
Abstract
Rationale.
Glucose is the main metabolic fuel of the brain. The rate of glucose delivery from food to the bloodstream depends on the nature of carbohydrates in the diet, which can be summarized as the glycaemic index (GI).
Objectives.
To assess the benefit of a low versus high GI breakfast on cognitive performances within the following 4 h.
Methods.
The influence of the GI of the breakfast on verbal memory of young adults was measured throughout the morning in parallel to the assessment of blood glucose levels. The learning abilities of rats performing an operant-conditioning test 3 h after a breakfast-like meal of various GI was also examined.
Results.
A low GI rather than high GI diet improved memory in humans, especially in the late morning (150 and 210 min after breakfast). Similarly, rats displayed better learning performance 180 min after they were fed with a low rather than high GI diet.
Conclusion.
Although performances appeared to be only remotely related to blood glucose, our data provide evidence that a low GI breakfast allows better cognitive performances later in the morning.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
The ingestion of dietary carbohydrate rapidly increases the level of blood glucose. However, the rate of glucose delivery depends on physicochemical properties of the food, which can be summarized by the glycaemic index (GI), a parameter that reflects the rate and extent of the postprandial rise and fall in blood glucose (Jenkins 1981). A low GI food leads to a smaller rise of longer duration in postprandial glycaemia than a high GI one. Tables of the GI of hundreds of food items are now available (Foster-Powell and Miller 1995; Foster-Powell et al. 2002). In vitro analysis has distinguished fractions of carbohydrates into slowly available glucose (SAG) and rapidly available glucose (RAG) (Englyst et al. 1999, 2000). The RAG fraction is rapidly absorbed and largely determines the glycaemic response to a food, i.e. its GI value.
Glucose is the main metabolic fuel for the brain and is therefore necessary for mental performance. There are reports that in both animals (Gold 1991) and humans (Benton and Owens 1993), the increased provision of glucose is associated with better cognition. In the present work, we studied the influence of the RAG and SAG contents of the breakfast on mental performances in both humans (memory tests) and rats (operant conditioning test). We found that for both rats and humans, the consumption of a low GI breakfast high in SAG was associated with better cognitive performance later in the morning.
Materials and methods
Diets
In all the experiments described, four different cereal-based diets were used and presented as plain biscuit (diet 1 and diet 3), cereal bar (diet 2) or breakfast cereals (diet 4). i.e. the usual form for this type of product. As described in Table 1, the main differences between these diets originate from the kind of carbohydrates (SAG versus RAG) they contain. The GI of the diet was calculated based on its composition, using a barycentric formula (Englyst et al. 2002).
Clinical study
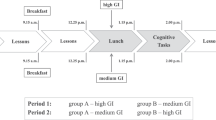
Informed consent was obtained from 106 female undergraduates, mean age 21 years and 1 month. After fasting overnight, the subjects were randomly allocated to one of two meals: 1) breakfast of diet 1 (n=36), and 2) breakfast of diet 2 (n=35). Eighty percent of the subjects were habitual breakfast eaters. However, in this as well as in previous similar studies, whether subjects normally ate breakfast or not had no influence on the results (unpublished data).
Breakfast was taken at 9.00 a.m., lasted 10 min and consisted of 50 g of one of the four diets with a sugar-free orange drink, decaffeinated coffee or tea with added skimmed milk and artificial sweetener if required. Later in the morning, the subjects drank water if desired.
Memory was assessed 30, 90, 150 and 210 min after breakfast. Four different lists of 30 words, 12 abstract and 18 concrete, matched for difficulty, were presented at a rate of one word every 2 s. Subjects wrote down as many words as they could recall immediately after presentation and again after 10 min of performing other tasks that are not reported. Concurrently, blood glucose determinations were made with a glucose oxidase assay using blood withdrawn from a fingerprick, before breakfast and after 20, 50, 80, 140, 200 and 230 min. The local Ethics Committee (University of Swansea, UK) approved the protocol.
Rat studies
In parallel to this human study, we investigated the effects of these foods on the learning abilities of rats to establish an animal model that would allow later investigation of the underlying cerebral mechanisms. All animal experiments were carried out according to the National Institute of Health Guide for the Care and Use of Laboratory Animals. In a first experiment, we focused on the effect of high SAG versus high RAG diets on the learning performances of adult Wistar male rats, 3 h after a breakfast-like meal. The diets were ground and mixed in order to obtain a homogenous pastry, which was presented as pellets to the animals. Four groups of rats (n=12 per group) were habituated to the diets and trained (during 10 days) to a breakfast-like feeding schedule. The animals were deprived of food during the light phase (light/dark 12:12). Then the rats were given a 30 min meal of 20% (i.e. between 3 g and 3.5 g of food, depending on the energy content of each diet) of their spontaneous daily energy intake, given at the beginning of the dark phase (when rats usually begin to eat) and followed by 3 h starvation until ad libitum feeding was allowed again for the remaining part of the dark phase. During the training period, the breakfast-like meal was made up of the experimental diet on odd days, whereas standard lab chow was given on even days, as well as during any other period of food availability (i.e. during nocturnal ad libitum feeding). This procedure avoided any side effects due to neophobia during the testing day. On day 11, learning performance was assessed by mean of an operant conditioning test based on the natural aversion of rats to bright light (Messaoudi et al. 1996). Three hours after breakfast, the animals were placed in a strongly illuminated (1200 lux) cage (50×40×37 cm) equipped with two levers. By pressing the active lever (AL) the rats switched off the light for 30 s, whereas the inactive lever (IL) had no effect on the environment. The test lasted 20 min. A significant difference in the number of time the rats pressed on the AL compared to the IL was interpreted as efficient learning. Differences between groups were assessed during the second half of the 20-min testing period by comparing of the AL to IL ratio (the discrimination ratio). The analysis was performed after verification that the overall number of lever pushes was not significantly different between the groups. If a rat did not press at least once on each lever during the first 10 min, it was discarded from the analyses. This happened for a maximum of three rats per group and was independent of the kind of diet. These procedures were applied under the French governmental authorization no. B-54805.
Results
Clinical study
The high RAG breakfast (diet 2) produced a marked rise in blood glucose that returned to baseline values over the course of the morning. The pattern for the high SAG breakfast (diet 1) was markedly different; the increase in blood glucose was relatively small and returned to baseline values more rapidly (Fig. 1 upper part).

Time course of the mean (±SEM) blood glucose levels recorded in humans in parallel to the cognitive tests (upper part), and results of the memory scores (mean number of words remembered±SEM) observed throughout the morning (lower part). Note that, when the effect of the diet on memory score appeared to be significant, blood glucose was no longer different. * Denotes a significant difference at P<0.05
The influence of breakfast on memory was examined using a three-way analysis of variance: Type of breakfast×Test session×Immediate/delayed recall, with the last two factors as repeated measures. As there was no interaction with immediate versus delayed recall, the results were further analysed as a global score that was the summation of immediate and delayed recall. When memory was considered (global score), there was a significant Meal×Time interaction [F(6,309)=2.12, P<0.05], reflecting a significant difference in memory 150 min [F(2,103)=3.11, P<0.05] and 210 min after breakfast [F(2,103)=5.83, P<0.004] but not after 30 and 90 min (Fig. 1, lower part).
The influence of the type of breakfast on memory was further examined by considering the ability to recall concrete words, that are easier to recall, and abstract words, that are more difficult to remember. When recall of concrete words was considered, the effect of meal was non-significant [F(2,103)=2.26, NS], although the Meal×Test session interaction approached significance [F(6,309)=2.03, P<0.06]. In fact those consuming the high SAG breakfast recalled more concrete words at 210 min [high SAG: 11±0.9 versus high RAG: 8.1±0.5 words F(2,103)=4.05, P<0.02], although not earlier. In contrast when recall of abstract words was considered there was a significant effect of the type of breakfast, the consumption of a high SAG breakfast was associated with better memory throughout the morning, the greatest difference being observed at 210 min [high SAG: 4.5±0.5 versus high RAG: 3±0.3 words, F(2,103)=3.48, P<0.03]. Thus the consumption of a high-SAG breakfast was associated with better memory throughout the morning, more particularly for the difficult abstract words.
Rat studies
Rats fed with the high SAG content diets significantly discriminated the AL from the IL (diet 1 and diet 3: P<0.05, diet 2 and diet 4: NS, two tailed Student's t-test). Eating of the high-SAG meal was associated with a significantly better discrimination ratio (Fig. 2, part 1, and Table2). The learning performance of adult rats was significantly better 3 h after a high-SAG rather than high-RAG breakfast of equal energy content. The same experiment was repeated, with a new group of animals, with diet 3 and diet 4, the only difference being that the breakfast-like meal provided 35% (i.e. between 5.5 g and 6 g of food depending on the energy content of each diet) rather than 20% of the daily energy intake of the rats. The results obtained were similar, i.e. only the high-SAG breakfast (diet 3) allowed efficient learning, and discrimination was significantly superior to that with the high-RAG breakfast (Fig. 2, part 2). Since there was no significant difference due to the caloric intake, it seems that the quality of the carbohydrates contained in the diet is of greater importance than the quantity of energy ingested.

Discrimination recorded in rats submitted to a light extinction test 3 h after a breakfast-like meal of whether 20% (part 1) or 35% (part 2) of their daily energy intake. Data are presented as mean±SEM of the discrimination ratio between the active (AL) and the inactive lever (IL) during the second half of the test period. High-SAG diets (diet 1 and diet 3) allowed better performance whatever the size of the breakfast. * Denotes a significant difference at P<0.05 (unpaired two-tailed Student's t-test)
We also tested how much the improvement of learning performance was resistant to an increased energy demand. To this end, we used two groups of rats fed with a high SAG diet (diet 3) in the same conditions than in experiment 1 (habituation to diet, breakfast-like schedule training). On the day of testing, animals had their breakfast-like meal and were then deprived of food until the end of the light extinction test that occurred 3 h after breakfast. During the 3-h deprivation period, one of the two groups of rats was submitted to a physical workload that began 140 min after the breakfast-like meal and lasted for 10 min. The rats then had 30 min rest before to be submitted to the light extinction test. The workload consisted of 10 min forced swimming in a circular 30-cm diameter water tank, which represents a significant, but not exhausting, exercise that increases the utilization of carbohydrates (Benthem et al. 1994). Comparison of the discrimination ratio did not show any significant difference due to presence or absence of workload (discrimination ratio with workload: 2.3±0.7, without workload: 1.9±0.2, NS). Thus, the beneficial effects of high SAG breakfast on later learning performances in rats appeared to be resistant to increased energy consumption (swimming) between breakfast and the learning test.
Discussion
Previous studies indicate that better memory is associated with breakfast rather than fasting in both human adults (Benton and Sargent 1992; Geisler and Polich 1992; Smith et al. 1994; Benton and Parker 1998) and rodents (Messier and Destrade 1988). Our study supports evidence that if energy and macronutrients content are similar, a high-SAG diet improves memory performance in human adults from 150 to 210 min after breakfast, but not earlier. In fact, the main result was that memory performance was maintained in the subjects who had a high-SAG breakfast, whereas with the high-RAG diet they declined progressively throughout the morning.
The results obtained with memory tests in humans were corroborated by the better learning observed in rats with the high-SAG diets. Interestingly, the better performances of high-SAG fed rats were maintained even though these animals were previously submitted to a physical workload, i.e. to increased carbohydrate utilisation, between breakfast and testing. Similarly, we did not find any significant difference in learning performances due to the size of the breakfast, i.e. to the gross quantity of carbohydrates ingested (20 versus 35% of the daily energy intake).
In fact, quantitative variations of the inflow (meal size) and outflow (workload or not) of carbohydrates did not affect performance. Moreover, in the human study, when memory performance appeared to be significantly different depending on the diet, blood glucose level had already returned to baseline. Thus any direct and simultaneous relationship between blood glucose level and performance was not apparent.
Nevertheless, in both the human and rat studies, the nature of the carbohydrates was associated with differences in cognitive performance. If blood glucose is not the underlying mechanism, other consequences of the postprandial metabolic state need to be explored.
Taken together, these results clearly suggest that the nature of the carbohydrates and the subsequent metabolic processes are important, although the mechanism is unclear. In particular, regarding memory in humans and learning in rats, there is an advantage for high-SAG, low GI as compared to high-RAG, high GI diets.
The involvement of general metabolism, and not only the level of blood glucose, may partly explain the fact that some, but not all, cognitive tests are directly sensitive to the provision of glucose (Donohoe and Benton 1999). A role for the general metabolic management of energy substrates, rather than only blood glucose, is supported by the demonstration that not only carbohydrates but also pure-fat or pure-protein meals improved memory in humans (Kaplan et al. 2001). It seems that energy intake as such may modulate cognitive performance.
The release of several types of peptides in the postprandial metabolic state offers a range of possible mechanisms. Cholecystokinine (Flood and Morley 1989), gastrin releasing peptide (Morley et al. 1992), amylin (Flood and Morley 1992) or insulin (Park et al. 2000) have all been shown to modulate cognitive performance.
In particular, there is a growing body of evidence that insulin has a key role in the relationship between glucose flux and mental performance, mainly via impaired glucose tolerance. For example, diabetes mellitus has a significant negative impact on cognition (Gispen and Biessels 2000). On the other hand, several neurodegenerative diseases such as Alzheimer's and Huntington's diseases have been found to be associated with impaired glucose regulation (for review, see Park 2001). More recently, a relationship between glucose regulation and cognitive performance was described in subjects with normal glucose tolerance (Kaplan et al. 2002).
Low GI diets are known to generate a smoother profile of postprandial insulin release than high GI diets (for review, see Ludwig 2002). Lower insulinaemia should result in less glucose being directed towards insulin-sensitive tissues such as muscles, liver or white adipose tissue. Consequently, the flow of glucose from the blood to the brain may be favoured, allowing a faster local energy recruitment when the cognitive demand occurs.
Although the underlying mechanisms are still a matter of debate, it appears that the qualitative characteristics of a diet can benefit cognitive performance. In particular we evidenced that 1) the consumption of a high-SAG, low GI breakfast is advantageous as compared to high-RAG, low GI one, 2) this phenomenon was observable throughout the morning and 3) the rat is an interesting model for future studies, especially invasive ones. As further investigations will determine which metabolic processes are involved, we will be able to understand better how, and how much, food can improve mental functioning for both healthy and diseased individuals.
References
Benthem L, Bolhuis JW, van der LJ, Steffens AB, Zock JP, Zijlstra WG (1994) Methods for measurement of energy expenditure and substrate concentrations in swimming rats. Physiol Behav 56:151–159
Benton D, Owens DS (1993) Blood glucose and human memory. Psychopharmacology 113:83–88
Benton D, Parker PY (1998) Breakfast, blood glucose, and cognition. Am J Clin Nutr 67:772S–778S
Benton D, Sargent J (1992) Breakfast, blood glucose and memory. Biol Psychol 33:207–210
Donohoe RT, Benton D (1999) Cognitive functioning is susceptible to the level of blood glucose. Psychopharmacology 145:378–385
Englyst KN, Englyst HN, Hudson GJ, Cole TJ, Cummings JH (1999) Rapidly available glucose in foods: an in vitro measurement that reflects the glycemic response. Am J Clin Nutr 69:448–454
Englyst KN, Hudson GJ, Englyst HN (2000) Encyclopaedia of analytical chemistry. Wiley, New York, pp 4246–4262
Englyst KN, Vinoy S, Englyst HN, Lang V (2002) Rapidly and slowly available glucose: physicochemical measures that describe the glycaemic index of cereal products. Br J Nutr (in press)
Flood JF, Morley JE (1989) Cholecystokinin receptors mediate enhanced memory retention produced by feeding and gastrointestinal peptides. Peptides 10:809–813
Flood JF, Morley JE (1992) Differential effects of amylin on memory processing using peripheral and central routes of administration. Peptides 13:577–580
Foster-Powell K, Hoft SHA, Brand-Miller JC (2002) International tables of glycemic index and glycemic load values: 2002. Am J Clin Nutr 76:5–56
Foster-Powell K, Miller JB (1995) International tables of glycemic index. Am J Clin Nutr 62:871S–890S
Gispen WH, Biessels G-J (2000) Cognition and synaptic plasticity in diabetes mellitus. Trends Neurosci 23:542–549
Geisler MW, Polich J (1992) P300, food consumption, and memory performance. Psychophysiology 29:76–85
Gold PE (1991) An integrated memory regulation system: from blood to brain. In: Frederickson RCA, McGaugh JL, Felton DL (eds) Peripheral signaling of the brain. Hogrefe and Huber, New York, pp 391–419
Jenkins DJ, Wolever TM, Taylor RH, Barker H, Fielden H, Baldwin JM, Bowling AC, Newman HC, Jenkins AL, Goff DV (1981) Glycemic index of foods: a physiological basis for carbohydrates exchange. Am J Clin Nutr 34:362–366
Kaplan RJ, Greenwood CE, Winocur G, Wolever TM (2001) Dietary protein, carbohydrate, and fat enhance memory performance in the healthy elderly. Am J Clin Nutr 74:687–693
Kaplan RJ, Greenwood CE, Winocur G, Wolever TM (2002) Cognitive performance is associated with glucose regulation in healthy elderly persons and can be enhanced with glucose and carbohydrates. Am J Clin Nutr 72:825–836
Ludwig DS (2002) The glycemic index physiological mechanisms relating to obesity, diabetes, and cardiovascular disease. JAMA 287:2414–2423
Messaoudi M, Tricoire A, Lalonde R, Canini F, Minn A (1996) Effects of MPTP on lever-pressing for light extinction in rats. Eur J Pharmacol 299:17–20
Messier C, Destrade C (1988) Improvement of memory for an operant response by post-training glucose in mice. Behav Brain Res 31:185–191
Morley JE, Flood J, Silver AJ (1992) Effects of peripheral hormones on memory and ingestive behaviors. Psychoneuroendocrinology 17:391–399
Park CR (2001) Cognitive effects of insulin in the central nervous system. Neurosci Biobehav Rev 25:311–323
Park CR, Seeley RJ, Craft S, Woods SC (2000) Intracerebroventricular insulin enhances memory in a passive-avoidance task. Physiol Behav 68:509–514
Smith A, Kendrick A, Maben A, Salmon J (1994) Effects of breakfast and caffeine on cognitive performance, mood and cardiovascular functioning. Appetite 22:39–55
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Benton, D., Ruffin, MP., Lassel, T. et al. The delivery rate of dietary carbohydrates affects cognitive performance in both rats and humans. Psychopharmacology 166, 86–90 (2003). https://doi.org/10.1007/s00213-002-1334-5
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00213-002-1334-5