
Abstract
Since extracts from the plants Rhois aromatica and Solidaginis virgaurea are being used in the phytotherapy of bladder dysfunction including the overactive bladder syndrome, and since muscarinic receptors are the main pharmacological target in the treatment of bladder dysfunction, we have investigated whether these extracts can inhibit carbachol-induced, muscarinic receptor-mediated contraction of rat and human bladder. In vitro contraction experiments were performed with rat and human bladder strips. Radioligand binding and inositol phosphate accumulation studies were done with cells transfected with human M2 or M3 muscarinic receptors. Both extracts concentration-dependently (final concentrations 0.01–0.1%) inhibited carbachol-induced contraction of rat and human bladder with insurmountable antagonism. Radioligand binding experiments and inositol phosphate accumulation studies with cloned receptors demonstrated direct but non-competitive effects on muscarinic receptors. Reductions of KCl-induced bladder contraction demonstrated that inhibition by the higher extract concentrations also involved receptor-independent effects. We conclude that extracts from Rhois aromatica and Solidaginis virgaurea inhibit muscarinic receptor-mediated contraction of rat and human bladder. While this could contribute to the beneficial effects of these extracts in patients with bladder dysfunction, such therapeutic effects remain to be demonstrated in controlled clinical studies.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Symptoms of bladder dysfunction are highly prevalent (Goepel et al. 2002; Milsom et al. 2001), markedly impair the quality of life of the afflicted patients and place a large socio-economic burden on society (Wyndaele 2001). The overactive bladder syndrome, the most frequent cause of bladder dysfunction, is characterized by urgency with and without urge incontinence, usually accompanied by urinary frequency and nocturia (Abrams et al. 2002). These symptoms are thought to result from involuntary bladder contractions during the filling phase of the micturition cycle (Wyndaele 2001). Muscarinic acetylcholine receptors are the predominant receptor system controlling bladder contractility (Andersson 1993). Therefore, muscarinic receptor antagonists such as oxybutynin or tolterodine are the mainstay of medical treatment for overactive bladder, but their use can be limited by side effects such as dry mouth (Chapple 2000; Herbison et al. 2003).
Many patients and physicians like to use phytotherapy to alleviate bladder dysfunction, and in some European countries certain plant extracts are fully reimbursable prescription medications. Solidaginis virgaurea (European golden rod), which is indigenous to middle Europe, is the plant that is most frequently extracted to yield preparations for the treatment of bladder dysfunction including the overactive bladder syndrome, but some medications additionally contain extracts from Rhois aromatica (sweet sumach bark), which is indigenous to Northern America (Schilcher et al. 1989). Despite the popularity of plant extracts in the treatment of urological disease, little is known about their clinical efficacy, their active ingredients or the mechanisms which might underlie their clinical effects. Rhois and Solidaginis extracts contain numerous constituents including several flavanoids and sterols, each of which can have multiple biological effects (Schilcher et al. 1989). Previous studies with the extracts and/or chemical entities isolated from them have shown diuretic, anti-inflammatory and anti-bacterial effects in vitro and/or in animals in vivo (Leuschner 1995; Schilcher et al. 1989). Against this background, we have tested whether Rhois and Solidaginis extracts inhibit the contraction of rat and human bladder to carbachol and KCl in vitro and which mechanisms might underlie such inhibition.
Materials and methods
Chemicals
Aqueous extracts of root cortex of Rhois aromatica and of leaves of Solidaginis virgaurea were obtained from Organotherapeutische Werke GmbH (Ettlingen, Germany). They had been prepared according to standardized proprietary Good Manufacturing Practice procedures for the extract-containing German prescription medication Inconturina SR. For the Solidago extract this involved aqueous extraction, evaporation of the eluate on spissum extract and dilution with water. For the Rhois extract a similar procedure was used except that 30% ethanol in water was used instead of water for extraction and dilution. For our experiments the aqueous solutions were diluted in the respective buffers. Carbachol HCl was purchased from Sigma-Aldrich (Taufkirchen, Germany). [3H]QNB ([3H]-l-quinuclidinylbenzylate, specific activity 48 Ci/mmol) and myo-[3H]inositol (specific activity 74 Ci/mmol, pre-purified with PT6–271) were obtained from Amersham (Braunschweig, Germany).
Tissue and receptor sources
Human bladder samples were obtained from patients of either gender undergoing radical cystectomy due to bladder cancer; all samples were from macroscopically tumour-free areas and were obtained with informed written consent following approval of the study protocol by the institutional ethical committee. Adult female Wistar rats were obtained from the breeding facility at the University of Essen. HEK 293 cells, which had been stable transfected with the human M2 and M3 receptors (Schmidt et al. 1995), were obtained from Dr. van Koppen at the Department of Pharmacology of the University of Essen.
Bladder contraction experiments
Contraction experiments were performed in Krebs-Henseleit solution as described using human bladder muscle strips of approximately 9–13 mm length and 1–2 mm diameter (Fetscher et al. 2002) or whole rat urinary bladders divided into four horizontal strips of approximately 12–20 mm length and 1 mm diameter (Kories et al. 2003). Cumulative carbachol concentration-response curves were constructed in the absence and then in the presence of up to three plant extract concentrations; responses in the presence of extract were normalized to % of the maximum response in its absence within the same muscle strip. In some experiments, KCl (50 mM) rather than a carbachol was used to elicit bladder contraction.
Radioligand binding studies
[3H]QNB saturation and competition binding experiments were performed in a total volume of 1,000 μl of binding buffer (10 mM Na2HPO4, 10 mM NaH2PO4 at pH 7.4) as previously described for porcine and human bladder (Goepel et al. 1998). Non-specific binding was defined as binding in the presence of 3 μM atropine. Six radioligand concentrations (approximately 10–800 pM, depending on the absence or presence of extract) were tested in the saturation binding experiments. In competition binding experiments a single radioligand concentration (approximately 300 pM) and 21 narrowly spaced concentrations of competitors were used.
Inositol phosphate measurements
Phospholipase C activation assessed as inositol phosphate formation in myo-[3H]inositol (2 μCi/ml) labelled M3 receptor-transfected cells was determined as previously described for α1-adrenoceptors (Taguchi et al. 1998). Aliquots of the cell suspension (approximately 106 cells) were incubated in a total volume of 1 ml of medium containing 20 mM HEPES and 50 mM LiCl at pH 7.4 with or without 10 μM carbachol and the indicated extract concentrations for 45 min at 37°C. Reactions were stopped by addition of 1 ml ice-cold methanol, and then 1.5 ml chloroform and 0.5 ml water were added. Following vigorous vortexing the phases were separated by centrifugation, and aliquots of the upper phase were placed on Dowex AG 1-X8 columns to elute total inositol phosphates. All measurements were done in triplicates in each experiment.
Data analysis
Data are means ± SEM of n experiments. Concentration-response curves were analysed by fitting of sigmoidal functions to the pooled experimental data using the Prism program (Graphpad Software, Sand Diego, CA, USA). The statistical significance of differences between groups was determined in a one-way ANOVA followed by Dunnett’s multiple comparison test with P<0.05 being considered as significant.
Results
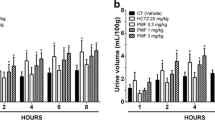
Carbachol contracted rat bladder strips with a −log EC50 of 5.79±0.03 and maximum effects of 45±2 mN (n=38). Both extracts concentration-dependently inhibited the contraction by reducing maximum responses with little changes of agonist potency; statistically significant inhibition was already seen at extract concentrations of 0.01% (Fig. 1, Table 1). In a second series of experiments, we have directly compared the effects of both extracts alone and in combination with that of time controls. In these experiments the combination reduced maximum carbachol responses significantly more than either extract alone in rats but not in humans (Fig. 2).

Effects of plant extracts on carbachol-induced contraction of rat bladder strips. Data are means ± SEM of 5–6 experiments and expressed as % of the maximum carbachol response within the same bladder strip prior to extract addition. A quantitative analysis of the data is provided in Table 1

Effects of plant extracts alone and in combination on maximum carbachol-induced contraction of rat and human bladder strips. Data are means ± SEM of 6 and 7 experiments for rat (top panel) and human bladder (bottom panel), respectively, and expressed as % inhibition relative to maximum effects within the same bladder strips in the first carbachol concentration-response curve, i.e., prior to extract or vehicle addition. *P<0.05, **P<0.01, and ***P<0.001, vs. control (time control without extract addition) in a one-way ANOVA followed by Dunnett’s multiple comparison test. ++ P<0.01 vs. combination
To determine whether inhibition of carbachol-induced contraction involved receptor-independent effects, experiments were performed with 50 mM KCl in rat bladder strips which caused an average contraction of 16.6±1.7 mN (n=28). Addition of either extracts or their combination significantly inhibited KCl-induced bladder contraction (Fig. 3), but this reached statistical significance only for the high concentration of 0.1%.

Effects of plant extracts on KCl-induced contraction of rat bladder strips. Data are means ± SEM of 7 experiments and expressed as % of the KCl response within the same bladder strip prior to extract addition. *P<0.05 and ***P<0.001, vs. control in a one-way ANOVA followed by Dunnett’s multiple comparison test
To determine whether the extracts can directly act on muscarinic receptors, radioligand binding studies were performed with cells expressing the human M2 or M3 receptor. Both extracts inhibited [3H]QNB binding in concentrations compatible with their inhibition of carbachol-induced bladder contraction (Fig. 4). To test whether this involved competitive or non-competitive inhibition, [3H]QNB saturation experiments were performed in the absence and presence of the extracts. These demonstrated a reduction of the apparent Bmax of the radioligand, which was accompanied by a statistically significant increase of the apparent Kd of [3H]QNB in the presence of Solidaginis but not of Rhois extract at concentrations compatible with inhibition of carbachol-stimulated bladder contraction (Fig. 5, Table 2). Higher concentrations of the Rhois extract paradoxically enhanced [3H]QNB binding (Fig. 4); this was due to a marked enhancement of non-specific binding despite a complete inhibition of specific binding (data not shown). Thus, both extracts can act directly on muscarinic receptors to cause non-competitive inhibition; moreover, high concentrations of the Rhois extract enhance non-specific radioligand binding.

Effects of plant extracts on [3H]QNB competition binding to rat bladder and cloned human M2 and M3 receptors. Data are means ± SEM of 4–5 experiments and expressed as % of specific radioligand binding in the absence of extract. Note that all extracts were tested in concentrations up to 1%, but concentrations higher than those shown here paradoxically increased radioligand binding (data not shown)

Effects of plant extracts on saturation binding isotherms to human M3 receptors. Data are from a representative experiment with each value determined in duplicate and are shown as total binding (TB), non-specific binding (NSB), and specific binding (SB) in the absence (control) and presence of 0.01% Rhois and 0.05% Solidaginis extract. A quantitative analysis of all such experiments is shown in Table 2
To test whether the direct extract effects on the muscarinic receptors cause functional antagonism, a possible inhibition of carbachol-stimulated inositol phosphate formation was studied in M3 receptor expressing cells. Carbachol (10 μM, corresponding to approximately 70-fold its EC50) enhanced inositol phosphate formation by 140±24% over basal (n=5). This response was inhibited almost completely by 0.1% of either extract (Fig. 6).

Effects of plant extracts on carbachol-stimulated inositol phosphate formation in cells stable transfected with human M3 receptors. Data are means ± SEM of 3–5 experiments and expressed as % above basal inositol phosphate formation
Finally, we have tested the effects of the Rhois and Solidaginis extracts on human bladder strips, which were contracted by carbachol with a −log EC50 of 6.01±0.05 and maximum effects of 35±4 mN (n=27). Both extracts inhibited carbachol-induced contraction, and this was largely due to diminished maximum carbachol responses (Fig. 2 bottom). On the other hand, the extracts had little effect on agonist potency (−log EC50 carbachol in the presence of Rhois 0.01% 5.92±0.11, 0.03% 5.81±0.09, 0.1% 5.56±0.11; −log EC50 carbachol in the presence of Solidaginis 0.01% 5.90±0.07, 0.03% 5.61±0.14, 0.1% 5.47±0.18). In this series of experiments, the combined effect of both extracts was not more effective than either extract alone (Fig. 2 bottom).
Discussion
Bladder tissue of humans, rats, rabbits, guinea pigs and pigs contains M2 and M3 muscarinic receptors in an approximately 75–80:20–25% ratio, but the contractile response to the exogenous muscarinic agonist carbachol and to endogenous agonist released by field stimulation occurs predominantly if not exclusively via M3 receptors in rats, mice, pigs, dogs and humans (Fetscher et al. 2002; Hegde and Eglen 1999). Inhibition of muscarinic receptors underlies the therapeutic effects of established, chemically-defined agents for the treatment of overactive bladder such as oxybutynin or tolterodine (Chapple 2000). Extracts from Solidaginis and, to a lesser extent, from Rhois have been used for a long time to treat bladder dysfunction but their potential mechanism of action remains unclear (Schilcher et al. 1989). Our experiments demonstrate for the first time that extracts from both Rhois and Solidaginis can concentration-dependently inhibit the muscarinic receptor-mediated contraction of rat bladder. Since it is well established that the overall voiding physiology in humans differs considerably from that in rats, it appears even more important that both extracts also inhibited muscarinic receptor-mediated contraction of human bladder.
While some chemically defined agents for the treatment of overactive bladder such as oxybutynin or tolterodine exhibit competitive, surmountable muscarinic receptor antagonism (Chapple 2000), others such as darifenacin exhibit insurmountable, at least partly non-competitive antagonism (Fetscher et al. 2002; Kories et al. 2003). Low concentrations of the Rhois and Solidaginis extracts also produced insurmountable inhibition of muscarinic receptor-mediated bladder contraction and, within the same concentration range, non-competitive inhibition of radioligand binding to muscarinic receptors. Moreover, the extracts also inhibited muscarinic receptor-stimulated inositol phosphate formation, which is compatible with their inhibitory effects on bladder contraction. Thus, the experiments with cloned receptors clearly demonstrate direct muscarinic antagonism of both extracts at the receptor level. In this regard the Rhois extract appeared to be slightly more potent, but neither extract exhibited relevant selectivity for M2 relative to M3 receptors.
While these data clearly demonstrate direct muscarinic receptor antagonism by both extracts, their functional effects appear to be complex as to be expected from plant extracts, which contain several ingredients (Schilcher et al. 1989). Thus, high concentrations (particularly of the Rhois extract) affected non-specific radioligand binding. Moreover, high concentrations of both extract also inhibited receptor-independent bladder contraction. Similarly, a Solidaginis extract has been reported to inhibit acetylcholine-, histamine- and BaCl2-induced contraction in guinea pig ileum (Leuschner 1995). Although this has not been addressed directly within our study, the receptor-independent effects of the extracts make it likely that they will also antagonise bladder contractions induced via other receptor systems such as purinergic, serotonin, bradykinin or protease-activated receptors (Kubota et al. 2003; Nakahara et al. 2003). Moreover, the presence of such receptor-independent extract effects may also explain why bladder contraction was slightly more sensitive to inhibition than inositol phosphate formation.
In summary our study demonstrates that extracts from both Rhois and Solidaginis inhibit muscarinic receptor-mediated contraction of rat and human bladder. Inhibition by low extract concentrations appears to result from non-competitive muscarinic receptor antagonism, whereas higher concentrations can also affect receptor-independent bladder contraction and may have non-specific effects. The relationship of either concentration in vitro to therapeutically used extract dosages remains unclear since the active ingredients of the extracts and their bioavailability are unknown. If tissue concentrations of active ingredients causing muscarinic receptor antagonism are reached in vivo, this could contribute to beneficial clinical effects in patients with overactive bladder. At least in rats the in vitro inhibition by combined administration of both extracts exceeded that obtained with either extract alone, which could provide a rationale for the combined use of both extracts. Whether the isolated and/or combined use of Rhois and Solidaginis extracts is indeed useful in the treatment of bladder dysfunction remains to be studied clinically. A placebo-controlled, double-blind study of these extracts is currently under way in patients with overactive bladder.
References
Abrams P, Cardozo L, Fall M, Griffiths D, Rosier P, Ulmsten U, van Kerrebroeck P, Victor A, Wein A (2002) The standardisation of terminology of lower urinary tract function: report from the standardisation sub-committee of the International Continence Society. Neurourol Urodyn 21:167–178
Andersson K-E (1993) Pharmacology of lower urinary tract smooth muscles and penile erectile tissues. Pharmacol Rev 45:253–308
Chapple CR (2000) Muscarinic receptor antagonists in the treatment of overactive bladder. Urology 55 [Suppl 5A]:33–46
Fetscher C, Fleichman M, Schmidt M, Krege S, Michel MC (2002) M3 muscarinic receptors mediate contraction of human urinary bladder. Br J Pharmacol 136:641–643
Goepel M, Gronewald A, Krege S, Michel MC (1998) Muscarinic receptor subtypes in porcine detrusor: comparison with humans and regulation by bladder augmentation. Urol Res 26:149–154
Goepel M, Hoffmann J, Piro M, Rübben H, Michel MC (2002) Prevalence and physician awareness of symptoms of urinary bladder dysfunction. Eur Urol 41:234–239
Hegde SS, Eglen RM (1999) Muscarinic receptor subtypes modulating smooth muscle contractility in the urinary bladder. Life Sci 64:419–428
Herbison P, Hay-Smith J, Ellis G, Moore K (2003) Effectiveness of anticholinergic drugs compared with placebo in the treatment of overactive bladder: systematic review. BMJ 326:841–844
Kories C, Czyborra C, Fetscher C, Schneider T, Krege S, Michel MC (2003) Gender comparison of muscarinic receptor expression and function in rat and human urinary bladder: differential regulation of M2 and M3? Naunyn-Schmiedebergs Arch Pharmacol 367:524–531
Kubota Y, Nakahara T, Mitani A, Maruko T, Saito M, Sakamoto K, Ishii K (2003) Possible involvement of Ca2+-independent phospholipase A2 in protease-activated receptor-2-mediated contraction of rat urinary bladder. Naunyn-Schmiedebergs Arch Pharmacol 367:588–591
Leuschner J (1995) Anti-inflammatory, spasmolytic and diuretic effects of a commercially available Solidago gigantea herb extract. Arzneimittelforschung 45:165–168
Milsom I, Abrams P, Cardozo L, Roberts RG, Thüroff JW, Wein AJ (2001) How widespread are the symptoms of an overactive bladder and how are they managed? A population-based prevalence study. BJU Int 87:760–766
Nakahara T, Kubota Y, Mitani A, Maruko T, Sakamoto K, Ishii K (2003) Protease-activated receptor-2-mediated contraction in the rat urinary bladder: the role of urinary bladder mucosa. Naunyn-Schmiedebergs Arch Pharmacol 367:211–213
Schilcher H, Boesel R, Effenberger S, Segebrecht S (1989) Neuere Untersuchungsergebnisse mit aquaretisch, antibakteriell und prostatatrop wirksamen Arzneipflanzen. Urologe B 29:267–271
Schmidt M, Bienek C, van Koppen CJ, Michel MC, Jakobs KH (1995) Differential calcium signalling by m2 and m3 muscarinic acetylcholine receptors in a single cell type. Naunyn-Schmiedebergs Arch Pharmacol 352:469–476
Taguchi K, Yang M, Goepel M, Michel MC (1998) Comparison of human α1-adrenoceptor subtype coupling to protein kinase C activation and related signalling pathways. Naunyn-Schmiedebergs Arch Pharmacol 358:100–110
Wyndaele JJ (2001) The overactive bladder. BJU Int 88:135–140
Acknowledgments
This work was supported in part by grants from Organotherapeutische Werke GmbH (Ettlingen, Germany) and the Deutsche Forschungsgemeinschaft (Mi 294/7–1). VB was recipient of a thesis fellowship of the intramural grant program of the University of Essen Medical School (IFORES). We thank Annette Kötting for her skilful technical help with the inositol phosphate experiments.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Borchert, V.E., Czyborra, P., Fetscher, C. et al. Extracts from Rhois aromatica and Solidaginis virgaurea inhibit rat and human bladder contraction. Naunyn-Schmiedeberg's Arch Pharmacol 369, 281–286 (2004). https://doi.org/10.1007/s00210-004-0869-x
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00210-004-0869-x