
Abstract
Rationale
A growing number of studies suggest that γ-aminobutyric acid type-B (GABAB) receptor agonists reduce alcohol use and craving.
Objectives
This study was designed to further clarify behavioral mechanism(s) by which GABAB agonists may decrease alcohol reinforcement.
Methods
Male C57BL/6 J mice were trained to lever press on a concurrent schedule of ethanol (10% v/v) and water reinforcement during 16-h overnight sessions. Effects of the GABAB agonist baclofen (0–17 mg/kg, IP) or SKF 97541 (0–1 mg/kg, IP) were examined on parameters of self-administration. Subsequently, potential motor inhibition and interaction with ethanol-induced sedation by GABAB agonists was examined in ethanol naive and self-administering mice.
Results
Baclofen (10 mg/kg) and SKF 97541 (0.3 mg/kg) reduced ethanol-reinforced responding. In a locomotor activity test, these doses of the GABAB agonists inhibited locomotion in the ethanol-experienced mice and in a group of ethanol-inexperienced mice; locomotor suppression was greater in the ethanol-inexperienced mice. These doses of the GABAB agonists also potentiated the sedative effects of ethanol (4 g/kg) and converted a nonsedative dose of ethanol (2 g/kg) into a fully sedative dose. GABAB agonist enhancement of the sedative effects of ethanol was less pronounced in ethanol self-administering mice, suggesting cross-tolerance at the low dose of ethanol.
Conclusions
GABAB agonists decrease the reinforcing effects of ethanol at doses that inhibit locomotor activity and potentiate the sedative hypnotic effects of ethanol. These nonspecific effects of GABAB agonists were reduced in alcohol experienced mice, suggesting cross-tolerance to the inhibitory properties of GABAB positive modulation. These data question the safety of prescribing GABAB agonists to alcoholics since these drugs may potentiate ethanol’s sedative/hypnotic effects during relapse.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
A growing number of preclinical and clinical studies have suggested that γ-aminobutyric acid type-B (GABAB) receptor agonists reduce self-administration of a variety of drugs of abuse, including cocaine, heroin, nicotine, and alcohol (for a review see Cousins et al. 2002). For example, in rats, treatment with baclofen has been shown to reduce ethanol consumption in two-bottle home-cage tests (Daoust et al. 1987; Colombo et al. 2000, 2002; but see Smith et al. 1992, 1999). Also, in an operant self-administration task, baclofen has been shown to reduce self-administration of sweetened ethanol versus sucrose (Petry 1997), ethanol or sucrose alone (Anstrom et al. 2003), and responding for ethanol during extinction (Colombo et al. 2003b). Preliminary clinical studies have shown baclofen to reduce craving in alcoholics (Addolorato et al. 2000, 2002).
Despite this evidence showing potential preclinical and clinical efficacy of baclofen in the treatment of alcoholism, the mechanism(s) by which GABAB receptor activation may alter ethanol consumption are not fully clarified. Baclofen does not appear to alter the conditioned rewarding effects of ethanol as measured by conditioned place preference (Chester and Cunningham 1999). Other evidence indicates that baclofen does not substitute for the discriminative stimulus effects of ethanol (Shelton and Balster 1994), indicating that drug substitution may not contribute to the alteration of ethanol consumption by GABAB receptor modulation.
GABAB agonists are known to have sedative and muscle relaxant properties, and, in fact, baclofen is used clinically for the treatment of muscle spasticity. Therefore, reductions in self-administration could be due to the nonspecific actions of these agonists and not necessarily to decreases in the reinforcing nature of the drug of abuse. Addressing the sedative effects of baclofen are especially pertinent in alcohol self-administration studies given that, at certain doses, both baclofen and ethanol alone can have sedative and motor impairing/locomotor depressing properties (Lewis et al. 1989; De Luca and Massotti 1990; Middaugh et al. 1992; Ågmo and Soria 1997; Munzar et al. 2000). Thus, co-administration of baclofen and alcohol could potentially result in enhancement of the sedative properties of either compound. For example, baclofen treatment significantly enhanced the duration of ethanol-induced sedation, as measured by the loss of righting reflex (Martz et al. 1983), and baclofen has been shown to reduce ethanol-induced locomotor stimulation (Cott et al. 1976; Chester and Cunningham 1999). Moreover, co-abuse of baclofen and ethanol can produce coma (VanDierendonk and Dire 1999). Therefore, at least one mechanism by which baclofen can affect ethanol self-administration is by enhancing the sedative or locomotor depressant properties of ethanol.
The present study was designed to further clarify mechanisms by which GABAB receptor agonists may decrease the reinforcing properties of ethanol. To accomplish this goal, the effects of the GABAB agonists baclofen and SKF 97541 were examined on parameters of operant ethanol versus water self-administration in inbred C57BL/6 J (B6) ethanol-preferring mice. Effective doses of baclofen and SKF 97541 were also examined on locomotor activity and ethanol-induced sedation to examine potential nonspecific mechanisms of GABAB agonist effects on ethanol self-administration. These supplemental assessments were made in ethanol-inexperienced and ethanol-experienced (e.g., trained in the self-administration task) mice to evaluate whether chronic ethanol exposure alters response to the agonists.
Methods
Animals
Male C57BL/6 J mice (n=16 for self-administration task; n=39 for supplemental assessments) were housed in standard Plexiglas cages with four mice per cage. Food and water were continuously available except during self-administration training. The colony room was maintained on a 12-h/12-h light/dark cycle with lights on at 0700 hours. At the beginning of agonist testing, mice in the self-administration study weighed an average of 33.0±0.52 g. Mice used for the supplemental assessments weighed an average of 27.4±0.39 g. All experimental procedures in this study were in accordance with the “Guide for the Care and Use of Laboratory Animals” (National Research Council, National Academy Press, 1996).
Apparatus
Sixteen Plexiglas test chambers (Med Associates, Georgia, VT), measuring 15.9×14×12.7 cm with stainless-steel grid floors were used for the self-administration experiment. Each chamber was housed in a sound-attenuating cubicle equipped with a fan that provided ventilation and served to mask external noise. The left and right walls of each chamber contained one liquid receptacle and an ultra-sensitive mouse response lever (ENV-310 M, Med Associates) that could be depressed by 2 g of weight. Each lever was 1.6 cm wide and protruded 9.5 mm into the test chamber at a height of 2.2 cm above the floor. Lever-press responses activated a syringe pump (PHM-100, Med Associates) that delivered 0.01 ml solution into the receptacle in a 0.33-s period. A stimulus light located above each response lever was illuminated during pump activation. To correspond with the 12-h/12-h light/dark cycle in the animal colony room, a house light was on during the first 2 h (1700–1900 hours) and last 2 h (0700–0900 hours) of each daily 16-h session. The chambers were controlled using a computer interface and PC software (Med Associates).
Four Plexiglas chambers (28×28 cm; Med Associates) were used to measure locomotor activity. Each chamber was equipped with two sets of 16 pulse-modulated infrared photo beams (2.5 cm from the floor) and placed on opposite walls to record x–y ambulatory movements. Activity chambers were computer interfaced (Med Associates) for data sampling at 100-ms resolution.
Ethanol self-administration training
Operant ethanol self-administration was measured during 16-h sessions (between 1700 hours and 0900 hours). On the first 2 days of training, mice were weighed and individually placed into a test chamber for overnight 16-h training sessions to establish reliable lever pressing behavior. Before each of these sessions, water was removed from the home cages 24 h before the first training session and returned for 2 h following each training session. During these sessions, both levers were active on a concurrent fixed-ratio 1 (CONC FR1 FR1) schedule with 10% sucrose and water presented as the reinforcers. That is, one lever response resulted in the presentation of 0.01 ml of the solution paired with that lever. On the following days, animals were trained to orally self-administer ethanol using a sucrose substitution procedure (Samson 1986, 1987; Hodge et al. 1995). Briefly, the FR1 contingency was in place and ethanol was gradually added to the 10% sucrose solution, and the sucrose was gradually faded out so that 10% ethanol maintained lever pressing. The mice experienced four training sessions a week (Monday through Thursday). One mouse failed to perform the operant task and was eliminated from the study. At the time of GABAB agonist testing, the mice had 4 months of experience self-administering ethanol.
Effects of GABAB agonists on ethanol self-administration
Baclofen
For each 16-h test session, mice were administered baclofen (0, 1, 3, 10, or 17 mg/kg, IP) and immediately placed in the chambers. Tests sessions were conducted on Tuesday and Thursday of each week. Baclofen was administered according to a randomized Latin-Square design, with the exception that the 17-mg/kg dose was tested last in all animals. Each mouse received a single injection of each dose.
SKF 97541
One week after baclofen testing, the effects of SKF 97541 were assessed. The same testing procedures were used as described for baclofen, except that SKF 97541 (0, 0.01, 0.03, 0.1 mg/kg, IP) was administered according to a randomized Latin-Square design prior to the sessions. Two higher doses, 1 mg/kg and 0.3 mg/kg, were administered in that order after the other SKF 97541 doses were tested.
Effects of GABAB agonists on locomotor activity
After the effects of SKF 97541 were assessed on ethanol self-administration, training sessions (Monday–Thursday) were resumed for 10 weeks. Then, self-administration sessions were suspended for 2 days (Wednesday and Thursday) and mice were tested in locomotor activity procedures. To test the effects of the GABAB agonists on locomotor ability, mice (n=5/group) were administered saline, SKF 97541 (0.3 mg/kg), or baclofen (10 mg/kg) IP 20 min before placement in the activity chambers. Self-administration sessions (Monday–Thursday) were resumed after locomotor testing. In order to assess whether locomotor response to the GABAB agonists was modified by the extensive ethanol self-administration history, locomotor activity was also tested in ethanol-inexperienced mice (n=5/group) following administration of saline, SKF 97541 (0.3 mg/kg ), or baclofen (10 mg/kg). In both experiments, GABAB agonists were administered 20 min before placement in the activity chambers and performance was then monitored for 2 h. Throughout the manuscript the term “ethanol-experienced” refers to mice with a history of ethanol self-administration, and the term “ethanol-inexperienced” refers to mice that have no history of self-administration.
Effects of GABAB agonists on ethanol-induced loss of righting reflex
Sedative ethanol dose (4 g/kg)
As in the locomotor testing procedure, self-administration sessions were suspended and mice were tested for ethanol-induced sedation. To determine whether GABAB agonists interact with the sedative-hypnotic effects of ethanol, ethanol-experienced (i.e., self-administration group; n=5/group) and inexperienced (i.e., a different group of mice from the locomotor test; n=8/group) mice were pretreated with saline, SKF 97541 (0.3 mg/kg), or baclofen (10 mg/kg) and returned to the home cage. Twenty minutes after GABAB agonist administration, mice were injected with a sedative dose of ethanol (4 g/kg, IP) and intermittently placed on their backs in a V-shaped trough to test for the loss of righting reflex. Loss of righting was defined as the inability of the animal to right itself after being placed on its back within a 30-s interval. Recovery of the righting reflex was determined when the animal righted itself three times within 30 s. If a mouse did not lose the righting reflex within 30 min after the ethanol injection, that mouse was excluded from the experiment. Duration was defined as the time interval between loss and recovery of the righting reflex. Mice in the ethanol-experienced group (i.e., self-administration history) received a different drug from the previous locomotor assessment. Loss of righting reflex experiments were conducted 5 weeks after the locomotor activity assessment in the self-administration group during a 2-day period and self-administration sessions resumed on Monday.
Non-sedative ethanol dose (2 g/kg)
To further examine the involvement of GABAB receptor activation in ethanol-induced loss of righting reflex, ethanol-experienced (i.e., self-administration group; n=5/group) and the same group of mice from the previous loss of righting assessment (n=8/group) were administered saline, SKF 97541 (0.3 mg/kg), or baclofen (10 mg/kg) followed 20 min later by a non-sedative dose of ethanol (2 g/kg, IP). The same procedures were used as previously described except that the ethanol dose was 2 g/kg. All mice received a different GABAB agonist than that experienced on the previous assessment.
Drugs
For self-administration, ethanol (95% w/v) was diluted in distilled water to a concentration of 10% (v/v). For the loss of righting reflex experiments, ethanol (95% w/v) was diluted in saline (0.9%) to 20% (v/v) and injected at different volumes to achieve the appropriate dosage. Baclofen and SKF 97541 (Tocris Cookson, Ellisville, MO) were dissolved in saline (0.9%) and injected IP at a volume of 0.01 ml/g body weight.
Data analysis
The total number of ethanol- and water-reinforced responses, and responses during each hour of the 16-h self-administration sessions were recorded. Hourly totals were converted to cumulative response totals to evaluate the emergence of drug effects on total responding. Ethanol intake was converted to grams per kilogram body weight (g/kg) for each test session. Data were analyzed by within- or between-subjects analysis of variance (ANOVA) or repeated-measures ANOVA where appropriate. Post-hoc comparisons were conducted using Tukey’s procedure.
Results
Effects of GABAB agonists on ethanol self-administration
Baclofen
Average alcohol intake for the 16-h session after saline pretreatment was 3.55±0.42 g/kg/16 h. Baclofen pretreatment significantly altered alcohol intake (F 4,56=2.84, P=0.03). This significant difference was likely a result of a suppression of ethanol intake at the highest baclofen dose (17 mg/kg), compared with the 3-mg/kg baclofen dose (P<0.05), because total ethanol intake did not differ from saline pretreatment levels at any dose tested. To examine the pattern and rate of responding across the entire 16-h session, cumulative ethanol and water responses were examined separately. Figure 1A, B illustrates cumulative ethanol and water responses, respectively, after saline injection and the baclofen doses tested. Two-way repeated-measures ANOVA showed a significant effect of baclofen dose (F 4,56=8.31, P<0.001) on ethanol-reinforced responding (Fig. 1A). Treatment with the highest baclofen dose (17 mg/kg) significantly reduced ethanol-reinforced responding relative to saline treatment. A trend for a significant overall reduction in ethanol-reinforced responding was also observed at the 10-mg/kg dose (P=0.06). The main effect of hour (F 15,210=48.58, P<0.001), and the dose by hour interaction (F 60,840=2.97, P<0.001) were also significant. Examination of cumulative response plots shows that the rate of ethanol-reinforced responding was suppressed by the two highest doses of baclofen. Specifically, treatment with 17 mg/kg baclofen significantly reduced ethanol-reinforced responding relative to saline treatment beginning at hour 6 and continuing throughout the remainder of the 16-h session. Treatment with the 10-mg/kg baclofen dose also reduced ethanol-reinforced responding beginning at hour 10 and continuing until hour 15.

A Mean (±SEM) cumulative ethanol responses after saline (closed circle) and 1 mg/kg (open triangle), 3 mg/kg (open diamond), 10 mg/kg (open circle) and 17 mg/kg (open square) baclofen injection at each hour of the self-administration session. B Mean (±SEM) cumulative water responses after saline (closed circle) and 1 mg/kg (open triangle), 3 mg/kg (open diamond), 10 mg/kg (open circle) and 17 mg/kg (open square) baclofen injection at each hour of the self-administration session. †Significant difference between saline and 10 mg/kg baclofen. *Significant difference between saline and 17 mg/kg baclofen (Tukey, P<0.05)
As illustrated in Fig. 1B, baclofen significantly altered water-reinforced responding (F 4,56=7.02, P<0.001), with the highest baclofen dose significantly reducing responding relative to saline treatment. A significant effect of hour (F 15,210=21.88, P<0.001) and a significant dose by hour interaction (F 60,840=3.60, P<0.001) were also observed. Treatment with 17 mg/kg baclofen suppressed water-reinforced responding relative to saline treatment at hours 14–16; no other baclofen doses differed from the saline at any time points. Taken together, these results show that baclofen reduced ethanol-reinforced responding at a dose (10 mg/kg) that did not affect water-reinforced responding.
SKF 97541
Average alcohol intake for the 16-h session after saline pretreatment was 3.34±0.32 g/kg/16-h. Alcohol intake was altered by SKF 97541 pretreatment (F 5,70=21.22, P=<0.001), with a significant reduction in ethanol observed at the highest SKF 97541 dose (1.0 mg/kg) relative to saline pretreatment. Cumulative ethanol-reinforced responding after treatment with saline and the SKF 97541 doses is illustrated in Fig. 2A. SKF 97541 significantly altered ethanol-reinforced responding (F 5,70=32.19, P<0.001), with the highest dose (1 mg/kg) significantly suppressing ethanol-reinforced responding relative to saline treatment (P<0.001). The main effect of hour (F 15,210=92.40, P<0.001), and the dose by hour interaction (F 75,1050=13.17, P<0.001) were also significant. Examining total responses at each hour revealed that 0.3 mg/kg SKF 97541 reduced ethanol-reinforced responding at hour 10, and this response suppression continued from hour 13 to 15. The highest SKF 97541 (1.0 mg/kg) dose also suppressed ethanol-reinforced responding relative to saline treatment from hour 6 and throughout the remainder of the session.

A Mean (±SEM) cumulative ethanol responses after saline (closed circle) and 0.01 mg/kg (open triangle), 0.03 mg/kg (open diamond), 0.1 mg/kg (inverted open triangle), 0.3 mg/kg (open circle), and 1.0 mg/kg (open square) SKF 97541 injection at each hour of the self-administration session. B Mean (±SEM) cumulative water responses after saline (closed circle) and 0.01 mg/kg (open triangle), 0.03 mg/kg (open diamond), 0.1 mg/kg (inverted open triangle), 0.3 mg/kg (open circle), and 1.0 mg/kg (open square) SKF 97541 injection at each hour of the self-administration session. *Significant difference from saline (Tukey, P<0.05)
SKF 97541 also affected water responses as shown in Fig. 2B. There was a significant main effect of dose (F 5,70=6.23, P<0.001), with the highest SKF 97541 dose suppressing water-reinforced responding. The main effect of hour (F 15,210=21.28) and the dose by hour interaction (F 75,1050=5.36) were also significant (P<0.001). The highest SKF 97541 dose (1.0 mg/kg) significantly altered water responding. That is, at hour 9 and throughout the remainder of the session, the cumulative water responses were significantly less than those after saline treatment. SKF 97541 did not affect cumulative water responses at any other dose. Thus, similar to baclofen treatment, ethanol-reinforced responding was reduced by a dose that did not affect water-reinforced responding (0.3 mg/kg).
Locomotor activity
In order to investigate the effects of the GABAB agonists on locomotor activity of the ethanol-experienced (i.e., self-administration history) and ethanol-inexperienced groups, a three-way ANOVA with time as a repeated factor was conducted. This analysis revealed a significant difference in the locomotor activity of the ethanol-experienced and inexperienced groups (F 1,24=14.26, P=0.001). There was also a significant change in activity across the 2-h session (F 11,264=4.27, P<0.001) with the GABAB agonists reducing locomotor activity (F 2,24=19.88, P<0.001). Further, there was a significant interaction between time interval and GABAB agonist treatment (F 22,264=15.23, P<0.001), and a significant interaction between the groups and time interval (F 11,264=10.90, P<0.001).
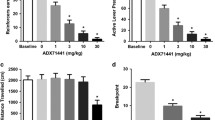
To further investigate the significant time by agonist, and the group by time interaction, a two-way ANOVA with time as a repeated measure was conducted for both groups. As illustrated in Fig. 3A, GABAB agonist pretreatment altered the pattern of activity in ethanol-experienced mice relative to saline control (F 2,12=5.30, P=0.02). Specifically, baclofen significantly reduced activity relative to the saline group (inset of Fig. 3A). An overall change in activity throughout the session was observed (F 11,132=5.69, P<0.001). Post-hoc analyses prompted by the significant drug by time interval interaction (F 22,132=5.06, P<0.001), revealed that baclofen (10 mg/kg) suppressed locomotor activity relative to the saline-treated controls for the first hour of the session. Locomotor suppression was also observed after SKF 97541 (0.3 mg/kg) treatment, but only for the first 30 min of the session.

A Mean (±SEM) distance traveled in centimeters in 10-min intervals after saline, SKF 97541 and baclofen injection in the ethanol-experienced group (i.e., self-administration history) (n=5/drug). Inset graph illustrates the total distance traveled in the 2-h session. B Mean (±SEM) distance traveled in centimeters in 10-min intervals after saline, SKF 97541 and baclofen injection in the ethanol-inexperienced group (n=5/drug). Inset graph illustrates the total distance traveled in the 2-h session. †Significant difference between saline (closed circle) and 0.3 mg/kg SKF 97541 (open circle). *Significant difference between saline and 10 mg/kg baclofen (open square) (Tukey, P<0.05)
The pattern of activity across the 2-h session for the ethanol-inexperienced group is illustrated in Fig. 3B. Baclofen (10 mg/kg) and SKF 97541 (0.3 mg/kg) significantly reduced locomotor activity as assessed by the total distance traveled during the 2-h test (F 2,12=21.76, P<0.001; inset of Fig. 3B). The main effect of time (F 11,132=15.30, P<0.001), and the drug by time interval interaction was also significant (F 22,132=20.14, P<0.001). Mice treated with baclofen and SKF 97541 were less active than the saline-treated mice throughout the entire 2-h session, except at the tenth 10-min interval. This data pattern is in contrast to that observed in the self-administration group given that baclofen suppressed locomotor activity in that group for the first hour and SKF 97541 suppressed activity for the first 30 min of the session. Thus, mice with extended ethanol exposure (i.e., self-administration group) showed less sensitivity to the locomotor suppressant effects of baclofen and SKF 97541 than ethanol-inexperienced mice.
Loss of righting reflex
Sedative ethanol dose (4 g/kg)
Duration of the loss of righting reflex in the ethanol-experienced (i.e., self-administration history) and ethanol-inexperienced groups is illustrated in the right and left panels of Fig. 4A, respectively. One mouse in the ethanol-inexperienced group (saline pretreatment condition) did not lose the righting reflex and was excluded from the analyses. When administered alone, baclofen (10 mg/kg) or SKF 97541 (0.3 mg/kg) did not induce loss of righting reflex. In contrast, both GABAB agonists significantly enhanced the duration of the loss of righting reflex induced by ethanol (F 2,32=58.55, P<0.001). SKF 97541 (0.3 mg/kg) pretreatment resulted in a 2.5-fold increase in the duration of the loss of righting reflex and baclofen increased the duration by threefold relative to ethanol (4 g/kg) alone (i.e., saline pretreatment). Effects of the GABAB agonists on ethanol-induced loss of righting reflex did not differ as a function of ethanol experience, and no significant interaction between the two groups and agonist treatment was observed. That is, previous ethanol exposure did not alter the effects of baclofen or SKF 97541 on ethanol-induced sedation.

A Mean (±SEM) duration of loss of righting reflex (LORR) induced by a sedative ethanol dose (4 g/kg) in the ethanol-inexperienced (n=23) and ethanol-experienced (i.e., self-administration history) (n=15). Mice received a pretreatment injection of saline (Sal), 0.3 mg/kg SKF 97541 (SKF), or 10 mg/kg baclofen (Bac) 20 min before the ethanol injection. B Mean (±SEM) duration of LORR after injection of a non-sedative ethanol dose (2 g/kg) in the ethanol-inexperienced (n=24) and ethanol-experienced (i.e., self-administration history) groups (n=15). Mice received a pretreatment injection of Sal, 0.3 mg/kg SKF, or 10 mg/kg Bac 20 min before the ethanol injection. *Significant difference from the respective Sal condition of each group (Tukey, P<0.05)
Non-sedative ethanol dose (2 g/kg)
The effect of a lower dose of ethanol (2 g/kg) was also examined alone and in combination with both GABAB agonists in ethanol-experienced and ethanol-inexperienced mice (Fig. 4B). In both ethanol experience groups, the 2-g/kg ethanol dose alone (i.e., saline pretreatment) did not induce the loss of righting reflex in any of the animals tested; however, baclofen (10 mg/kg) and SKF 97541 (0.3 mg/kg) potentiated the effects of ethanol (2 g/kg) to induce loss of righting reflex (F 2,33=29.88, P<0.001). The ethanol-inexperienced group was more sensitive to potentiation by the GABAB agonists than the self-administration group (F 1,33=17.83, P<0.001). Further, there existed a significant interaction (F 2,33=5.62, P=0.008) between ethanol experience and drug treatment. That is, post-hoc comparisons showed that both GABAB agonists significantly enhanced ethanol (2 g/kg)-induced loss of righting to a greater extent in the ethanol-inexperienced group than the ethanol-experienced group (P<0.001 for SKF 97541, P<0.02 for baclofen). In addition, SKF 97541 pretreatment significantly enhanced loss of righting relative to mice pretreated with saline (P<0.001) only in the ethanol-inexperienced group. However, baclofen potentiation of the loss of righting reflex was significantly greater than saline pretreatment in both groups (P<0.002).
Discussion
A growing literature suggests that the GABAB agonist baclofen may be useful in the medical management of problems associated with alcohol and drug abuse (Cousins et al. 2002). Preclinical studies have shown that baclofen reduces alcohol intake (Daoust et al. 1987; Colombo et al. 2000, 2002), responding during extinction (Colombo et al. 2003b), and elevated intake associated with the alcohol deprivation effect (Colombo et al. 2003a), which is a rodent model of relapse to alcohol drinking (McBride et al. 2002). In the present study, effects of the GABAB agonists baclofen and SKF 97541 were tested in ethanol-preferring inbred C57BL/6 J mice trained to concurrently self-administer ethanol versus water during 16-h sessions. Both GABAB agonists selectively reduced ethanol-reinforced responding at a dose that did not alter water-reinforced responding (e.g., 10 mg/kg baclofen; 0.3 mg/kg SKF 97541). Higher doses of the GABAB agonists, however, reduced both ethanol- and water-reinforced responding, suggesting nonspecific effects on motor ability or general reinforcement mechanisms at higher agonist doses. Accordingly, the doses of baclofen and SKF 97541 that selectively decreased ethanol-reinforced responding also reduced locomotor activity and enhanced the sedative effects of ethanol. The preclinical profile that emerges from these studies is that GABAB agonists may reduce alcohol self-administration to some extent by inhibiting locomotor ability and/or by enhancing the pharmacological effects of ethanol.
The selective reduction of ethanol-reinforced responding produced by baclofen and SKF 97541 in C57BL/6 J mice is consistent with previous work showing selective reductions in ethanol self-administration by rats under concurrent reinforcement conditions (Petry 1997). In that study, rats had access to two response levers; responses on one lever resulted in the delivery of a sweetened ethanol solution (10% ethanol/10% sucrose), while responses on the other lever resulted in the delivery of an isocaloric sucrose (24%) solution. Baclofen selectively reduced sweetened ethanol-reinforced responding at a dose that did not affect sucrose responding (2.5 mg/kg). Similarly, baclofen produced a dose-dependent reduction in voluntary ethanol intake in a two-bottle choice test with a compensatory increase in water intake by Sardinian alcohol-preferring (Sp) rats (Colombo et al. 2000). Together, these data indicate that GABAB agonists selectively and dose dependently decrease ethanol self-administration in a variety of animal models and behavioral conditions.
The GABAB agonists decreased ethanol-reinforced responding by C57BL/6 J mice in a dose- and time-dependent manner. Doses of baclofen (e.g., 1–3 mg/kg) that have been shown to decrease sweetened ethanol-reinforced responding by rats (Petry 1997) were without effect on ethanol (10% v/v)-reinforced responding during the 16-h sessions. Higher doses of each agonist significantly suppressed response rate only after several hours into the session. This time-dependent effect is likely due to the pattern of mouse operant behavior during the light and dark phases of the 16-h sessions. The first 2 h of these sessions were during the light cycle, and very little behavior occurred on either the water- or ethanol-associated lever. Response rate increased during the initial stages of the dark cycle (i.e., after hour 2) as evidenced by an increase in the slope of the cumulative response plots, but response totals remained relatively low, which precluded seeing a statistical difference after GABAB agonist treatment. At later time points (i.e., after hour 6), significant effects of the response rate suppressing effect of the agonists emerged as response totals increased (Fig. 1A and Fig. 2A). Importantly, the pharmacological effects of the agonists likely extended into the dark cycle, given that the concentration of baclofen after IV administration is present in rat plasma and brain after 6 h (Faigle and Keberle 1972; Mandema et al. 1992), and the commonly reported elimination half-life for baclofen in humans is up to 6 h (Shellenberger et al. 1999). The rate-suppressing effects of baclofen and SKF 97541 were diminished during the last hour of the sessions (i.e., hour 16). These data indicate that doses of the GABAB agonists selectively suppressed ethanol-reinforced responding by slowing operant response rate during the dark phase of the circadian cycle.
The highest dose of baclofen or SKF 97541 tested in the present study decreased both ethanol- and water-reinforced responding, indicating loss of specificity for ethanol reinforcement. It is important to consider issues of specificity when interpreting decreases in ethanol self-administration because baclofen is known to reduce locomotor activity (Levy 1977; Gianutsos 1978; Rodgers and Depaulis 1982; Agmo and Giordano 1985; Mead and Little 1995; Broadbent and Harless 1999), food intake (Zarrindast et al. 1989), and food-reinforced operant response rate (Grech and Balster 1993). Indeed, similar nonspecific reductions of sweetened ethanol- and concurrent sucrose-reinforced responding (Petry 1997) as well as reductions of both ethanol- and sucrose-reinforced responding when presented separately (Anstrom et al. 2003) have been reported.
To determine whether GABAB agonist-induced reductions in ethanol self-administration were associated with locomotor inhibition, effects of the agonists were assessed on spontaneous open-field locomotor activity by ethanol-experienced and ethanol-inexperienced mice. The doses that selectively reduced ethanol self-administration (10 mg/kg baclofen; 0.3 mg/kg SKF 97541) suppressed locomotor activity throughout the entire 2-h test session in ethanol-inexperienced mice. Interestingly, while the same agonist doses suppressed activity in the ethanol-experienced group, the extent of the suppression was much less severe than that observed in the inexperienced group. That is, locomotor activity returned to baseline levels by the first 30 min after SKF 97541 administration and by 1 h after baclofen administration. A plausible explanation for the differential effect of the agonists in the two groups is that chronic ethanol exposure reduced sensitivity to the suppressant effects of the GABAB agonists. This is supported by electrophysiological data in which hippocampal slices from rats chronically treated with ethanol showed less baclofen-induced hyperpolarization relative to ethanol-naive rats (i.e., less sensitivity; Frye et al. 1991). Importantly, both GABAB agonists administered alone induced acute motor impairing effects, but given that locomotor activity returned to control levels by the first hour of the test session, it is possible that in the self-administration task ethanol-reinforced responding was not significantly altered by an agonist-induced motor impairment. Thus, the motor-impairing effects of GABAB agonists may be attenuated by a history of ethanol consumption.
An important factor to consider is that the GABAB agonists may have interacted with the self-administered ethanol to produce motor inhibition. For example, baclofen-induced locomotor suppression (7.5 mg/kg) was significantly enhanced by co-administration of ethanol (2 g/kg) (Broadbent and Harless 1999). Similarly, baclofen (5 mg/kg and 7.5 mg/kg) has been shown to significantly reduce ethanol-induced locomotor stimulation (Chester and Cunningham 1999; Cott et al. 1976). Further, Martz et al. (1983) found that motor incoordinating effects of baclofen were potentiated by ethanol administration. Thus, the possibility exists that self-administered ethanol interacted with baclofen and SKF 97541 and contributed to the reduction in ethanol-reinforced responding.
Another potential mechanism for the GABAB agonist-induced reduction in ethanol-reinforced responding may be an increase of the sedative properties of ethanol. In the present study, SKF 97541 (0.3 mg/kg) and baclofen (10 mg/kg) increased the duration of the loss of righting reflex induced by a 4-g/kg dose of ethanol by 2- and 3-fold, respectively. This striking increase in ethanol-induced sedation with the 4-g/kg dose was evident regardless of the previous ethanol experience of the mice (e.g., self-administration and ethanol-naive groups did not differ in duration). These results are consistent with Martz et al. (1983), and extend the findings of that work to show that another GABAB agonist SKF 97541 also interacts with ethanol to increase the duration of the loss of righting reflex. It is unclear, however, whether ethanol-induced sedation is directly mediated through GABAB receptor activation since the GABAB antagonist phaclofen does not attenuate the duration of ethanol-induced loss of righting reflex (Allan and Harris 1989). Alternatively, evidence indicates that combination of subthreshold concentrations of ethanol and baclofen potentiate GABAA receptor-mediated Cl− flux (Allan et al. 1991). Thus, GABAB agonists may potentiate ethanol-induced loss of righting reflex by enhancing ethanol’s effects on GABAA receptors (but see Mehta and Ticku 1990).
A novel finding of the present study is that pretreatment with non-sedative doses of baclofen or SKF 97541 induced loss of righting when combined with a non-sedative dose of ethanol (2 g/kg). This data pattern suggests that pretreatment with the agonists may have enhanced the sedative effects of self-administered ethanol resulting in disruption of ‘normal’ responding. That is, when a lower dose of ethanol was administered (2 g/kg), baclofen and SKF 97541 potentiated loss of righting in all of the mice in the ethanol-inexperienced group. In contrast, the effects were less consistent in the ethanol-experienced mice. SKF 97541 induced sedation in two of the five mice, while baclofen induced sedation in four of the five mice. This pattern suggests reduced sensitivity to the potentiation of sedation by GABAB agonists in ethanol-experienced mice, which might reflect cross-tolerance, or other adaptive mechanisms (Zaleski et al. 2001) between ethanol and the GABAB agonists. There was no evidence of cross-tolerance between ethanol and the GABAB agonists when a high dose of ethanol (4 g/kg) was used to induce sedation.
Given the evidence for pharmacological interactions between ethanol and GABAB agonists, utilizing baclofen as a potential pharmacotherapy for the treatment of alcohol disorders should raise safety concerns. That is, if an alcoholic is prescribed baclofen in an outpatient setting (access to alcohol may be readily available), a relapse episode could have detrimental effects as the sedative/hypnotic properties of alcohol may be greatly enhanced (VanDierendonk and Dire 1999). However, the data from the present study showed that alcohol-experienced mice were less sensitive to the sedative properties of the agonists than naive mice. This result suggests that the combination of baclofen and alcohol or baclofen alone would have less of a negative impact on an alcoholic than a naive drinker. Indeed, in a clinical study examining the effects of baclofen in alcoholics, two patients continued drinking during baclofen treatment and no specific mention was made of any serious side effects in those patients (Addolorato et al. 2000). Importantly, however, the effects of baclofen after a period of abstinence have not been tested and possible cross tolerance to the agonist may dissipate after periods of abstinence. Thus, before baclofen is considered as a clinical therapeutic for alcoholism, further research is warranted to understand the mechanisms involved in pharmacological interactions between ethanol and the GABAB receptor.
References
Addolorato G, Caputo F, Capristo E, Colombo G, Gessa GL, Gasbarrini G (2000) Ability of baclofen in reducing alcohol craving and intake. II. Preliminary clinical evidence. Alcohol Clin Exp Res 24:67–71
Addolorato G, Caputo F, Capristo E, Domenicali M, Bernardi M, Janiri L, Agabio R, Colombo G, Gessa GL, Gasbarrini G (2002) Baclofen efficacy in reducing alcohol craving and intake: a preliminary double-blind randomized controlled study. Alcohol Alcohol 37:504–508
Agmo A, Giordano M (1985) The locomotor-reducing effects of GABAergic drugs do not depend on the GABAA receptor. Psychopharmacology 87:51–54
Ågmo A, Soria P (1997) GABAergic drugs and sexual motivation, receptivity and exploratory behaviors in the female rat. Psychopharmacology 129:372–381
Allan AM, Harris RA (1989) A new alcohol antagonist: phaclofen. Life Sci 45:1771–1779
Allan AM, Burnett D, Harris RA (1991) Ethanol-induced changes in chloride flux are mediated by both GABA(A) and GABA(B) receptors. Alcohol Clin Exp Res 15:233–237
Anstrom KK, Cromwell HC, Markowski T, Woodward DJ (2003) Effect of baclofen on alcohol and sucrose self-administration in rats. Alcohol Clin Exp Res 27:900–908
Broadbent J, Harless WE (1999) Differential effects of GABA(A) and GABA(B) agonists on sensitization to the locomotor stimulant effects of ethanol in DBA/2J mice. Psychopharmacology 141:197–205
Chester JA, Cunningham CL (1999) Baclofen alters ethanol-stimulated activity but not conditioned place preference or taste aversion in mice. Pharmacol Biochem Behav 63:325–331
Colombo G, Agabio R, Carai MA, Lobina C, Pani M, Reali R, Addolorato G, Gessa GL (2000) Ability of baclofen in reducing alcohol intake and withdrawal severity. I. Preclinical evidence. Alcohol Clin Exp Res 24:58–66
Colombo G, Serra S, Brunetti G, Atzori G, Pani M, Vacca G, Addolorato G, Froestl W, Carai MA, Gessa GL (2002) The GABA(B) receptor agonists baclofen and CGP 44532 prevent acquisition of alcohol drinking behaviour in alcohol-preferring rats. Alcohol Alcohol 37:499–503
Colombo G, Serra S, Brunetti G, Vacca G, Carai MA, Gessa GL (2003a) Suppression by baclofen of alcohol deprivation effect in Sardinian alcohol-preferring (sP) rats. Drug Alcohol Depend 70:105–108
Colombo G, Vacca G, Serra S, Brunetti G, Carai MA, Gessa GL (2003b) Baclofen suppresses motivation to consume alcohol in rats. Psychopharmacology 167:221–224
Cott J, Carlsson A, Engel J, Lindqvist M (1976) Suppression of ethanol-induced locomotor stimulation by GABA-like drugs. Naunyn Schmiedebergs Arch Pharmacol 295:203–209
Cousins MS, Roberts DC, de Wit H (2002) GABA(B) receptor agonists for the treatment of drug addiction: a review of recent findings. Drug Alcohol Depend 65:209–220
Daoust M, Saligaut C, Lhuintre JP, Moore N, Flipo JL, Boismare F (1987) GABA transmission, but not benzodiazepine receptor stimulation, modulates ethanol intake by rats. Alcohol 4:469–472
De Luca C, Massotti M (1990) Phaclofen antagonizes the antinociceptive but not the sedative effects of (−)-baclofen. Prog Neuropsychopharmacol Biol Psychiatry 14:597–607
Faigle JW, Keberle H (1972) The chemistry of lioresal. Postgrad Med J Suppl 5:9–13
Frye GD, Taylor L, Trzeciakowski JP, Griffith WH (1991) Effects of acute and chronic ethanol treatment on pre- and postsynaptic responses to baclofen in rat hippocampus. Brain Res 560:84–91
Gianutsos GMK (1978) Tolerance to the effects of baclofen and gamma-butyrolactone on locomotor activity and dopaminergic neurons in the mouse. J Pharmacol Exp Ther 207:859–869
Grech DM, Balster RL (1993) Pentobarbital-like discriminative stimulus effects of direct GABA agonists in rats. Psychopharmacology 110:295–301
Hodge CW, Chappelle AM, Samson HH (1995) GABAergic transmission in the nucleus accumbens is involved in the termination of ethanol self-administration in rats. Alcohol Clin Exp Res 19:1486–1493
Levy RA, Proudfit HK (1977) The analgesic action of baclofen [beta-(4-chlorophenyl)-gamma-aminobutyric acid]. J Pharmacol Exp Ther 202:437–445
Lewis JL, Westerberg VS, LaBella FS (1989) Mechanisms of general anaesthesia: brain regional responses to baclofen. Physiol Behav 46:3–8
Mandema JW, Heijligers-Feijen CD, Tukker E, De Boer AG, Danhof M (1992) Modeling of the effect site equilibration kinetics and pharmacodynamics of racemic baclofen and its enantiomers using quantitative EEG effect measures. J Pharmacol Exp Ther 261:88–95
Martz A, Deitrich RA, Harris RA (1983) Behavioral evidence for the involvement of gamma-aminobutyric acid in the actions of ethanol. Eur J Pharmacol 89:53–62
McBride WJ, Le AD, Noronha A (2002) Central nervous system mechanisms in alcohol relapse. Alcohol Clin Exp Res 26:280–286
Mead AJ, Little HJ (1995) Do GABAB receptors have a role in causing behavioural hyperexcitability, both during ethanol withdrawal and in naive mice? Psychopharmacology 117:232–239
Mehta AK, Ticku MK (1990) Are GABAB receptors involved in the pharmacological effects of ethanol? Eur J Pharmacol 182:473–480
Middaugh LD, Frackelton WF, Boggan WO, Onofrio A, Shepherd CL (1992) Gender differences in the effects of ethanol on C57BL/6 mice. Alcohol 9:257–260
Munzar P, Kutkat SW, Miller CR, Goldberg SR (2000) Failure of baclofen to modulate discriminative-stimulus effects of cocaine or methamphetamine in rats. Eur J Pharmacol 408:169–174
Petry NM (1997) Benzodiazepine-GABA modulation of concurrent ethanol and sucrose reinforcement in the rat. Exp Clin Psychopharmacol 5:183–194
Rodgers RJ, Depaulis A (1982) GABAergic influences on defensive fighting in rats. Pharmacol Biochem Behav 17:451–456
Samson HH (1986) Initiation of ethanol reinforcement using a sucrose-substitution procedure in food- and water-sated rats. Alcohol Clin Exp Res 10:436–442
Samson HH (1987) Initiation of ethanol-maintained behavior: a comparison of animal models and their implication to human drinking. In: Thompson T, Dews PB, Barrett JE (eds) Adv Behav Pharmacol, vol 6. Neurobehavioral Pharmacology. Lawrence Erlbaum, Hillsdale, NJ, pp 221–248
Shellenberger MK, Groves L, Shah J, Novack GD (1999) A controlled pharmacokinetic evaluation of tizanidine and baclofen at steady state. Drug Metab Dispos 27:201–204
Shelton KL, Balster RL (1994) Ethanol drug discrimination in rats: substitution with GABA agonists and NMDA antagonists. Behav Pharmacol 5:441–451
Smith BR, Robidoux J, Amit Z (1992) GABAergic involvement in the acquisition of voluntary ethanol intake in laboratory rats. Alcohol Alcohol 27:227–231
Smith BR, Boyle AE, Amit Z (1999) The effects of the GABA(B) agonist baclofen on the temporal and structural characteristics of ethanol intake. Alcohol 17:231–240
VanDierendonk DR, Dire DJ (1999) Baclofen and ethanol ingestion: a case report. J Emerg Med 17:989–993
Zaleski MJ, Nunes Filho JR, Lemos T, Morato GS (2001) GABA(B) receptors play a role in the development of tolerance to ethanol in mice. Psychopharmacology 153:415–424
Zarrindast MR, Hosseini-Nia T, Allah-Maddadi S (1989) Food intake suppressant effect of baclofen in rats. Gen Pharmacol 20:701–703
Acknowledgements
This work was supported by grants AA09981 and AA011605 to C.W.H. from the National Institute on Alcohol Abuse and Alcoholism and by the Bowles Center for Alcohol Studies.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Besheer, J., Lepoutre, V. & Hodge, C.W. GABAB receptor agonists reduce operant ethanol self-administration and enhance ethanol sedation in C57BL/6J mice. Psychopharmacology 174, 358–366 (2004). https://doi.org/10.1007/s00213-003-1769-3
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00213-003-1769-3