rate of VTA dopaminergic neurons from 3.2 ± 0.7 spikes/s (baseline) to 2.7 ± 0.6 spikes/s (0.4 mg/kg i.v.) and 2.7 ± 0.7 spikes/s (0.8 mg/kg i.v.; p < 0.03, one-way repeated measures ANOVA; p < 0.05 between both doses of ARI and baseline; non-significant differences between 0.4 and 0.8 mg/kg; post hoc Tukey t test). When considering the maximal effect in all neurons, irrespective of the dose required, the firing rate was reduced to 2.4 ± 0.6 spikes/s (p < 0.008, paired Student’s t test vs baseline). The percentage of spikes fired in burst was also reduced by ARI treatment, from 12.3 ± 3.6% (baseline) to 5.9 ± 2.3% (0.4 mg/kg i.v.) and 5.1 ± 3.1% (0.8 mg/kg i.v.; p = 0.057, one-way repeated measures ANOVA). The maximal effect was to 4.4 ± 2.6% (p < 0.035 vs baseline). Figure 4 shows representative examples of the effect of ARI administration on two dopaminergic neurons.

Integrated firing rate histograms showing the effects of i.v. injections (arrows) of aripiprazole (ARI; a, b) and haloperidol (HAL; c) on the firing activity of dopaminergic neurons in the VTA (ordinate: spikes/10 s; see bar time in abscissa). The upper traces in these panels show representative burst trains corresponding to 2-min recordings obtained in baseline conditions and after the administration of 0.8 mg/kg i.v. ARI or 0.2 mg/kg i.v. HAL. The neuron in a had a basal firing rate of 4.7 spikes/s and decreased to 4.1 and 3.8 spikes/s after 0.4 and 0.8 mg/kg i.v. ARI. Burst firing was 20% and was reduced to 14% at both doses of ARI. The neuron in b had a basal firing rate of 2.27 spikes/s and decreased to 2.1 and 1.58 spikes/s after 0.4 and 0.8 mg/kg i.v. ARI. Burst firing was 13%, 11% and 5% in baseline conditions and after the i.v. injection of 0.4 and 0.8 mg/kg ARI, respectively. Panel c shows a representative example of the effect of i.v. HAL on the firing rate of VTA dopaminergic neurons. Contrary to ARI, HAL increased the firing rate of dopaminergic neurons (from 2.1 spikes/s in baseline conditions to 4.9 and 6.2 spikes/s after 0.1 and 0.2 mg/kg). Burst firing was also increased from 23% to 65%. The neuron in c corresponds to data reported in Díaz-Mataix et al. (2005)
The moderate suppressing effect of ARI on dopaminergic cell firing was consistent with a weak partial agonist action. We then examined whether ARI could reverse the inhibition of cell firing induced by APO. Rats were injected with a single dose of APO (50 μg/kg i.v.), which induced a marked suppression of the firing of DA cells in the VTA. Two minutes later, rats were administered with either saline (n = 9), HAL (200 μg/kg i.v.; n = 6) or ARI (400 or 800 μg/kg i.v; n = 15 and 4, respectively.). As expected, HAL administration completely reversed the suppressant effect of APO on the overall firing rate (Fig. 5). The i.v. administration of 400 or 800 μg/kg ARI significantly reversed the suppressant effect of APO on firing rate, but the reversal was less marked than that produced by HAL. The effects of 400 μg/kg ARI were examined in various groups of rats (up to a total of n = 15), which gave consistently the same partial recovery of DA cell firing.

Integrated firing rate histograms of VTA dopaminergic neurons showing representative examples of the inhibitory effect of apomorphine (50 μg/kg i.v., first arrow) followed by injections (second arrow) of saline (a), haloperidol 200 μg/kg i.v. (HAL, b) and aripiprazole 400 μg/kg i.v. (ARI, c). Note the suppression of cell firing induced by apomorphine in all units. As expected, HAL produced a full reversal of the inhibition followed by a rebound increase that is likely due to the full blockade of self-inhibitory mechanisms mediated by DA D2 receptors at cell body level. In contrast, ARI induced a partial, but significant, reversal of the apomorphine-induced inhibition
Figure 5 shows representative examples of the suppressant effect of APO on DA cell firing followed by spontaneous reversal (Fig. 5a) and by the effects of HAL (Fig. 5b) and ARI (Fig. 5c). Figure 6 shows the average effects in all units examined. Analysis of the data with two-way ANOVA showed a significant effect of the group (p < 0.01), time (p < 0.00001) and time × group interaction (p < 0.00001). Post hoc analysis revealed a significant difference between HAL and saline, ARI and saline and HAL and ARI, with no significant differences between the two ARI doses. Likewise, burst firing was significantly different among rats treated with APO alone and with APO + HAL and APO + ARI (0.8 ± 0.6%, 18.6 ± 11.7% and 3.0 ± 2.1%, respectively; p < 0.05 one-way ANOVA; burst firing was calculated from the last 3 min of recording; see Fig. 5).

Average effects of the administration of apomorphine followed by saline (open circles, n = 9), haloperidol 200 μg/kg i.v. (HAL, open squares, n = 6), aripiprazole 400 μg/kg i.v. (ARI, filled circles, n = 15) and ARI 800 μg/kg i.v. (filled squares, n = 4). *p < 0.05 vs apomorphine + saline, **p < 0.05 vs apomorphine + HAL
Modulation of the mesocortical DA release by ARI in mouse PFC
Baseline DA values in mPFC dialysates were 7 ± 1 fmol/fraction (WT, n = 38) and 6 ± 1 fmol/fraction (5-HT1A KO, n = 35). The i.p. administration of the vehicle did not alter DA release in the mPFC of WT and 5-HT1A KO mice. A short-lasting increase was observed after the injection of the vehicle, which, as that seen for 5-HT, can be attributed to stress (Fig. 7a). The administration of the selective 5-HT1A agonist 8-OH-DPAT (0.5 mg/kg i.p.) increased the DA output in WT but not in 5-HT1A mice (p < 0.01 group effect, p < 0.0001 time effect and p < 0.02 time × group interaction; Fig. 7b). In WT mice, the maximal increase in DA output was to 181 ± 24% of baseline. DA values remained stable for approximately four dialysate fractions (80 min). Thereafter, DA values ranged between 140 and 160% of baseline. In contrast, 8-OH-DPAT did not alter DA output in 5-HT1A KO mice.

The administration of the selective 5-HT1A agonists 8-OH-DPAT (b) and BAYx3702 (c) significantly increased the DA output in the mPFC of WT but not of 5-HT1A KO mice (n = 6–8 rats/group). The injection of saline did not alter the DA output in WT or KO mice (n = 6 and 7, respectively). See text for statistical analysis
Similarly, another selective 5-HT1A agonist, BAYx3702 (0.5 mg/kg i.p.) also increased DA output in WT but not in 5-HT1A mice (p < 0.0001 group effect, p < 0.05 time effect and p < 0.00001 time × group interaction). The maximal increase in the DA output produced by BAYx3702 was comparable to that elicited by 8-OH-DPAT (Figs. 7 and 9).
The systemic administration of 3 mg/kg i.p. ARI produced a sustained increase the DA output in the mPFC of WT mice. However, the same treatment failed to elevate cortical DA in the mPFC of 5-HT1A KO mice, except for a sharp increase just after the injection of ARI, which was also observed in WT mice (p < 0.02, group effect and p < 0.0001 time effect; Fig. 8b). At the end of the experiment, mice received a subsequent challenge dose of 8-OH-DPAT, which induced a further elevation of DA output in WT but not 5-HT1A mice (data not shown).

a) The administration of the vehicle used to dissolve aripiprazole (ARI) did not alter the DA output in wild-type (WT) and 5-HT1A knockout (KO) mice (n = 5 mice/group). b The i.p. administration of ARI at 3 mg/kg increased the DA output in the mPFC of WT but not 5-HT1A mice, except for a transient increase which was probably related to the injection stress (n = 6 mice/group). The injection of 30 mg/kg ARI also induced a significant increase in DA output in WT mice but not in 5-HT1A KO mice (n = 5 mice/group) but the effect was similar to that of 3 mg/kg. See text for statistical analysis
The administration of 30 mg/kg i.p. ARI induced a DA increase in WT mice that was not statistically different to that produced by the dose of 3 mg/kg (p < 0.0001 time effect but not significant group or time× group interactions; Figs. 8c and 9). As also observed for the 3-mg/kg dose, 30 mg/kg ARI did not increase the DA output in 5-HT1A KO mice.

Bar graph showing the average effect off the various treatments shown in Figs. 7 and 8 on control (wild-type, WT) and 5-HT1A knockout (KO) mice (SAL, saline; VEH, aripiprazole vehicle; DPAT, 8-OH-DPAT 0.5 mg/kg i.p.; BAY, 0.5 mg/kg i.p. BAYx3702; ARI, aripiprazole). Individual values have been calculated by averaging the DA values in fractions 6–16 (omitting the first fraction after injection), expressed as percentage of baseline. *p < 0.05 vs. saline (SAL), ^p < 0.05 vs. vehicle (VEH)
Discussion
The present study examined the in vivo actions of ARI on the serotonergic and dopaminergic systems in rat and mouse brain using microdialysis in awake animals and single-unit recordings of VTA dopaminergic neurons in anesthetized rats. The present data support and extend previous in vitro and in vivo observations on a dual action of ARI on both neurotransmitter systems, yet with converging effects on the DA system. In particular, we show that ARI has a partial agonist effect on the activity of dopaminergic neurons that can account for previous neurochemical and behavioural actions of this agent in experimental animals. These data can also explain, at least partly, the lack of extrapyramidal side effects of ARI despite a very large DA D2 receptor occupancy in schizophrenic patients (Grunder et al. 2003).
Electrophysiology
The data obtained indicates that ARI has a consistent but weak partial agonist effect on the activity of dopaminergic neurons in the VTA. This effect was observed on both experimental models used (basal activity and APO-induced suppression of cell firing).
ARI modestly but consistently reduced the overall firing rate of dopaminergic neurons as well as the percentage of spikes fired in bursts. The ARI-induced reduction in cell firing can be attributed to its agonist action on DA D2 receptors controlling dopaminergic cell firing (Semba et al. 1995; Shapiro et al. 2003). In contrast, ARI induced a partial but very consistent partial reversal of the suppressant action of APO on DA neuron activity, both in terms of the overall firing rate and in burst firing.
The effect of ARI appeared to reach a ceiling effect, as the suppression of cell firing when administered alone was similar at the two doses examined (0.4 and 0.8 mg/kg i.v.). These observations are consistent with a partial agonist character of ARI, which displays a low intrinsic efficacy in vitro (Burris et al. 2002; Tadori et al. 2005). The effect of ARI totally differed from that of the classical D2 receptor antagonist HAL, which greatly enhanced the activity of VTA DA neurons (Chiodo and Bunney 1983; Gessa et al. 2000; Díaz-Mataix et al. 2005).
In agreement with previous studies (Chiodo 1988), the dose of APO chosen (50 μg/kg i.v.) fully suppressed the firing rate of the majority of DA neurons recorded. Consistent with its full antagonist properties at DA D2 receptors, the subsequent administration of HAL increased the firing rate above baseline levels, most likely by antagonizing an endogenous tone on DA D2 autoreceptors. However, the administration of ARI significantly increased the firing rate of DA neurons above that produced by the spontaneous reversal (control rats received the vehicle after APO), but its effect was far from that achieved with HAL. Two different doses of ARI were used, with no significant differences between them, indicating that 400 and 800 μg/kg i.v. were maximal and equally effective, as also observed for the inhibition of spontaneous cell firing. The partial recovery of DA cell firing produced by ARI is consistent with its partial DA D2 agonist character previously observed in vitro and in vivo (Amano et al. 1995; Green 2004; Cosi et al. 2005).
Hence, ARI moderately inhibits the activity of VTA DA neurons when given alone and partially antagonizes the suppression of DA cell firing induced by APO. Indeed, few previous reports have characterized the in vivo effects of ARI and other partial DA agonists on the activity of VTA DA neurons. To our knowledge, this is the first report supporting the partial agonist effect of ARI on VTA DA cells in vivo. These observations have been confirmed in a recent independent report (Dahan et al. 2006), which strengthens their value. Thus, the present observations provide a cellular basis for the effects of ARI on DA-mediated functions in experimental animals and humans and contribute to further characterize its complex pharmacological profile.
Similar to ARI, bifeprunox also displays partial agonist properties at DA D2 receptors with an intrinsic efficacy greater than that of ARI in vitro (Cosi et al. 2005) and in vivo (Dahan et al. 2006). Contrary to these two agents, preclamol [(-)3-PPP], which also displays partial agonist activity at DA D2 receptors, failed to demonstrate sustained antipsychotic activity, perhaps due to the emergence of tolerance (Tamminga 2002). It is unclear whether the different clinical activity of ARI and preclamol is due to their distinct degree of intrinsic efficacy at DA D2 receptors or to the existence of additional activities (e.g., 5-HT receptors) in the case of ARI. Our comparison of the effect of ARI with the classical DA D2 full antagonist HAL does not permit to clarify this issue. It is possible that affinity for 5-HT1A receptors may confer some additional degree of clinical antipsychotic activity to DA D2 receptor blockade, as preclinical results suggest that this pharmacological activity is key to elevate prefrontal DA transmission (Rollema et al. 1997; Ichikawa et al. 2001; Díaz-Mataix et al. 2005; see below), and the addition of the 5-HT1A receptor agonist tandospirone improved the cognitive status of schizophrenic patients treated with standard neuroleptics (Sumiyoshi et al. 2000, 2001).
Microdialysis experiments
In rats, ARI reduced the 5-HT output in a terminal area (mPFC) and in the cell body area (DR) at doses of 3–30 mg/kg i.p. This decrease was moderate and was not dose-dependent, as similar reductions were noted at 3, 10 and 30 mg/kg, in agreement with recent data in the literature (Assié et al. 2005). In mice, the dose of 3 mg/kg produced also a moderate decline in the 5-HT output in the mPFC. However, this effect does not depend on the activation of 5-HT1A receptors, as it was common to WT and 5-HT1A KO mice. The subsequent 8-OH-DPAT administration produced a further decrease in 5-HT output in WT but not KO mice, as expected, which indicates that the 5-HT decrease in WT mice at this dose was due to other pharmacological activities of ARI. It is unlikely that a partial agonist action of ARI on DA D2 receptors may contribute to the 5-HT decrease in mPFC and DR, as DA D2 receptor agonists increase 5-HT release in the DR and the activity of 5-HT neurons (Ferré et al. 1994; Martín-Ruiz et al. 2001b). 5-HT2A receptor blockade is also unlikely to be involved, given that the 5-HT2A/2C receptor agonist DOI reduced the firing of 5-HT cells and the 5-HT output in the mPFC of anesthetized rats after systemic administration (Martín-Ruiz et al. 2001a). Among the various pharmacological activities of ARI, blockade of α1-adrenoceptors may possibly participate in this effect, as the α1-adrenoceptor antagonist prazosin reduces 5-HT cell firing (VanderMaelen and Aghajanian 1983) and 5-HT release in the DR and terminal areas (Rouquier et al. 1994; Bortolozzi and Artigas 2003). Moreover, the atypical antipsychotics clozapine and olanzapine reduce 5-HT cell firing in the DR through α1-adrenocepor blockade (Sprouse et al. 1999).
At 30 mg/kg, ARI reduced 5-HT significantly more in WT than in KO mice, which suggests that this higher dose produced a direct activation of 5-HT1A receptors controlling 5-HT release. However, the degree of activation of 5-HT1A autoreceptors produced by this dose of ARI appears to be low, as the subsequent injection of 8-OH-DPAT (0.5 mg/kg i.p.) induced a more marked reduction in 5-HT release. Overall, these data suggest a partial agonist action at 5-HT1A receptors in vivo, also consistent with previous in vitro evidence (see Shapiro et al. 2003 for review; see also below for extended discussion on this point).
As previously observed for other classical and atypical antipsychotic drugs (Bortolozzi et al. 2003), the co-perfusion of ARI in mPFC reversed the increase in 5-HT release produced by local application of the 5-HT2A/2C agonist DOI. This effect may involve various activities of ARI, such as 5-HT2A receptor antagonism, 5-HT1A receptor agonism and/or α1-adrenoceptor antagonism, as antagonists of both excitatory receptors (e.g., M100907 for 5-HT2A receptors, prazosin for α1-adrenoceptors) or agonists of inhibitory receptors (e.g., 8-OH-DPAT or BAYx3702 for 5-HT1A receptors) reverse the increase in 5-HT release produced by DOI application in mPFC (Martín-Ruiz et al. 2001a; Bortolozzi et al. 2003; Amargós-Bosch et al. 2004). Indeed, this functional antagonism is not due to a lack of specificity of the agents used but to the fact that these receptors are largely co-expressed in pyramidal neurons of the mPFC (Amargós-Bosch et al. 2004; Santana et al., unpublished observations) and play opposite roles on pyramidal cell activity (Araneda and Andrade 1991; Amargós-Bosch et al. 2004; Puig et al. 2005). Then, blockade of 5-HT2A or α1-adrenergic receptors or agonist actions at 5-HT1A receptors oppose to the action of DOI, resulting in changes in 5-HT release mediated by the mPFC-raphe circuit (Celada et al. 2001).
Recent observations indicate that atypical antipsychotics (clozapine, olanzapine and ziprasidone), but not HAL, increased the DA output in the mPFC of WT but not 5-HT1A KO mice (Díaz-Mataix et al. 2005). In the present study, we extend these observations to the case of ARI. The administration of 3 and 30 mg/kg i.p. ARI enhanced DA release in the mPFC of WT but not KO mice (except for a short-lasting increase occurring in both genotypes possibly due to injection and handling stress). The effect size was comparable to that produced by doses of the selective 5-HT1A agonists 8-OH-DPAT and BAYx3702 that are maximal or nearly maximal to suppress 5-HT release in mouse brain (0.5 mg/kg i.p.; typically ~tenfold higher than in rats). Previous studies have reported discordant results on the effect of ARI on DA output in rodent mPFC. In the rat, a bell-shaped dose–response (maximum 50% increase at 0.3 mg/kg, no change at 0.1, 1 and 3 mg/kg) was reported by Li et al. (2004), whereas no effect at all was found between 2 and 40 mg/kg by Jordan et al. (2004) and Assié et al. (2005). On the other hand, Semba et al. (1995) reported a moderate 20–25% reduction after 10 and 40 mg/kg ARI. In mouse mPFC, a significant 80% increase was found at 0.3 mg/kg (no change at 0.1, 1 and 3 mg/kg; Zocchi et al. 2005). However, this effect occurred 3 h after ARI administration, whereas Li et al. (2004) reported a short-term increase soon (80–90 min) after ARI administration. The reasons for such discrepancies are unclear and can perhaps be attributed to the insolubility of ARI in water and the different ways used to suspend or dissolve it in an aqueous vehicle before systemic administration. In this paper, we used hydroxypropyl-β-cyclodextrin, similar to the cyclodextrin used to dissolve ARI in the intramuscular commercial preparation (Abilify®). This enabled us to completely dissolve ARI at the doses used in the present study. Alternatively, given its partial agonist action on various receptors, it may be that the effect of ARI on cortical DA output depends on the behavioural state of the animals used. Likewise, as the main metabolite of ARI in rodents (DM 451) is a high affinity antagonist at DA D2 receptors (Lawler et al. 1999), it may also be that the effects on DA release can depend on the dose and/or route of administration of ARI.
5-HT1A receptor agonists and atypical antipsychotics stimulate the DA output in rat mPFC and the electrical activity of VTA DA neurons (Arborelius et al. 1993a,b; Rollema et al. 1997; Lejeune and Millan 1998; Gobert et al. 1999; Gessa et al. 2000; Ichikawa et al. 2001; Díaz-Mataix et al. 2005). This effect appears to depend in part on the activation of 5-HT1A receptors in mPFC, which may then result in an increased activity of pyramidal neurons projecting to the VTA (Díaz-Mataix et al. 2005, 2006) and the subsequent increase in the activity of mesocortical DA neurons and DA release. Interestingly, the latter effect is shared by (a) 5-HT1A receptor agonists; (b) atypical antipsychotics such as ziprasidone (Rollema et al. 2000) or ARI (Li et al. 2004; Assié et al. 2005; this study), with moderate–high in vitro affinity for 5-HT1A receptors; and (c) atypical antipsychotics with very low (clozapine) or negligible in vitro affinity for 5-HT1A receptors, such as clozapine, olanzapine or risperidone (Rollema et al. 1997; Ichikawa et al. 2001; Díaz-Mataix et al. 2005). The increase in DA output produced by atypical antipsychotics, including those without direct in vitro affinity for 5-HT1A receptors, is partly or totally antagonized by WAY-100635 (selective 5-HT1A receptor antagonist; Rollema et al. 1997, 2000; Ichikawa et al. 2001; see, however, Assié et al. 2005) and by the genetic deletion of 5-HT1A receptors (Díaz-Mataix et al. 2005).
The cellular basis for this “indirect” activation of 5-HT1A receptors by some atypical antipsychotics is presently unknown. We recently suggested the involvement of 5-HT1A receptors in cortical GABAergic interneurons (Santana et al. 2004) based on the ability of the GABAA receptor antagonist bicuculline to prevent the DA-enhancing effect of BAYx3702 (selective 5-HT1A agonist) and the atypical antipsychotics clozapine and olanzapine in rat mPFC (Díaz-Mataix et al. 2005).
ARI increased the DA output in mPFC by a 5-HT1A receptor-dependent mechanism at 3 and 30 mg/kg, with no differences between doses. However, 3 mg/kg ARI failed to reduce the 5-HT output through the activation of 5-HT1A receptors, suggesting that this dose preferentially activates a subset of 5-HT1A receptors (likely postsynaptic) involved in the modulation of DA output, different from those controlling 5-HT release in mPFC and DR (mostly, although not exclusively, 5-HT1A autoreceptors; Celada et al. 2001). ARI was active in a 5-HT1A receptor-dependent experimental paradigm (reversal of catalepsy) at 10 and 100 mg/kg (Bardin et al. 2006), which is in agreement with the doses found in this paper to elevate cortical DA release through a 5-HT1A receptor-dependent mechanism. Interestingly, a highly selective 5-HT1A receptor agonist (F15599) with a 100-fold selectivity over other monoaminergic receptors (Newman-Tancredi et al. 2006) shows the same regional/neurotransmitter profile. Hence, F15599 increased the DA output in mPFC with an ED50 of 0.03 mg/kg i.p. and reduced hippocampal 5-HT release with an ED50 of 0.24 mg/kg (eightfold difference; Assié et al. 2006).
Indeed, differences in the sensitivity of pre- and postsynaptic 5-HT1A receptors to selective agonists are known since the early report by Sprouse and Aghajanian (1988). 5-HT1A receptor agonists tend to exhibit a higher efficacy at raphe receptors controlling 5-HT cell firing and release than at hippocampal postsynaptic 5-HT1A receptors (not involved in the control of 5-HT cell firing or release), possibly due to the receptor reserve in the raphe nuclei (Cox et al. 1993). However, ARI and F15599 appear to display an opposite profile, i.e. they appear to be more efficacious at 5-HT1A receptors enhancing DA release, possibly located in mPFC (Díaz-Mataix et al. 2005) than at presynaptic 5-HT1A autoreceptors. Differences in these drugs’ abilities to stimulate the distinct signalling pathways coupled to 5-HT1A receptors may be involved. Hence, 5-HT1A receptors have been reported to be coupled to several transduction mechanisms, such as G protein-coupled inward rectifying K+ channels (Andrade et al. 1996), adenylate cyclase (both negatively and positively; Devivo and Maayani 1985; Markstein et al. 1986), microtubule-associated protein (MAP) kinase (Millan et al. 2001) or extracellular signal-regulated kinase (ERK; Bruins-Slot et al. 2006). Further work is clearly needed to elucidate whether 5-HT1A receptors may exhibit agonist-directed trafficking as it occurs with other 5-HT receptors (e.g., 5-HT2A and 5-HT2C receptors; Berg et al. 1998), as was suggested (Hoyer and Boddeke 1993; Schoeffter et al. 1997).
In summary, the present results show that ARI acts in vivo as a weak partial agonist at DA D2 receptors and is able to partially recover the inhibition in DA cell firing produced by APO. In the presence of an elevated dopaminergic tone in ventral striatum as suspected in schizophrenia (Laruelle et al. 1996; Breier et al. 1997), ARI might contribute to normalize dopaminergic transmission in this area by replacing endogenous DA at a high occupancy level (~80%) and dampening intracellular signals due to its low intrinsic efficacy. On the other hand, in common with other atypical antipsychotic drugs such as clozapine, olanzapine and ziprasidone, ARI increases DA release in mPFC by a 5-HT1A receptor-dependent mechanism, an effect that may contribute to alleviate the cognitive deficits in schizophrenia.
Abbreviations
- 5-HT:
-
5-hydroxytryptamine or serotonin
- APO:
-
apomorphine
- ARI:
-
aripiprazole
- BAY:
-
BAYx3702
- DA:
-
dopamine
- DPAT:
-
8-OH-DPAT
- DR:
-
dorsal raphe nucleus
- HAL:
-
haloperidol
- KO:
-
knockout
- mPFC:
-
medial prefrontal cortex
- VTA:
-
ventral tegmental area
- WT:
-
wild type
References
Adell A, Casanovas JM, Artigas F (1997) Comparative study in the rat of the actions of different types of stress on the release of 5-HT in raphe nuclei and forebrain areas. Neuropharmacology 36:745–751
Akil M, Pierri JN, Whitehead RE, Edgar CL, Mohila C, Sampson AR, Lewis DA (1999) Lamina-specific alterations in the dopamine innervation of the prefrontal cortex in schizophrenic subjects. Am J Psychiatry 156:1580–1589
Amano T, Matsubayashi H, Momiyama T, Ishihara K, Todo N, Sasa M (1995) Antagonizing effects of a novel antipsychotic quinolinone derivative (OPC-14597) on dopaminergic inhibition of neuronal activities in the nucleus accumbens. Prog Neuropsychopharmacol Biol Psychiatry 19:105–116
Amargós-Bosch M, Bortolozzi A, Puig MV, Serrats J, Adell A, Celada P, Toth M, Mengod G, Artigas F (2004) Co-expression and in vivo interaction of serotonin1A and serotonin2A receptors in prefrontal cortex. Cereb Cortex 14:281–299
Andrade R, Malenka RC, Nicoll RA (1996) A G protein couples serotonin and GABAB receptors to the same channels in hippocampus. Science 1986(234):1261–1265
Araneda R, Andrade R (1991) 5-Hydroxytryptamine2 and 5-hydroxytryptamine1A receptors mediate opposing responses on membrane excitability in rat association cortex. Neuroscience 40:399–412
Arborelius L, Chergui K, Murase S, Nomikos GG, Hook BB, Chouvet G, Hacksell U, Svensson TH (1993a) The 5-HT(1A) receptor selective ligands, (R)-8-OH-DPAT and (S)-UH-301, differentially affect the activity of midbrain dopamine neurons. Naunyn-Schmiedeberg’s Arch Pharmacol 347:353–362
Arborelius L, Nomikos GG, Hacksell U, Svensson TH (1993b) (R)-8-OH-DPAT preferentially increases dopamine release in rat medial prefrontal cortex. Acta Physiol Scand 148:465–466
Assié MB, Ravailhe V, Faucillon V, Newman-Tancredi A (2005) Contrasting contribution of 5-hydroxytryptamine-1A receptor activation to neurochemical profile of novel antipsychotics: frontocortical dopamine and hippocampal serotonin release in rat brain. J Pharmacol Exp Ther 315:265–272
Assié MB, Bardin L, Depoortere R, Carilla-Durand E, Newman-Tancredi A (2006) F15599, a highly selective serotonin 5HT1A receptor agonist: in vivo profile in neurochemical and behavioural models of serotonergic activity. Eur Neuropsychopharmacol 16(Suppl 4):S233
Bardin L, Kleven MS, Barret-Grevoz C, Depoortere R, Newman-Tancredi A (2006) Antipsychotic-like vs cataleptogenic actions in mice of novel antipsychotics having D2 antagonist and 5-HT1A agonist properties. Neuropsychopharmacology 31:1869–1879
Berg KA, Maayani S, Goldfarb J, Scaramellini C, Leff P, Clarke WP (1998) Effector pathway-dependent relative efficacy at serotonin type 2A and 2C receptors: evidence for agonist-directed trafficking of receptor stimulus. Mol Pharmacol 54:94–104
Bortolozzi A, Artigas F (2003) Control of 5-hydroxytryptamine release in the dorsal raphe nucleus by the noradrenergic system in rat brain. Role of alpha-adrenoceptors. Neuropsychopharmacology 28:421–434
Bortolozzi A, Amargos-Bosch M, Adell A, Diaz-Mataix L, Serrats J, Pons S, Artigas F (2003) In vivo modulation of 5-hydroxytryptamine release in mouse prefrontal cortex by local 5-HT(2A) receptors: effect of antipsychotic drugs. Eur J Neurosci 18:1235–1246
Bortolozzi A, Amargós-Bosch M, Toth M, Artigas F, Adell A (2004) In vivo release of serotonin in the dorsal raphe nucleus of 5-HT1Aeceptor knockout mice. J Neurochem 88:1373–1379
Breier A, Su TP, Saunders R, Carson RE, Kolachana BS, de Bartolomeis A, Weinberger DR, Weisenfeld N, Malhotra AK, Eckelman WC, Pickar D (1997) Schizophrenia is associated with elevated amphetamine-induced synaptic dopamine concentrations: evidence from a novel positron emission tomography method. Proc Natl Acad Sci U S A 94:2569–2574
Bruins-Slot LA, De Vries L, Newman-Tancredi A, Cussac D (2006) Differential profile of antipsychotics at serotonin 5-HT1A and dopamine D-2S receptors coupled to extracellular signal-regulated kinase. Eur J Pharmacol 534:63–70
Burris KD, Molski TF, Xu C, Ryan E, Tottori K, Kikuchi T, Yocca FD, Molinoff PB (2002) Aripiprazole, a novel antipsychotic, is a high-affinity partial agonist at human dopamine D2 receptors. J Pharmacol Exp Ther 302:381–389
Celada P, Paladini CA, Tepper JM (1999) GABAergic control of rat substantia nigra dopaminergic neurons: role of globus pallidus and substantia nigra pars reticulata. Neuroscience 89:813–825
Celada P, Puig MV, Casanovas JM, Guillazo G, Artigas F (2001) control of raphe serotonin neurons by the medial prefrontal cortex. Involvement of serotonin 1A, GABA (A) and glutamate receptors. J Neurosci 21:9917–9924
Chiodo LA (1988) Dopamine-containing neurons in the mammalian central nervous system: electrophysiology and pharmacology. Neurosci Biobehav Rev 12:49–91
Chiodo LA, Bunney BS (1983) Typical and atypical neuroleptics: differential effects of chronic administration on the activity of A9 and A10 midbrain dopaminergic neurons. J Neurosci 3:1607–1619
Cosi C, Carilla-Durand E, Assié MB, Ormiere AM, Maraval M, Leduc N, Newman-Tancredi A (2005) Partial agonist properties of the antipsychotics SSR181507, aripiprazole and bifeprunox at dopamine D2 receptors: G protein activation and prolactin release. Eur J Pharmacol 535:135–44
Cox RF, Meller E, Waszczak BL (1993) Electrophysiological evidence for a large receptor reserve for inhibition of dorsal raphe neuronal firing by 5-HT(1A) agonists. Synapse 14:297–304
Dahan L, Mnie-Filali M, Arnt J, Hertel P, Hadjeri N (2006) Effects of bifeprunox on the rat ventral tegmental area dopamine and dorsal raphe serotonin neuronal activity. Eur Neuropsychopharmacol 16(Suppl 4):S429
DeLeon A, Patel NC, Crismon ML (2004) Aripiprazole: a comprehensive review of its pharmacology, clinical efficacy, and tolerability. Clin Ther 26:649–666
Devivo M, Maayani S (1985) Inhibition of forskolin-stimulated adenylate-cyclase activity by 5-HTreceptor agonists. Eur J Pharmacol 119:231–234
Díaz-Mataix L, Scorza MC, Bortolozzi A, Toth M, Celada P, Artigas F (2005) Involvement of 5-HT1A receptors in prefrontal cortex in the modulation of dopaminergic activity. Role in atypical antipsychotic action. J Neurosci 25:10831–10843
Díaz-Mataix L, Artigas F, Celada P (2006) Activation of pyramidal cells in rat medial prefrontal cortex projecting to ventral tegmental area by a 5-HT1A receptor agonist. Eur Neuropsychopharmacol 16:288–296
Elvevag B, Goldberg TE (2000) Cognitive impairment in schizophrenia is the core of the disorder. Crit Rev Neurobiol 14:1–21
Fa M, Mereu G, Ghiglieri V, Meloni A, Salis P, Gessa GL (2003) Electrophysiological and pharmacological characteristics of nigral dopaminergic neurons in the conscious, head-restrained rat. Synapse 48:1–9
Ferré S, Cortes R, Artigas F (1994) Dopaminergic regulation of the serotonergic raphe-striatal pathway: microdialysis studies in freely moving rats. J Neurosci 14:4839–4846
Franklin KBJ, Paxinos G (1997) The mouse brain in stereotaxic coordinates. Academic, San Diego
Gessa GL, Devoto P, Diana M, Flore G, Melis M, Pistis M (2000) Dissociation of haloperidol, clozapine, and olanzapine effects on electrical activity of mesocortical dopamine neurons and dopamine release in the prefrontal cortex. Neuropsychopharmacology 22:642–649
Gobert A, Rivet JM, Cistarelli L, Melon C, Millan MJ (1999) Buspirone modulates basal and fluoxetine-stimulated dialysate levels of dopamine, noradrenaline and serotonin in the frontal cortex of freely moving rats: activation of serotonin(1A) receptors and blockade of alpha(2)-adrenergic receptors underlie its actions. Neuroscience 93:1251–1262
Goldner EM, Hsu L, Waraich P, Somers JM (2002) Prevalence and incidence studies of schizophrenic disorders: a systematic review of the literature. Can J Psychiatry 47:833–843
Grace AA, Bunney BS (1984) The control of firing pattern in nigral dopamine neurons: burst firing. J Neurosci 4:2877–2890
Green B (2004) Focus on aripiprazole. Curr Med Res Opin 20:207–213
Grunder G, Carlsson A, Wong DF (2003) Mechanism of new antipsychotic medications—occupancy is not just antagonism. Arch Gen Psychiatry 60:974–977
Harvey PD, Keefe RSE (2001) Studies of cognitive change in patients with schizophrenia following novel antipsychotic treatment. Am J Psychiatry 158:176–184
Hoyer D, Boddeke HGWM (1993) Antagonists, agonists, partial agonists: dilemmas of definition. Trends Pharmacol Sci 14:270–275
Ichikawa J, Ishii H, Bonaccorso S, Fowler WL, OLaughlin IA, Meltzer HY (2001) 5-HT2A and D-2 receptor blockade increases cortical DA release via 5-HT1A receptor activation: a possible mechanism of atypical antipsychotic-induced cortical dopamine release. J Neurochem 76:1521–1531
Jordan S, Koprivica V, Chen R, Tottori K, Kikuchi T, Altar CA (2002) The antipsychotic aripiprazole is a potent, partial agonist at the human 5-HT1A receptor. Eur J Pharmacol 441:137–140
Jordan S, Koprivica V, Dunn R, Tottori K, Kikuchi T, Altar CA (2004) In vivo effects of aripiprazole on cortical and striatal dopaminergic and serotonergic function. Eur J Pharmacol 483:45–53
Kapur S, Zipursky R, Jones C, Remington G, Houle S (2000) Relationship between dopamine D-2 occupancy, clinical response, and side effects: a double-blind PET study of first-episode schizophrenia. Am J Psychiatry 157:514–520
Kikuchi T, Tottori K, Uwahodo Y, Hirose T, Miwa T, Oshiro Y, Morita S (1995) 7-(4-[4-(2,3-Dichlorophenyl)-1-piperazinyl]butyloxy)-3,4-dihydro-2(1H)-quinolinone (OPC-14597), a new putative antipsychotic drug with both presynaptic dopamine autoreceptor agonistic activity and postsynaptic D2 receptor antagonistic activity, J Pharmacol Exp Ther 274:329–336
Laruelle M, Abi-Dargham A, van Dyck CH, Gil R, D’Souza CD, Erdos J, McCance E, Rosenblatt W, Fingado C, Zoghbi SS, Baldwin RM, Seibyl JP, Krystal JH, Charney DS, Innis RB (1996) Single photon emission computerized tomography imaging of amphetamine-induced dopamine release in drug-free schizophrenic subjects. Proc Natl Acad Sci U S A 93:9235–9240
Lawler CP, Prioleau C, Lewis MM, Mak C, Jiang D, Schetz JA, Gonzalez AM, Sibley DR, Mailman RB (1999) Interactions of the novel antipsychotic aripiprazole (OPC-14597) with dopamine and serotonin receptor subtypes. Neuropsychopharmacology 20:612–627
Lejeune F, Millan MJ (1998) Induction of burst firing in ventral tegmental area dopaminergic neurons by activation of serotonin (5-HT)(1A) receptors: WAY 100, 635-reversible actions of the highly selective ligands, flesinoxan and S-15535. Synapse 30:172–180
Lewis DA, Lieberman JA (2000) Catching up on schizophrenia: natural history and neurobiology. Neuron 28:325–334
Li Z, Ichikawa J, Dai J, Meltzer HY (2004) Aripiprazole, a novel antipsychotic drug, preferentially increases dopamine release in the prefrontal cortex and hippocampus in rat brain. Eur J Pharmacol 493:75–83
Markstein R, Hoyer D, Engel G (1986) 5-HT1A-receptors mediate stimulation of adenylate cyclase in rat hippocampus. Naunyn Schmiedebergs Arch Pharmacol 333:335–341
Martín-Ruiz R, Puig MV, Celada P, Shapiro D, Roth BL, Mengod G, Artigas F (2001a) Control of serotonergic function in medial prefrontal cortex by serotonin-2A receptors through a glutamate-dependent mechanism. J Neurosci 21:9856–9866
Martín-Ruiz R, Ugedo L, Honrubia MA, Mengod G, Artigas F (2001b) Control of serotonergic neurons in rat brain by dopaminergic receptors outside the dorsal raphe nucleus. J Neurochem 77:762–775
Meltzer HY (1999) The role of serotonin in antipsychotic drug action. Neuropsychopharmacology 21:S106–S115
Millan MJ (2000) Improving the treatment of schizophrenia: focus on serotonin (5-HT1A) receptors. J Pharmacol Exp Ther 295:853–861
Millan MJ, Newman-Tancredi A, Duqueyroix D, Cussac D (2001) Agonist properties of pindolol at h5-HT1A receptors coupled to mitogen-activated protein kinase. Eur J Pharmacol 424:13–17
Momiyama T, Amano T, Todo N, Sasa M (1996) Inhibition by a putative antipsychotic quinolinone derivative (OPC-14597) of dopaminergic neurons in the ventral tegmental area. Eur J Pharmacol 310:1–8
Newcomer JW (2005) Second-generation (atypical) antipsychotics and metabolic effects—a comprehensive literature review. CNS Drugs 19:1–93
Newman-Tancredi A, Assié MB, Cosi C, Bruins Slot L, Cussac D, Martel JC (2006) F15599, a highly selective serotonin 5HT1A receptor agonist: in vitro affinity and efficacy profile at native rat and recombinant human 5HT1A receptors. Eur Neuropsychopharmacol 16(Suppl 4):S232
Parks CL, Robinson PS, Sibille E, Shenk T, Toth M (1998) Increased anxiety of mice lacking the serotonin(1A) receptor. Proc Natl Acad Sci U S A 95:10734–10739
Paxinos G, Watson C (1998) The rat brain in stereotaxic coordinates. Academic, Sydney
Puig MV, Artigas F, Celada P (2005) Modulation of the activity of pyramidal neurons in rat prefrontal cortex by raphe stimulation in vivo: involvement of serotonin and GABA. Cereb Cortex 15:1–14
Rollema H, Lu Y, Schmidt AW, Zorn SH (1997) Clozapine increases dopamine release in prefrontal cortex by 5-HT1A receptor activation. Eur J Pharmacol 338:R3–R5
Rollema H, Lu Y, Schmidt AW, Sprouse JS, Zorn SH (2000) 5-HT(1A) receptor activation contributes to ziprasidone-induced dopamine release in the rat prefrontal cortex. Biol Psychiatry 48:229–237
Rouquier L, Claustre Y, Benavides J (1994) Alpha-1 adrenoceptor antagonists differentially control serotonin release in the hippocampus and striatum: a microdialysis study. Eur J Pharmacol 261:59–64
Santana N, Bortolozzi A, Serrats J, Mengod G, Artigas F (2004) Expression of Serotonin(1A) and Serotonin(2A) receptors in pyramidal and GABAergic neurons of the rat prefrontal cortex. Cereb Cortex 14:1100–1109
Schoeffter P, Bobirnac I, Boddeke HWGM, Hoyer D (1997) Agonist/antagonist interactions with cloned human 5-HT1A receptors: studies in transfected HeLa cells. Neuropharmacology 36:429–438
Semba J, Watanabe A, Kito S, Toru M (1995) Behavioural and neurochemical effects of OPC-14597, a novel antipsychotic drug, on dopaminergic mechanisms in rat brain. Neuropharmacology 34:785–791
Shapiro DA, Renock S, Arrington E, Chiodo LA, Liu LX, Sibley DR, Roth BL, Mailman R (2003) Aripiprazole, a novel atypical antipsychotic drug with a unique and robust pharmacology. Neuropsychopharmacology 28:1400–1411
Sprouse JS, Aghajanian GK (1988) Responses of hippocampal pyramidal cells to putative serotonin 5-HT1A and 5-HT1B agonists: a comparative study with dorsal raphe neurons. Neuropharmacology 27:707–715
Sprouse JS, Reynolds LS, Braselton JP, Rollema H, Zorn SH (1999) Comparison of the novel antipsychotic ziprasidone with clozapine and olanzapine: Inhibition of dorsal raphe cell firing and the role of 5-HT1A receptor activation. Neuropsychopharmacology 21:622–631
Sumiyoshi T, Matsui M, Yamashita I, Nohara S, Kurachi M, Uehara T, Sumiyoshi S, Sumiyoshi C, Meltzer HY (2001) The effect of tandospirone, a serotonin(1A) agonist, on memory function in schizophrenia. Biol Psychiatry 49:861–868
Sumiyoshi T, Matsui M, Yamashita I, Nohara S, Uehara T, Kurachi M, Meltzer HY (2000) Effect of adjunctive treatment with serotonin-1A agonist tandospirone on memory functions in schizophrenia. J Clin Psychopharmacol 20:386–388
Tadori Y, Miwa T, Tottori K, Burris KD, Stark A, Mori T, Kikuchi T (2005) Aripiprazole’s low intrinsic activities at human dopamine D2L and D2S receptors render it a unique antipsychotic. Eur J Pharmacol 515:10–19
Tamminga CA (2002) Partial dopamine agonists in the treatment of psychosis. J Neural Transm 109:411–420
Vandermaelen CP, Aghajanian GK (1983) Electrophysiological and pharmacological characterization of serotonergic dorsal raphe neurons recorded extracellularly and intracellularly in rat brain slices. Brain Res 289:109–119
Weinberger DR, Aloia MS, Goldberg TE, Berman KF (1994) The frontal lobes and schizophrenia. J Neuropsychiatry Clin Neurosci 6:419–427
Williams GV, Goldman-Rakic PS (1995) Modulation of memory fields by dopamine D1 receptors in prefrontal cortex. Nature 376:572–575
Yokoi F, Grunder G, Biziere K, Stephane M, Dogan AS, Dannals RF, Ravert H, Suri A, Bramer S, Wong DF (2002) Dopamine D2 and D3 receptor occupancy in normal humans treated with the antipsychotic drug aripiprazole (OPC 14597): a study using positron emission tomography and [11C]raclopride. Neuropsychopharmacology 27:248–259
Zocchi A, Fabbri D, Heidbreder CA (2005) Aripiprazole increases dopamine but not noradrenaline and serotonin levels in the mouse prefrontal cortex. Neurosci Lett 387:157–161
Acknowledgment
This work was supported by grants from the Spanish Ministry of Education and Science (SAF 2004-05525) and Bristol Myers Squibb. PC and AB are recipients of a Ramón y Cajal contract from the Ministry of Science and Technology. LDM is recipient of a predoctoral fellowship from IDIBAPS. Support from the Spanish Ministry of Health, Instituto de Salud Carlos III, Red de Enfermedades Mentales (REM-TAP Network) is also acknowledged. We thank Leticia Campa and Judith Ballart for skilful technical assistance.
Author information
Authors and Affiliations
Corresponding author
Additional information
A. Bortolozzi and L. Díaz-Mataix have contributed equally to this study.
Rights and permissions
About this article
Cite this article
Bortolozzi, A., Díaz-Mataix, L., Toth, M. et al. In vivo actions of aripiprazole on serotonergic and dopaminergic systems in rodent brain. Psychopharmacology 191, 745–758 (2007). https://doi.org/10.1007/s00213-007-0698-y
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00213-007-0698-y