
Abstract
Intercellular communications within the cancer microenvironment coordinate the assembly of various cell types. Exosomes are mediators of intercellular communication in immune signaling, tumor promotion, stress responses, and angiogenesis. The present research aimed to determine whether miRNAs secreted from human bronchial epithelial (HBE) cells transformed by 1.0 μM arsenite are transferred into normal HBE cells and are functionally active in the recipient cells. The results show that miR-21 is involved in exosome-mediated intercellular communication between neoplastic and normal HBE cells. Exosomes derived from transformed HBE cells stimulated proliferation of normal HBE cells, whereas exosomes from miR-21 depleted cells failed to stimulate proliferation. In normal HBE cells, the expression of phosphatase and tensin homolog, a target gene for miR-21, was increased by exosomal miR-21, indicating that exogenous miRNAs, via exosomal transport, function-like endogenous miRNAs. Concordantly, specific reduction of miR-21 content in exosome-producing transformed cells abolished the stimulation of proliferation by exosomes. Collectively, the data indicate that transformed HBE cells release exosomes containing miR-21, stimulating proliferation in neighboring normal HBE cells and supporting the concept that exosomal miRNAs are involved in cell–cell communication during carcinogenesis induced by environmental chemicals.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
The development of human cancers is a multistep process in which normal cells acquire characteristics that ultimately lead to their conversion into cancer cells (Hu and Polyak 2008). Considerable interest in the cancer field is focused on the characteristics of the tumor microenvironment and on intercellular communication of malignant and nonmalignant cells (Kucharzewska et al. 2013). Intercellular communications within the cancer microenvironment coordinate the assembly of various cell types for the form and function of cancers (Calvo and Sahai 2011; Gerdes and Pepperkok 2013). Transformed cells exchange signals with surrounding fibroblasts, endothelial cells, and immune cells through direct cell–cell interactions and through secreted molecules (Hale et al. 2012).
Exosomes are small membrane vesicles with an endosome origin that are released by cells into the extracellular environment. They transfer a cargo of proteins, lipids, and nucleic acids to recipient cells, thereby altering the biochemical composition, signaling pathways, and gene regulation of recipient cells (Ge et al. 2012; Simona et al. 2013). Exosome-mediated signaling promotes tumor progression through communication between the tumor and surrounding stromal tissue (Lin et al. 2013), activation of proliferative and angiogenic pathways (Tadokoro et al. 2013; Yang et al. 2013), bestowing immune suppression (Zhang and Grizzle 2011), and initiation of pre-metastatic sites (Bissell and Hines 2011).
MicroRNAs (miRNAs), which are small, noncoding RNA molecules of 21–23 nucleotides, are involved in the regulation of biological processes, including cell proliferation, differentiation, apoptosis, homeostasis, and stress responses (DeCastro et al. 2013; Ling et al. 2012). The biological significance of miRNAs secreted outside of cells is beginning to be recognized (Liang et al. 2013). Via exosomes, cells secrete miRNAs into the extracelluar environment, where they may be involved in the propagation of cancer cells (Cortez et al. 2011; Huang et al. 2013). We hypothesized that malignant effusions have a specific, cell-free miRNA profile that conveys information regarding the presence of cancer cells. Exosomal miRNAs, however, have not been considered to be involved in the transformation of normal cells to malignant cells induced by environmental carcinogens.
Ours and other groups have reported that expression patterns of aberrant miRNAs and genetic changes contribute to environmental carcinogen-induced malignant transformation and subsequent tumor formation (Han et al. 2013; Luo et al. 2013; Tellez et al. 2011). Arsenite, an environmental carcinogen, up-regulates expression of cellular miR-21, which is involved in malignant transformation of cells (Ling et al. 2012). Although the mechanism by which miR-21 is involved in arsenite-induced cell transformation has been elucidated, it has not been determined if miR-21-mediated cell-to-cell communication contributes in arsenite-induced malignant transformation and subsequent tumor formation.
In this study, we evaluated the interaction of arsenite-transformed human bronchial epithelial (HBE) cells and normal HBE cells, using a no-contact, coculture system. The results show that miR-21, transferred from transformed HBE cells into normal HBE cells, is an extracellular signaling molecule that affects the proliferation of HBE cells, a process that can transform normal cells to malignant cells. We report, for the first time, that malignant cell-to-normal cell communication mediated by an exosomal miRNA may be involved in malignant transformation of cells and tumor formation induced by environmental carcinogens. Such information contributes to an understanding of carcinogenesis caused by arsenite.
Materials and methods
Cell culture and reagents
HBE cells, a SV40-transformed normal human bronchial epithelial cell line, are nontumorigenic and retain features of human bronchial epithelial cells. They are useful for studies of multistage bronchial epithelial carcinogenesis. These cells were obtained from the Shanghai Institute of Cell Biology, Chinese Academy of Sciences (Shanghai, China) and were maintained under 5 % CO2 at 37 °C in minimum essential medium, eagle’s medium (MEM), supplemented with 10 % fetal bovine serum (FBS, Life Technologies/Gibco, Grand Island, NY), 100 U/ml penicillin, and 100 μg/ml streptomycin (Life Technologies/Gibco, Gaithersburg, MD). We previously established the model of arsenite-transformed HBE cells (Xu et al. 2012). Cells (1 × 106) were seeded into 10-cm (diameter) dishes for 24 h and maintained in 0.0 or 1.0 μM sodium arsenite (NaAsO2, Sigma, St. Louis, MO) for 48–72 h per passage. This process was continued for about 15 weeks (30 passages). Human IL-6-neutralizing antibody (anti-IL-6 Ab, Clone 6,708), and negative control Ab (anti-IGg) were purchased from R&D Systems (Minneapolis, MN). Sodium arsenite (CAS no 7784-46-5, purity: 99 %) was purchased from Sigma. All other reagents used were of analytical grade or the highest grade available.
Coculture and preparation of culture medium
Arsenite-transformed HBE cells and HBE cells were cocultured, physically separate, using Transwell filters (polycarbonate membrane insert, 0.45-mm pore; Millipore, Billerica, MA, USA). Before coculture, HBE cells were plated onto the bottom of a 24-well plate at a density of 5 × 105 cells/well, and transformed cells and normal cells were cultured in serum-free MEM. The next day, transformed HBE cells were seeded onto the inside of an insert above the membrane at a density of 5 × 105 cells/ml, and coculture was started by setting the insert on the 24-well plate. After 24 h of coculture, the culture medium was collected and centrifuged at 2,000g for 15 min. The supernatant was filtered through a 0.22-mm polyvinylidene difluoride (PVDF) filter (Millipore) to eliminate cellular debris.
Determination of cell proliferation
Cell proliferation was evaluated by WST-8 hydrolysis using Cell Counting Kit-8 (Dojindo Molecular Technologies, Inc.) as described previously (Xu et al. 2012). Briefly, cells were seeded into 96-well tissue culture plates at 4,000 cells per well. Plates were incubated for 24 h at 37 °C with 5 % CO2 in a humidified incubator. After cells were treated and after incubation, 20 μl of WST-8 was added to each well and the incubation continued for an additional 3 h. Plates were read on a Biorad ELISA plate reader (Bio-TEK Instruments, Winooski, VT, USA) using a 450-nm filter. Results of at least three independent experiments were analyzed in duplicate. The relative cell proliferation ratios were plotted along with non-treated controls to determine the 100 % level of activity.
Growth kinetics
HBE cells were seeded in 6-well plates at a concentration of 1 × 105 per well. The plates were incubated at 37 °C under 5 % CO2 in MEM medium supplemented with 10 % FBS for 1–7 days, then collected by trypsinization. Cells were counted in triplicate using a hemocytometer under a microscope. Doubling times were calculated by the formula (Nekanti et al. 2010):
where, T D is the population doubling time expressed as hours per doubling, T is days of assay, N 0 is the number of cells at time 0, and N t is the cell number on the last day.
Quantitative real-time PCR
Total cellular RNA was isolated by use of Trizol (Invitrogen) according to the manufacturer’s recommendations. Extracelluar RNA was isolated by use of mirVana™ PARIS™ Kits (Life Technologies) according to the manufacturer’s recommendations. For detection of miR-21, 2 μg of total RNA, miRNA-specific stem-loop reverse transcriptase (RT) primers, and MMLV RT (Promega Corp., Madison, WI, USA) were used in reverse transcription following the manufacturer’s protocol. Cel-miR-39 (RiBoBio, Guangzhou, China) and U6 were used as controls for conditioned medium and cells, respectively. The sequences of mature miRNAs were from Sanger miRBase (http://microrna.sanger.ac.uk/sequences/). Forward (F) and reverse (R) primers were as follows: miR-21-F, 5′-ACACTCCAGCTGGGTAGCTTATCAGACTGA-3′; miR-21-R, 5 -TGGTGTCGTGGAGTCG-3′; U6-F, 5′-CGCTTCGGCAGCACATATACTAAAATTGGAAC-3′; U6-R, 5′-GCTTCACGAATTTGCGTGTCATCCTTGC-3′. All of the primers were synthesized by Invitrogen. Quantitative real-time PCR was performed with an Applied Biosystems 7300HT machine and MaximaTM SYBR Green/ROX qPCR Master Mix (Fermentas, USA). The concentrations of extracellular miRNAs were calculated based on their Ct values, normalized by those of cel-miR-39, which was present in each reaction mixture at 1 nM (Kosaka et al. 2010). Fold changes in expression of each gene were calculated by a comparative threshold cycle (Ct) method using the formula 2−(ΔΔCt) (Livak and Schmittgen 2001).
Exosome isolation
The culture medium was collected and centrifuged at 3,000g for 15 min, and the supernatant was filtered through a 0.22-mm PVDF filter (Millipore). An appropriate volume of exoquick exosome precipitation solution (System Biosciences) was added to the filtered culture medium and mixed well. After refrigeration for 24 h, the mixture was centrifuged at 1,500g for 30 min, and the supernatant was removed. Exosome pellets were resuspended with the appropriate volume of the serum-free medium. Exosomes from 1 × 106 cells were suspended in 150 μl of medium.
Exosome labeling
Exosomes from 1.5 × 106 cells were suspended in 180 μl of PBS with 20 μl of 1:50 diluted PKH67 (Sigma, in Diluent C). After 3 min of incubation at room temperature (RT), 3.8 ml of exosome-free medium was added to terminate the labeling reaction, and exosomes were harvested and washed twice with PBS by centrifugation (100,000g for 1 h). Exosomes were suspended in 9.6 ml of basal medium, and 250 μl was added to a sub-confluent layer of HBE cells, which were incubated for 3 h at 37 °C. Cells were washed twice with PBS, fixed with 4 % paraformaldehyde in PBS for 30 min at RT. To stain the nuclei, 4′,6-diamidino-2-phenylindole (DAPI, Sigma) was added for 10 min, and stained cells were observed under a fluorescence microscope (Zeiss, LSM700B, Germany).
Western blots
Cell lysates were prepared with a detergent buffer as described previously (Xu et al. 2012). Protein concentrations were measured according to the manufacturer’s manual using the BCA protein assay (Beyotime Institute of Biotechnology, Shanghai, China). Exosome samples were diluted 1:1 in exosome sample buffer [5 % SDS, 9 M urea, 10 mM EDTA, 120 mM Tris–HCl (pH 6.8), 2.5 % mercaptoethanol] and heated (95 °C, 5 min). Protein concentrations were measured according to the manufacturer’s manual using the Bradford protein assay (Beyotime Institute of Biotechnology). Equal amounts (80 μg) of protein were separated by 10 % sodium dodecyl sulfate–polyacrylamide gel electrophoresis and transferred to polyvinylidene fluoride membranes (Millipore, Billerica, MA, USA). Membranes were then incubated overnight at 4 °C with a 1:1,000 dilution of anti-glyceraldehyde 3-phosphate dehydrogenase (GAPDH, Sigma) and with Abs for phosphatase and tensin homolog (PTEN, Cell Signaling Technology), CD63, and flotillin-1(Abcam). After additional incubation with a 1:1,000 dilution of an anti-immunoglobin horseradish peroxidase-linked Ab for 1 h, the immune complexes were detected by enhanced chemiluminescence (Cell Signaling Technology).
Cell transfection
Cells (5 × 105) were seeded into 6-well plates at 18–24 h prior to transfection. MicroRNA-21-mimic, anti-miR-21, miR-NC-mimic, and anti-miR-nc (a negative control) were synthesized by RiBoBio (Guangzhou, China). Cells were transiently transfected using the Lipofectamine 2,000 reagent (Invitrogen, Carlsbad, CA, USA) according to the manufacturer’s protocol. After 6 h, cells were harvested and used for experiments. Transfections of HBE cells were performed with the N-TER™ Nanoparticle siRNA Transfection System (Sigma) following the manufacturer’s protocol. The siRNA nanoparticle formation solution was prepared by adding target gene siRNA dilutions to N-TER peptide dilutions. The preparations were incubated at room temperature for 30 min. NFS transfection medium (2 ml) containing target gene siRNA was transferred to each well of the culture plates, and, after for 24 h, cells were treated and harvested for analysis. Control siRNA and STAT3 siRNA were purchased from Santa Cruz Biotechnology (Santa Cruz, CA).
Statistical analyses
Derived values are presented as the mean ± SD. Comparison of mean data among multiple groups was analyzed by one-way analysis of variance (ANOVA), and a multiple-range, least significant difference was used for intergroup comparisons. P values <0.05 were considered statistically significant. All statistical analyses were performed with SPSS 16.0.
Results
Arsenite-transformed HBE cells in coculture enhance proliferation of HBE cells
Intercellular communications within the cancer microenvironment coordinate the assembly and function of multiple cell types to form a cancer (Bissell and Hines 2011). Communication between normal and abnormal cells may occur in precancerous states. Since we previously reported that the medium from arsenite-transformed HBE cells induced activation of STAT3 (Luo et al. 2013), we predicted that transformed cells promote proliferation of normal cells by contact-independent interactions. To evaluate this hypothesis, we exposed HBE and arsenite-transformed HBE cells to basal medium, medium from normal HBE cells, and medium from arsenite-transformed HBE cells. After 24 or 48 h of incubation of HBE cells, the medium from arsenite-transformed HBE cells promoted their growth by ~200 % relative to the growth of cells exposed to basal medium (Fig. 1a). The growth of cells incubated in the control medium (CM) of HBE cells was not changed. For arsenite-transformed HBE cells, the CM of HBE cells and medium from transformed HBE cells (T-CM) had no effect on proliferation (Fig. 1b). To determine the effect of no-contact interaction on cell growth, cell proliferation was tested in mono- and coculture media. Cocultured HBE cells showed a higher proliferation rate, but cocultured transformed HBE cells showed relatively less proliferation (Fig. 1c). These results indicate that cancer cells secrete molecules that effect proliferation of normal cells, and that normal cells influence proliferation of cancer cells.

In coculture, arsenite-transformed HBE cells enhance HBE cell proliferation. Basal basal medium, CM medium from normal HBE cells, T-CM medium from arsenite-transformed HBE cells, coculture arsenite-transformed HBE cells cocultured with HBE cells, HBE normal HBE cells, T-HBE arsenite-transformed HBE cells. HBE cells were exposed to basal medium, medium from normal HBE cells, or medium from arsenite-transformed HBE cells for 24 or 48 h. a Proliferation efficiency was measured by use of Cell Counting Kit-8 assay, and the relative ratios of cell proliferation were determined by comparison with cells in basal medium (mean ± SD, n = 3). *P < 0.05 difference from cells in basal medium. Transformed HBE cells were exposed to basal medium, medium from normal HBE cells, or medium from arsenite-transformed HBE cells for 24 h or 48 h. b Proliferation efficiency was measured by use of a Cell Counting Kit-8 assay, and the relative ratios of cell proliferation were determined by comparison with cells in basal medium (mean ± SD, n = 3). HBE cells and arsenite-transformed HBE cells were cocultured (no-contact) in Transwells for 24 or 48 h. c The proliferation efficiency of HBE cells and transformed cells was measured by use of a Cell Counting Kit-8 assay, and the relative ratios of cell proliferation were determined by comparison with cells in basal medium (mean ± SD, n = 3). *P < 0.05 difference from cells in basal medium
Extracellular miR-21, derived from arsenite-transformed cells, transfers into normal HBE cells
Cancer cells secrete autocrine and/or paracrine factors that regulate the functions of neighboring cells, leading to abnormal development (Kosaka et al. 2012). These factors include miRNAs, which are thought to be involved in intercellular communication (Umezu et al. 2013). We previously reported that miR-21 is up-regulated in arsenite-transformed cells (Luo et al. 2013). The intracellular and extracellular expression of miR-21 in HBE cells with or without medium from arsenite-transformed HBE cells was determined. The medium from arsenite-transformed cells induced expression of miR-21 in HBE cells (Fig. 2a), but, for arsenite-transformed cells, conditioned media did not influence the expression of miR-21 (Fig. 2b). In the supernatant, miR-21, which was preferentially expressed by transformed HBE cells, was reduced by the supernatant of transformed HBE cells cocultured with HBE cells (Fig. 2c). Moreover, the intracellular levels of miR-21 were up-regulated in HBE cells cocultured with transformed HBE cells (Fig. 2d). However, the expression of intracellular miR-21 in transformed cells did not differ with or without coculture with HBE cells (Fig. 2e), indicating that intracellular miR-21 in transformed cells was not influenced by the presence of HBE cells. These results indicate that extracellular miR-21, derived from transformed cells, transfers into HBE cells.

Extracellular miR-21, derived from arsenite-transformed cells, transfers into normal HBE cells. Basal basal medium, CM medium from normal HBE cells, T-CM medium from arsenite-transformed HBE cells, coculture arsenite-transformed HBE cells cocultured with HBE cells, HBE normal HBE cells, T-HBE arsenite-transformed HBE cells. HBE cells were exposed to basal medium, medium from normal HBE cells, or medium from arsenite-transformed HBE cells for 24 h. a levels of miR-21 were determined by qRT-PCR assays (mean ± SD, n = 3). *P < 0.05 difference from cells in basal medium. Transformed HBE cells were exposed to basal medium, medium from normal HBE cells, or medium from arsenite-transformed HBE cells for 24 h. b Levels of miR-21 were determined by qRT-PCR assays (mean ± SD, n = 3). HBE cells and arsenite-transformed HBE cells were cocultured (no-contact) in Transwells for 24 h. c The expression of miR-21 in supernatants in mono-culture medium (supernatant of HBE cells, or arsenite-transformed HBE cells only) and coculture medium (supernatant of HBE cells with arsenite-transformed HBE cells) was determined by qRT-PCR assays (mean ± SD, n = 3). *P < 0.05 difference from supernatant of arsenite-transformed HBE cells, # P < 0.05 difference from coculture medium. HBE cells and arsenite-transformed HBE cells were cocultured (no-contact) in Transwells for 24 h. d miR-21 expression in HBE cells with or without arsenite-transformed HBE cells was determined by qRT-PCR assays (mean ± SD, n = 3). *P < 0.05 difference from HBE cells without arsenite-transformed HBE cells. e miR-21 expression in arsenite-transformed HBE cells with or without HBE cells was determined by qRT-PCR assays (mean ± SD, n = 3)
Transfer of miR-21 derived from arsenite-transformed cells to HBE cells via exosomes
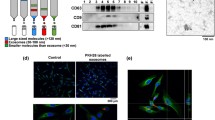
Exosomes, small membrane vesicles of endocytic origin secreted by most cancer cell types, contain miRNA and are thought to be involved in intercellular communication (Keller et al. 2006). Thus, we considered the possibility that miRNA is transferred from cancer cells into neighboring normal cells via exosomes. Exosomal fractions were prepared from the culture media of HBE cells and transformed HBE cells. Expression of the exosome markers, CD63 and flotillin-1, were assessed by immunoblotting (Keller et al. 2006; Luga et al. 2012). The expression levels of CD63 and flotillin-1 were not different in the exosomes of HBE cells and transformed HBE cells (Fig. 3a). Moreover, when HBE cells were exposed to 1.0 μM arsenite and the exosome fraction was isolated after 24 h, the expression levels of CD63 and flotillin-1 were not changed (Fig. 3b). Since the amounts of exosomes released from normal cells, transformed cells, and arsenite-treated cells did not differ, the contents of exosomes in different conditioned media were determined. The levels of miR-21 were higher in the exosomes of arsenite-treated and transformed cells (Fig. 3c, d). These results showed that, although the amounts of exosomes released from these types of cells did not differ, there were differences in exosome contents of miR-21.

Transfer of miR-21 derived from arsenite-transformed cells to HBE cells via exosomes. CM medium from normal HBE cells, T-CM medium from arsenite-transformed HBE cells. Exosomes of HBE cells and arsenite-transformed HBE cells were fractionated by Exoquick. a CD63 and flotillin-1 immunoblot of exosomes derived from HBE cells and arsenite-transformed HBE cells. HBE cells were exposed to 0.0 or 1.0 μM arsenite for 24 h, and exosomes of treated cells were fractionated. b CD63 and flotillin-1 immunoblot of exosomes derived from HBE cells and arsenite-treated HBE cells. c Real-time PCR detection of miR-21 expression in exosomes of HBE cells and arsenite-transformed HBE cells (mean ± SD, n = 3). *P < 0.05 difference from exosomes of HBE cells. d Real-time PCR detection of miR-21 expression in exosomes of HBE cells and arsenite-treated HBE cells (mean ± SD, n = 3). *P < 0.05 difference from exosomes of untreated HBE cells. The exosomes of arsenite-transformed cells were labeled with PKH67. e HBE cells after 3 h incubation of exosomes with fluorescently labeled PKH67; green represents PKH67, blue represents nuclear DNA staining by DAPI (color figure online)
The capacity of exosomes from transformed HBE cells to be transferred to recipient HBE cells was examining by assessing the uptake of isolated exosomes labeled with a green fluorescent dye, PKH67. After 3 h, allowing time for exosome uptake, a prerequisite for subsequent RNA transfer (Pegtel et al. 2010), fluorescence microscopy demonstrated that PKH67 had been taken up and transferred to perinuclear compartments, presumably representative of late endocytic compartments (Fig. 3e). In sum, extracellular miR-21 apparently transfers into HBE cells via exosomes derived from transformed HBE cells.
IL-6 up-regulates extracellular miR-21 levels in exosomes derived from HBE cells via activating STAT3 in HBE cells exposed to arsenite
We previously reported that arsenite causes up-regulation of miR-21, which is involved in the transformation of HBE cells induced by chronic exposure to arsenite, and that the medium from arsenite-transformed HBE cells induces activation of STAT3, which up-regulates miR-21 expression (Luo et al. 2013). In the present study, we determined whether IL-6 mediates miR-21 levels in exosomes via inducing STAT3 activation in cells exposed to arsenite. Anti-IL-6 Ab was used to investigate the role of IL-6 in the exosomal miR-21 levels. In the arsenite-treated HBE cells, the activation of STAT3 was reversed by decreasing the levels of IL-6 (Fig. 4a), and inhibition of IL-6 reversed the arsenite-induced increases of miR-21 expression in HBE cells (Fig. 4b). These results are consistent with our previous findings (Luo et al. 2013). In exosomes derived from HBE cells, inhibition of IL-6 also reversed the arsenite-induced increases of miR-21 expression in the supernatant of HBE cells (Fig. 4c). Transfection with STAT3 siRNA essentially abolished the arsenite-induced activation of STAT3 (Fig. 4d). Inhibition of STAT3 activation also reversed the arsenite-induced increases of miR-21 expression (Fig. 4e), an observation consistent with our previous findings (Luo et al. 2013). These results indicate that, in HBE cells, STAT3 is involved in the arsenite-induced increases of miR-21 levels. In exosomes derived from HBE cells, inhibition of STAT3 also reversed the arsenite-induced increases of miR-21 expression in the supernatant of HBE cells (Fig. 4f). These results indicate that IL-6 mediation of miR-21 levels, not only in HBE cells, but also in exosomes derived from HBE cells, is related to STAT3.

IL-6 up-regulates miR-21 levels in exosomes derived from HBE cells via activating STAT3 in HBE cells exposed to arsenite. HBE cells were cultured in the presence of anti-IL-6 Ab or anti-IgG for 3 h, then exposed to 1.0 μM arsenite for 24 h. a Western blots of p-STAT3 in HBE cells. b Levels of miR-21 measured by qRT-PCR assays in HBE cells (mean ± SD, n = 3). c Levels of miR-21 in exosomes determined by qRT-PCR assays (mean ± SD, n = 3). *P < 0.05 difference from arsenite-treated cells in the absence of anti-IL-6 Ab. HBE cells were transfected with 20 nM of control siRNA, and 20 nM STAT3 siRNA for 24 h, then they were exposed to 0.0 or 1.0 μM arsenite for 24 h. d Western blots of p-STAT3 in HBE cells. e Levels of miR-21 measured by qRT-PCR assays in HBE cells (mean ± SD, n = 3). f Levels of miR-21 determined in different treated cell-derived exosomes by qRT-PCR assays (mean ± SD, n = 3). *P < 0.05 difference from arsenite-treated cells
Regulation of the proliferation of HBE cells by cell-derived exosomes
Exosomes released by cancer cells transmit signals to other cancer cells and to stromal cells within the cancer microenvironment (Ge et al. 2012). To determine the effects of cell-derived exosomes on the regulation of proliferation of HBE and transformed HBE cells, their growth was analyzed following treatment with various concentrations of exosomes for 24 h. The results showed that exosomes derived from normal HBE cells had no effect on cell proliferation (Fig. 5a). In contrast, exosomes from arsenite-transformed HBE cells induced the growth of HBE cells in a dose-dependent manner but did not influence the growth of transformed HBE cells (Fig. 5b). Further, HBE cells were incubated with either basal medium or with different conditioned media. Proliferation, determined after 24 h of culture, increased when conditioned medium from transformed HBE cells was used. Proliferation was impaired when the conditioned medium was depleted of exosomes by ultracentrifugation. Re-addition of isolated exosomes to exosome-depleted conditioned medium, however, restored proliferation (Fig. 5c). These data demonstrate that exosomes derived from transformed HBE cells promote growth of normal HBE cells.

Effect of cell-derived exosomes on the proliferation of HBE and arsenite-transformed HBE cells. HBE normal HBE cells, T-HBE arsenite-transformed HBE cells, CM-Exosome exosomes derived from normal HBE cells, T-CM-Exosome exosomes derived from arsenite-transformed HBE cells, Basal basal medium, T-CM medium from arsenite-transformed HBE cells, T-CM-deplet T-CM-exosome-depleted conditioned medium, T-CM-re-add T-CM-exosome-depleted conditioned medium after re-addition of T-CM-exosomes. HBE and transformed HBE cells were treated with various concentrations of exosomes derived from HBE cells (0, 10, 40, 100, or 200 μg/ml) for 24 h. a Proliferation efficiency was measured by use of a Cell Counting Kit-8 assay, and the relative ratios of cell proliferation were determined by comparison with cells not exposed to exosomes (mean ± SD, n = 3). HBE and transformed HBE cells were treated with various concentrations exosomes derived from arsenite-transformed HBE cells (0, 10, 40, 100, or 200 μg/ml) for 24 h. b Proliferation efficiency was measured by use of a Cell Counting Kit-8 assay, and the relative ratios of cell proliferation were determined by comparison to cells not exposed to exosomes (mean ± SD, n = 3). *P < 0.05 difference from cells with no exosome exposure. HBE cells incubated with different conditioned media for 24 h. c Proliferation efficiency was measured by use of a Cell Counting Kit-8 assay, and the relative ratios of cell proliferation were determined by comparison with cells in basal medium (mean ± SD, n = 3). *P < 0.05 difference from cells in basal medium. # P < 0.05 difference from cells treated with medium from arsenite-transformed cells. P < 0.05 difference from cells treated with T-CM-exosome-depleted medium
Analysis of the function of exosomal miR-21 transferred into HBE cells
To establish that exosomal miR-21 is functional in the recipient cells, the expression of PTEN, one of the target genes, was evaluated in recipient HBE cells. PTEN was repressed by treatment of HBE cells with exosomal miR-21 derived from transformed HBE cells, in agreement with up-regulation of miR-21 by transfecting the miR-21 mimic (Fig. 6a). The results indicate that, in HBE cells, exogenous miR-21, incorporated by exosomal transport, functions like endogenous miR-21. To determine whether miR-21 is involved in the observed effects of exosomes, miR-21 levels were reduced by transfecting cells with anti-miR-21. As determined by qRT-PCR, transformed HBE cells with anti-miR-21 transfection produced exosomes with reduced levels of miR-21 (Fig. 6b). The expression of flotillin-1 was not changed, indicating that the amounts of exosomes released from anti-miR-NC-transfected cells and anti-miR-21-transfected cells did not differ (Fig. 6c). Nevertheless, differences in exosome content other than miR-21 cannot be excluded.

Function analysis of exosomal miR-21 transferred into HBE cells. HBE normal HBE cells, T-HBE arsenite-transformed HBE cells, Basal basal medium; CM-Exosome, exosomes derived from normal HBE cells, T-CM-Exosome exosomes derived from arsenite-transformed HBE cells. HBE cells were transfected with 100 nM miR-NC-mimic or miR-21-mimic for 24 h, or HBE cells were treated with exosomes derived from HBE or arsenite-transformed HBE cells (40 μg/ml) for 24 h. a Western blots of PTEN were made. Arsenite-transformed HBE cells were transfected with 100 nM anti-miR-NC or anti-miR-21 for 24 h. b Levels of miR-21 were determined in transformed cells and in exosomes by qRT-PCR assays (mean ± SD, n = 3). *P < 0.05 difference from cells transfected with anti-miR-NC. c Flotillin-1 immunoblots of exosomes derived from transfected, arsenite-transformed HBE cells. Arsenite-transformed HBE cells were transfected with 100 nM anti-miR-NC or anti-miR-21 for 24 h, and exosomes were fractionated by Exoquick. HBE cells were treated with exosomes derived from different conditioned media for 24 h. d Proliferation efficiency was measured by use of a Cell Counting Kit-8 assay, and the relative ratios of cell proliferation were determined by comparison to cells in basal medium (mean ± SD, n = 3). *P < 0.05 difference from cells in basal medium. # P < 0.05 difference from cells treated with T-CM-exosomes. e Growth curves for HBE cells treated with different conditions (mean ± SD, n = 3). Doubling times were calculated by the formula: T D = T × log2/(log N t − logN 0), *P < 0.05 difference from cells treated with T-CM-exosomes
Exosomes isolated from cells transfected with anti-miR-21 inhibited proliferation of HBE cells, but this effect was not evident when anti-miR-NC-transfected-exosomes were used (Fig. 6d). The doubling time of HBE cells treated with anti-miR-21-exosomes was longer than that for cells treated with anti-miR-NC-exosomes (Fig. 6d). Together, these results indicate that exosome-mediated stimulation of HBE cell proliferation is dependent on miR-21 expression in exosomes of transformed HBE cells.
Discussion
Acquisition of properties such as proliferation and resistance to programed death is not sufficient for a cell to become a tumor (Hanahan and Weinberg 2011). Cell interactions within the microenvironment are necessary in the progression from single tumor cells to a local tumor mass and to metastases (Bao et al. 2014). Transformed cells exchange signals with surrounding fibroblasts, endothelial cells, and immune cells through direct cell–cell interactions and through secreted molecules (Gerdes and Pepperkok 2013; Kosaka et al. 2010). For instance, tumors secrete growth factors for endothelial cells and also chemokines and cytokines that attract and modify the functions of immune cells (Zhang and Grizzle 2011). These results show that cancer cells have the capacity of effecting surrounding and neighboring cells by contact-independent interactions.
In coculture, Sertoli cells promote proliferation of endothelial and neural stem cells via excreting cytokines such as epithelial growth factor, nerve growth factor, and IL-6 (Zhang et al. 2012). We previously reported that transformed cells have the capacity to secrete IL-6 to promote malignant transformation of cells (Luo et al. 2013). Further, miR-143 derived from noncancerous cells suppresses cancer cell proliferation due to competition between cancer cells and non-cancer cells (Kosaka et al. 2012). Based on our previous results and those of others, we hypothesized that arsenite-transformed cells secrete molecules to regulate growth of normal cells. Oncogenesis is characterized by genetic and metabolic changes that reprogram cells to undergo uncontrolled proliferation (Hanahan and Weinberg 2011). Consistent with this concept, we also hypothesized that transformed cells induce proliferation of normal cells and affect the surrounding environment, leading to the formation of cancers.
Here, we found that, in transformed cells cocultured with normal cells, growth of the transformed cells was inhibited. We considered that an anti-proliferation molecule was secreted by normal cells to affect growth of the transformed cells. In competitive conditions, normal epithelial cells regulate the secretion of autocrine and paracrine factors that prevent aberrant growth of neighboring cells, leading to healthy development and normal metabolism, which are conducive to the maintenance of normal growth (Kosaka et al. 2010). In competitive conditions, the normal cells have only slight inhibitory effects on transformed cells.
Recently, it was recognized that RNAs serve as extracellular signals, transferring their information to recipient cells and thereby altering the biochemical composition, signaling pathways, and gene regulation of recipient cells (Cortez et al. 2011; Gerdes and Pepperkok 2013). Secreted miRNAs are considered to be involved in intercellular communication (Chen et al. 2012). Such miRNAs, transferred from cancer cells into neighboring cells, can affect the biological properties of recipient cells (Kosaka et al. 2010; Tadokoro et al. 2013).
We reported that arsenite causes up-regulation of miR-21, which is involved in the transformation of HBE cells induced by chronic exposure to arsenite, and that the medium from arsenite-transformed HBE cells induces activation of STAT3, which up-regulates miR-21 expression (Luo et al. 2013). These findings led us to consider the possibility that transformed cells secrete miR-21 for transfer to the surrounding normal cells and to promote their proliferation. The present results show that the expression of miR-21, preferentially expressed in supernatant of transformed HBE cells, was reduced in the supernatant of transformed HBE cells cocultured with HBE cells. Therefore, extracelluar miR-21 derived from transformed cells apparently transfers into normal HBE cells. Thus, in the supernatant, the miR-21 levels are reduced. Moreover, the expression of intracellular miR-21 in HBE cells was affected by coculture with transformed HBE cells, while the expression of intracellular miR-21 in transformed cells was not influenced by the presence of HBE cells. In our opinion, the extracelluar miR-21 transferred into HBE cells induces the up-regulation of intracellular miR-21. This concept is agreement with reports that Cy3-labeled miR-92a derived from K562 cells was transferred to human umbilical vein endothelial cells (HUVECs) (Umezu et al. 2013).
We previously reported that, in HBE cells, IL-6 activates STAT3 to mediate miR-21 levels. In the present study, we found that the levels of miR-21 were higher in the exosomes of arsenite-treated and transformed cells, indicating that IL-6 activates STAT3 to mediate of miR-21 levels in the exosomes of arsenite-treated cells. The results show that, with inhibition of IL-6 and inactivation of STAT3, the expression of miR-21 is reduced in exosomes derived from arsenite-treated HBE cells. Therefore, extracellular miR-21 derived from arsenite-treated cells is regulated by the IL-6/STAT3 signaling pathway. We hypothesize that arsenite-induced inflammation is involved in the intracellular and extracellular expression of miR-21.
It is now clear that, in addition to growth factors and cytokines, exosomes provide signaling for cell proliferation, differentiation, and angiogenesis (Katakowski et al. 2013; Tadokoro et al. 2013). In particular, miRNAs contained in exosomes are released from mammalian cells and act as signal transducers (Munoz et al. 2013). Endothelial cells require miR-214 to secrete exosomes that suppress senescence and induce angiogenesis in human and mouse endothelial cells (van Balkom et al. 2013). Further, THP-1 cells package miR-150 into multivesicular bodies and secrete it into the extracellular environment. Extracellular miR-150 then enters endothelial cells and enhances endothelial migration via repression of c-Myb, the target gene of miR-150 (Zhang et al. 2010). We also found that miR-21is transferred from transformed cells into normal cells via exosomes. Similar to another report (Umezu et al. 2013), our results show transfer of PKH67-labeled exosomes from transformed cells to normal cells.
Although exosomal miRNAs released from cancer cells have been extensively studied, exosomal miRNAs released from cells exposed an environmental carcinogen have not been characterized. In the present study, we evaluated the exosomal miR-21 released from arsenite-transformed cells and the exosomal miR-21 released from cells with acute exposure to arsenite. We found that the amount of exosomes did not change when cells were cultured in the presence of arsenite. Therefore, we determined the contents of these exosomes. Exosomal miR-21 was increased in cells exposed to arsenite. For cells exposed to stress, the expression of exosomal miRNAs would be consistent with the cellular expression, and this subpopulation with abnormal expression would have effects on the surrounding cells (Kosaka et al. 2012; Tadokoro et al. 2013).
Tumor exosomes, released by BT-474 human breast adenocarcinoma cells, attach to cell surfaces and increase proliferation of the releasing cells (Koga et al. 2005), and exosomes derived from SGC7901 cells promote their proliferation in a time- and dose-dependent manner (Qu et al. 2009). These results indicate that tumor-derived exosomes promote proliferation of the releasing cells and homologous tumor cells. The present results show that the exosomes from arsenite-transformed HBE cells promote proliferation of normal HBE cells in dose-dependent manner. When exposed to the different levels of exosomes derived from normal cells, there is no change in cell proliferation. Thus, in exosomes derived from normal cells, the miR-21 levels are lower than in exosomes of transformed cells. The low levels of exosomal miR-21, however, did not affect cell proliferation. To confirm the effects of exosomes on the cell proliferation, we used transformed cells, which produce exosomes containing miR-21, as a model to investigate the effects of exosomes. Exosomes from the medium of transformed cells stimulated proliferation of normal cells. Exosome-depleted medium of transformed cells reduced cell growth. Nevertheless, re-addition of exosomes to exosome-depleted medium restored the stimulatory effect, demonstrating that the procedure for exosome isolation did not affect the functional properties of exosomes. Nevertheless, not all stimulatory effects may be attributed to exosomes; growth factors and cytokines, which are also secreted by transformed cells, are also likely to contribute. Together, the present results demonstrate that exosomes derived from transformed HBE cells promote growth of normal HBE cells.
Furthermore, exosomes should not be considered to act as transfection agents, but rather as multi-component signaling devices. Exosome-derived miR-214 is functional, as the expression of the miR-214 target gene, ATM, is up-regulated upon exposure of cells to anti-miR-214 exosomes (van Balkom et al. 2013). Exosomes containing high levels of miR-210 down-regulate EFNA3, and thereby enhance tube formation of HUVECs (Tadokoro et al. 2013). PTEN is a target gene of miR-21 (Qi et al. 2009). Our results show that exosomes containing high levels of miR-21 function like endogenous miR-21 to down-regulate PTEN. In addition, by transfecting exosome-producing transformed cells with a miR-21 inhibitor, exosome miR-21 levels were reduced without affecting exosome secretion. Anti-miR-21-exosomes had a reduced capacity to stimulate proliferation. Also, miR-21 is known to affect cell proliferation (Lan et al. 2011). Our findings agree with the observed role for miR-21 in the stimulation of cell proliferation and contribute to an understanding of miR-21 biological functions.
In this research, we demonstrated that transformed cells release exosomes containing miR-21, that exosomal miR-21 is transported from arsenite-transformed cells to normal cells, and that exogenous miR-21 modulates proliferation of normal cells. We also demonstrated that exosomal miRNAs function in cell–cell communication during carcinogenesis induced by an environmental chemical. The results provide a strategy for intervention in the process of carcinogenesis.
References
Bao B, Azmi AS, Li Y et al (2014) Targeting CSCs in tumor microenvironment: the potential role of ROS-associated miRNAs in tumor aggressiveness. Curr Stem Cell Res Ther 9(1):22–35
Bissell MJ, Hines WC (2011) Why don’t we get more cancer? A proposed role of the microenvironment in restraining cancer progression. Nat Med 17(3):320–329. doi:10.1038/nm.2328
Calvo F, Sahai E (2011) Cell communication networks in cancer invasion. Curr Opin Cell Biol 23(5):621–629. doi:10.1016/j.ceb.2011.04.010
Chen X, Liang H, Zhang J, Zen K, Zhang CY (2012) Secreted microRNAs: a new form of intercellular communication. Trends Cell Biol 22(3):125–132. doi:10.1016/j.tcb.2011.12.001
Cortez MA, Bueso-Ramos C, Ferdin J, Lopez-Berestein G, Sood AK, Calin GA (2011) MicroRNAs in body fluids–the mix of hormones and biomarkers. Nat Rev Clin Oncol 8(8):467–477. doi:10.1038/nrclinonc.2011.76
DeCastro AJ, Dunphy KA, Hutchinson J et al (2013) MiR203 mediates subversion of stem cell properties during mammary epithelial differentiation via repression of DeltaNP63alpha and promotes mesenchymal-to-epithelial transition. Cell Death Dis 4:e514. doi:10.1038/cddis.2013.37
Ge R, Tan E, Sharghi-Namini S, Asada HH (2012) Exosomes in cancer microenvironment and beyond: have we overlooked these extracellular messengers? Cancer Microenviron 5(3):323–332. doi:10.1007/s12307-012-0110-2
Gerdes HH, Pepperkok R (2013) Cell-to-cell communication: current views and future perspectives. Cell Tissue Res 352(1):1–3. doi:10.1007/s00441-013-1590-1
Hale JS, Li M, Lathia JD (2012) The malignant social network: cell–cell adhesion and communication in cancer stem cells. Cell Adh Migr 6(4):346–355. doi:10.4161/cam.21294
Han Z, Zhang Y, Xu Y et al (2013) Cell cycle changes mediated by the p53/miR-34c axis are involved in the malignant transformation of human bronchial epithelial cells by benzo[a]pyrene. Toxicol Lett. doi:10.1016/j.toxlet.2013.12.008
Hanahan D, Weinberg RA (2011) Hallmarks of cancer: the next generation. Cell 144(5):646–674. doi:10.1016/j.cell.2011.02.013
Hu M, Polyak K (2008) Microenvironmental regulation of cancer development. Curr Opin Genet Dev 18(1):27–34. doi:10.1016/j.gde.2007.12.006
Huang Y, Hu Q, Deng Z, Hang Y, Wang J, Wang K (2013) MicroRNAs in body fluids as biomarkers for non-small cell lung cancer: a systematic review. Technol Cancer Res Treat. doi:10.7785/tcrt.2012.500377
Katakowski M, Buller B, Zheng X et al (2013) Exosomes from marrow stromal cells expressing miR-146b inhibit glioma growth. Cancer Lett 335(1):201–204. doi:10.1016/j.canlet.2013.02.019
Keller S, Sanderson MP, Stoeck A, Altevogt P (2006) Exosomes: from biogenesis and secretion to biological function. Immunol Lett 107(2):102–108. doi:10.1016/j.imlet.2006.09.005
Koga K, Matsumoto K, Akiyoshi T et al (2005) Purification, characterization and biological significance of tumor-derived exosomes. Anticancer Res 25(6A):3703–3707
Kosaka N, Iguchi H, Yoshioka Y, Takeshita F, Matsuki Y, Ochiya T (2010) Secretory mechanisms and intercellular transfer of microRNAs in living cells. J Biol Chem 285(23):17442–17452. doi:10.1074/jbc.M110.107821
Kosaka N, Iguchi H, Yoshioka Y, Hagiwara K, Takeshita F, Ochiya T (2012) Competitive interactions of cancer cells and normal cells via secretory microRNAs. J Biol Chem 287(2):1397–1405. doi:10.1074/jbc.M111.288662
Kucharzewska P, Christianson HC, Welch JE et al (2013) Exosomes reflect the hypoxic status of glioma cells and mediate hypoxia-dependent activation of vascular cells during tumor development. Proc Natl Acad Sci USA 110(18):7312–7317. doi:10.1073/pnas.1220998110
Lan H, Lin CY, Yuan HY, Xiong B (2011) Overexpression of miR-21 promotes proliferation and reduces apoptosis in non-small cell lung cancer. Zhonghua Zhong liu za zhi [Chinese journal of oncology] 33(10):742–746
Liang H, Gong F, Zhang S, Zhang CY, Zen K, Chen X (2013) The origin, function, and diagnostic potential of extracellular microRNAs in human body fluids. Wiley Interdiscip Rev RNA. doi:10.1002/wrna.1208
Lin R, Wang S, Zhao RC (2013) Exosomes from human adipose-derived mesenchymal stem cells promote migration through Wnt signaling pathway in a breast cancer cell model. Mol Cell Biochem 383(1–2):13–20. doi:10.1007/s11010-013-1746-z
Ling M, Li Y, Xu Y et al (2012) Regulation of miRNA-21 by reactive oxygen species-activated ERK/NF-kappaB in arsenite-induced cell transformation. Free Radic Biol Med 52(9):1508–1518. doi:10.1016/j.freeradbiomed.2012.02.020
Livak KJ, Schmittgen TD (2001) Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) method. Methods 25(4):402–408. doi:10.1006/meth.2001.1262
Luga V, Zhang L, Viloria-Petit AM et al (2012) Exosomes mediate stromal mobilization of autocrine Wnt-PCP signaling in breast cancer cell migration. Cell 151(7):1542–1556. doi:10.1016/j.cell.2012.11.024
Luo F, Xu Y, Ling M et al (2013) Arsenite evokes IL-6 secretion, autocrine regulation of STAT3 signaling, and miR-21 expression, processes involved in the EMT and malignant transformation of human bronchial epithelial cells. Toxicol Appl Pharmacol 273(1):27–34. doi:10.1016/j.taap.2013.08.025
Munoz JL, Bliss SA, Greco SJ, Ramkissoon SH, Ligon KL, Rameshwar P (2013) Delivery of functional anti-miR-9 by mesenchymal stem cell-derived exosomes to glioblastoma multiforme cells conferred chemosensitivity. Mol Ther Nucleic Acids 2:e126. doi:10.1038/mtna.2013.60
Nekanti U, Dastidar S, Venugopal P, Totey S, Ta M (2010) Increased proliferation and analysis of differential gene expression in human Wharton’s jelly-derived mesenchymal stromal cells under hypoxia. Int J Biol Sci 6(5):499–512
Pegtel DM, Cosmopoulos K, Thorley-Lawson DA et al (2010) Functional delivery of viral miRNAs via exosomes. Proc Natl Acad Sci USA 107(14):6328–6333. doi:10.1073/pnas.0914843107
Qi L, Bart J, Tan LP et al (2009) Expression of miR-21 and its targets (PTEN, PDCD4, TM1) in flat epithelial atypia of the breast in relation to ductal carcinoma in situ and invasive carcinoma. BMC Cancer 9:163. doi:10.1186/1471-2407-9-163
Qu JL, Qu XJ, Zhao MF et al (2009) Gastric cancer exosomes promote tumour cell proliferation through PI3 K/Akt and MAPK/ERK activation. Dig Liver Dis 41(12):875–880. doi:10.1016/j.dld.2009.04.006
Simona F, Laura S, Simona T, Riccardo A (2013) Contribution of proteomics to understanding the role of tumor-derived exosomes in cancer progression: state of the art and new perspectives. Proteomics 13(10–11):1581–1594. doi:10.1002/pmic.201200398
Tadokoro H, Umezu T, Ohyashiki K, Hirano T, Ohyashiki JH (2013) Exosomes derived from hypoxic leukemia cells enhance tube formation in endothelial cells. J Biol Chem 288(48):34343–34351. doi:10.1074/jbc.M113.480822
Tellez CS, Juri DE, Do K et al (2011) EMT and stem cell-like properties associated with miR-205 and miR-200 epigenetic silencing are early manifestations during carcinogen-induced transformation of human lung epithelial cells. Cancer Res 71(8):3087–3097. doi:10.1158/0008-5472.CAN-10-3035
Umezu T, Ohyashiki K, Kuroda M, Ohyashiki JH (2013) Leukemia cell to endothelial cell communication via exosomal miRNAs. Oncogene 32(22):2747–2755. doi:10.1038/onc.2012.295
van Balkom BW, de Jong OG, Smits M et al (2013) Endothelial cells require miR-214 to secrete exosomes that suppress senescence and induce angiogenesis in human and mouse endothelial cells. Blood 121(19):3997–4006, S1–S15 doi:10.1182/blood-2013-02-478925
Xu Y, Li Y, Pang Y et al (2012) Blockade of p53 by HIF-2alpha, but not HIF-1alpha, is involved in arsenite-induced malignant transformation of human bronchial epithelial cells. Arch Toxicol 86(6):947–959. doi:10.1007/s00204-012-0810-x
Yang L, Wu XH, Wang D, Luo CL, Chen LX (2013) Bladder cancer cell-derived exosomes inhibit tumor cell apoptosis and induce cell proliferation in vitro. Mol Med Rep 8(4):1272–1278. doi:10.3892/mmr.2013.1634
Zhang HG, Grizzle WE (2011) Exosomes and cancer: a newly described pathway of immune suppression. Clin Cancer Res 17(5):959–964. doi:10.1158/1078-0432.CCR-10-1489
Zhang Y, Liu D, Chen X et al (2010) Secreted monocytic miR-150 enhances targeted endothelial cell migration. Mol Cell 39(1):133–144. doi:10.1016/j.molcel.2010.06.010
Zhang F, Hong Y, Liang W, Ren T, Jing S, Lin J (2012) Co-culture with Sertoli cells promotes proliferation and migration of umbilical cord mesenchymal stem cells. Biochem Biophys Res Commun 427(1):86–90. doi:10.1016/j.bbrc.2012.09.007
Acknowledgments
The authors wish to thank Donald L. Hill (University of Alabama at Birmingham, USA) for editing. This work was supported by the Natural Science Foundations of China (30872146, 81072327, and 81273114), the Research Fund for the Doctoral Program of Higher Education of China (20103234110005), the Key Program of Educational Commission of Jiangsu Province of China (11KJA330002), and the Priority Academic Program Development of Jiangsu Higher Education Institutions (2010).
Conflict of interest
The authors listed participated in the design, execution, and analysis of these experiments. All have read the manuscript and have agreed to submit it in its current form for consideration for publication in this journal. They declare that there are no conflict of interests.
Author information
Authors and Affiliations
Corresponding author
Additional information
Yuan Xu and Fei Luo authors contributed equally.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
About this article

Cite this article
Xu, Y., Luo, F., Liu, Y. et al. Exosomal miR-21 derived from arsenite-transformed human bronchial epithelial cells promotes cell proliferation associated with arsenite carcinogenesis. Arch Toxicol 89, 1071–1082 (2015). https://doi.org/10.1007/s00204-014-1291-x
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00204-014-1291-x