
Abstract
Methanococcus maripaludis is a stringently anaerobic archaeon with two studied surface structures, archaella and type IV pili. Previously, it was shown that three pilin genes (mmp0233 [epdA], mmp0236 [epdB] and mmp0237 [epdC]) located within an 11 gene cluster in the genome were necessary for normal piliation. This study focused on analysis of the remaining genes to determine their potential involvement in piliation. Reverse transcriptase PCR experiments demonstrated the 11 genes formed a single transcriptional unit. Deletions were made in all the non-pilin genes except mmp0231. Electron microscopy revealed that all the genes in the locus except mmp0235 and mmp0238 were essential for piliation. Complementation with a plasmid-borne wild-type copy of the deleted gene restored at least some piliation. We identified genes for an assembly ATPase and two versions of the conserved pilin platform forming protein necessary for pili assembly at a separate genetic locus.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Archaea, such as Bacteria, are known to possess a variety of cell surface structures (Jarrell et al. 2013; Lassak et al. 2012a; Ng et al. 2008; Pohlschroder et al. 2011). Many of these archaeal appendages appear to be made using a system like that employed by bacteria to assemble type IV pili (Jarrell et al. 2009, 2013; Lassak et al. 2012a; Ng et al. 2008; Pohlschroder et al. 2011). The core components of such a system include structural subunits with class three signal peptides (Pohlschroder et al. 2005) which are removed by a specific signal peptidase (type IV prepilin signal peptidase), an ATPase to incorporate new subunits into the base of the growing structure (sometimes a second ATPase to remove subunits from the structure) and a conserved membrane protein that is thought to interact with the ATPase(s) as an export complex for the structural proteins (Burrows 2012; Pohlschroder et al. 2011; Takhar et al. 2013). These core components of type IV pili systems have been observed in archaea in the loci responsible for formation of archaella [formerly archaeal flagella; (Jarrell and Albers 2012)] (Ghosh and Albers 2011; Lassak et al. 2012b; Ng et al. 2006), type IV-like pili (Albers and Pohlschroder 2009; Frols et al. 2008; Henche et al. 2012a; Ng et al. 2011; Szabo et al. 2006) and the bindosome involved in substrate uptake in Sulfolobus solfataricus (Albers et al. 1999; 2007, Zolghadr et al. 2011). In addition, the Iho670 fibers of Ignicoccus hospitalis are also made from type IV pilin-like proteins (Yu et al. 2012). Presumably, there are genes that encode the other core components, but they do not appear to be located in the immediate vicinity of the structural protein gene (Muller et al. 2009). While the archaellum is the best studied of the archaeal surface structures (Ghosh and Albers 2011; Jarrell et al. 1996; Jarrell and McBride 2008, Thomas et al. 2001), more recently, various studies have targeted the structure, function and genetics of archaeal pili systems, especially ones that are type IV like (Esquivel et al. 2013; Frols et al. 2008; Henche et al. 2012a; Lassak et al. 2012a; Ng et al. 2011; Wang et al. 2008). Among the archaeal type IV-like pili, the majority of progress has been reported in Sulfolobus species (Henche et al. 2012a; Orell et al. 2013a, b; van Wolferen et al. 2013) and, to a lesser degree, in Methanococcus maripaludis (Jarrell et al. 2011; Nair et al. 2013; Ng et al. 2011; Wang et al. 2008) and Haloferax volcanii (Esquivel et al. 2013).
In Sulfolobus acidocaldarius, there are at least two different operons in the genome that are responsible for two unique pili types on the cell surface, namely Aap pili and Ups pili (Lassak et al. 2012a; Pohlschroder et al. 2011). Aap pili, observed on cells during growth under normal laboratory conditions, are involved in adhesion of cells (Henche et al. 2012a). The Ups pili, on the other hand, are observed only when the cells are subjected to DNA damaging conditions such as UV exposure (Ajon et al. 2011; Frols et al. 2008). The upregulation of the ups operon led to cell aggregation and an enhanced ability of the cells to exchange DNA (Ajon et al. 2011). A recent study showed the importance of both Ups and Aap pili on the structure of the biofilm produced by S. acidocaldarius (Henche et al. 2012b; Orell et al. 2013a). A genetic locus containing five genes (encoding two predicted pilins, a type IV pilus polymerizing ATPase, a conserved pilus membrane protein and a putative iron-sulfur oxidoreductase) has been implicated in Aap pili formation (Henche et al. 2012a). Mutants carrying deletions in any one of the five genes do not assemble Aap pili. The ups locus consists also of five genes, encoding two prepilins, a hypothetical protein as well as homologues of a type IV pilus polymerization ATPase and conserved membrane protein. No pili were observed if the gene for the ATPase or the conserved membrane protein was deleted (van Wolferen et al. 2013). Mutants carrying deletions of either of the two prepilin genes still made pili, but these cells were defective in aggregation (van Wolferen et al. 2013). Mutants carrying a deletion of the gene encoding the hypothetical protein UpsX still made pili, but decreased DNA exchange was observed (van Wolferen et al. 2013). Very recently, insights into the regulation of the pili systems of S. acidocaldarius have also been presented (Orell et al. 2013b; Vassart et al. 2012), including studies that show that there is an intertwined regulation of archaella and Aap pili formation (Henche et al. 2012a; Orell et al. 2013b; Reimann et al. 2012).
The type IV pili-like locus in M. maripaludis genome was originally predicted to consist of 11 potential genes, with three of them, mmp0233 (epdA), mmp0236 (epdB) and mmp0237 (epdC), thought to encode structural proteins (pilins) and another shown to be a type IV prepilin peptidase (mmp0232, EppA) required to cleave the class three signal peptides from the prepilins (Szabo et al. 2007). Subsequent genetic work demonstrated that all three of the genes encoding the predicted structural proteins were essential for normal piliation (Ng et al. 2011). However, mass spectrometry of purified pili revealed that the major structural pilin was MMP1685, encoded by a gene located outside the pilin locus. The deletion and complementation of mmp1685 showed that it was indeed essential for piliation (Ng et al. 2011). More recently, another minor pilin, encoded by mmp1283, was also shown to be essential for pili formation (Nair et al. 2013). An important, and thus far unique, characteristic of the pili system in M. maripaludis is the presence of a second, apparently pilin-specific peptidase [EppA, (Szabo et al. 2007)], distinct from the prepilin peptidase, FlaK, (Bardy and Jarrell 2002, 2003) needed to process archaellins for archaella assembly.
In addition to the genetic studies, the structure of M. maripaludis pili was also determined and shown to be different from that of any known bacterial pili (Wang et al. 2008). Furthermore, at least one function for the pili of M. maripaludis was demonstrated, that of surface adhesion. However, this function is dependent on the co-expression of archaella (Jarrell et al. 2011).
In this study, we complete the in-frame deletion and complementation study on the genes in the type IV pili-like locus of M. maripaludis. In addition, we identify an ATPase (mmp0040) as well as two conserved type IV pili membrane component homologues (mmp0038 and mmp0039) critical for pili assembly which are located adjacent to each other but separate from the known pili locus.
Materials and methods
Strains and growth conditions
Methanococcus maripaludis (Mm900) (Moore and Leigh 2005) and a ΔflaK mutant strain derived from Mm900 (Ng et al. 2009) were grown in Balch medium III (Balch et al. 1979) at 35 °C under a headspace of CO2/H2 (20:80). McCas medium (Moore and Leigh 2005) was used for transformation experiments with the addition of neomycin (1 mg/ml) or 8-azahypoxanthine (240 μg/ml) for selection at various steps of the procedure. Puromycin (2.5 μg/ml) was used to select for transformants carrying the complementation vectors. For complementation studies, cells were grown in nitrogen-free medium (Blank et al. 1995) supplemented with sterile anaerobic solutions of either NH4Cl (10 mM) or alanine (10 mM). Escherichia coli strain DH5α (Novagen) or E. coli TOP10 cells (Invitrogen) used for various cloning steps was grown at 37 °C in Luria–Bertani medium, with ampicillin (100 μg/ml) added for selection, when necessary.
Bioinformatic analysis
Each predicted pilus locus gene product was analyzed by a variety of online tools to gather information about its likely subcellular location and possible function. This included programs which predicted archaeal signal peptides (PRED-SIGNAL [http://bioinformatics.biol.uoa.gr/PRED-SIGNAL/ (Bagos et al. 2009)], transmembrane domains [TmPred, http://www.ch.embnet.org/software/TMPRED_form.html (Hofmann and Stoffel 1993)] and subcellular location [PSORTb version 3.0.2 trained to archaea, http://www.psort.org/psortb/index.html (Yu et al. 2010)] as well as ones that searched for conserved motifs [BLAST search Basic Local Alignment Search Tool, http://blast.ncbi.nlm.nih.gov/ (Altschul et al. 1990)] and InterProScan [http://www.ebi.ac.uk/Tools/pfa/iprscan/ (Quevillon et al. 2005)].
Plasmid construction to create gene deletions
Plasmids used for the generation of inframe deletions of pilus locus genes were generated as previously described (Moore and Leigh 2005; VanDyke et al. 2008). Briefly, P1 and P2 PCR primers for each gene (Supplemental Table 1) were selected to amplify approximately 1 kb upstream and the P3 and P4 PCR primers to amplify approximately 1 kb downstream of the targeted gene. The P2 and P3 primers were designed so that after ligation, a small internal fragment of the targeted gene was left. The gene-specific P1 and P4 primers had added BamH1 restriction sites while P2 and P3 had added Asc1 restriction sites. The upstream and downstream PCR products were ligated after digestion with Asc1, and this product was used as template for a further PCR using primers P1 and P4. This approximately 2-kb piece was digested with BamHI and cloned into pCRPrtNeo to create the plasmids used for generating the deletion strains (Moore and Leigh 2005).
M. maripaludis mutant generation
The pCRPrtNeo derivatives carrying deletions of the pilus locus genes (listed in Supplemental Table 2) were transformed into M. maripaludis ΔflaK using a PEG precipitation method as described by Tumbula et al. (Tumbula et al. 1994). After recovery overnight, the cells were transferred to McCas medium containing neomycin (1 mg/ml) to select for transformants that had integrated the vector. This culture was then used to inoculate McCas media without neomycin to allow for a second recombination event to remove the vector. At this stage, a recombination event that removes the inserted plasmid can either return the chromosome to its wild-type sequence or result in a deletion of the targeted gene.
The culture was plated onto McCas agar containing 8-azahypoxanthine (240 μg/ml), which would be lethal to any cells that retained the vector-borne hpt gene. Following incubation at 37 °C in an anaerobic canister for 1 week, individual colonies were picked and inoculated into Balch medium III for analysis. The individual transformants were screened by using washed whole cells resuspended in 2 % NaCl as template for PCR along with sequencing primers (Supplemental Table 1) designed to amplify across the target gene. The PCR products were examined by agarose gel electrophoresis and the size compared to that predicted for the wild type and deletion versions of the gene in order to identify the specific gene deletion mutants. Transformants showing the deletion size PCR products were restreaked onto Balch medium III plates and single colonies picked and again screened by PCR to confirm their purity.
Complementation of the gene deletion strains
Plasmids used for complementation of the gene deletions were constructed as previously described (Chaban et al. 2007; Lie et al. 2005) and listed in Supplemental Table 2. The vector used for complementation was the self-replicating plasmid, pHW40, a derivative of pWLG40 (Gardner and Whitman 1999), in which the transcription of the cloned gene is under the control of an inducible nif promoter (Kessler and Leigh 1999). pHW40 plasmids with the wild-type version of the complementing gene were transformed into the corresponding deletion strain using the PEG procedure (Tumbula et al. 1994). Complemented strains were grown in nitrogen-free medium supplemented with either 10 mM NH4Cl (where transcription from the nif promoter is repressed) or 10 mM alanine (where transcription from the nif promoter is on). The PCR primers used to amplify the wild-type versions of each gene for creating the complementation vectors for each gene are listed in Supplemental Table 1 (gene-specific primers labeled Comp-for and Comp-rev). NsiI and MluI restriction sites were added to the forward and reverse primers, respectively, for cloning purposes. In the case of mmp0232 complementation, site-directed mutagenesis was used to remove an internal NsiI site, using primers listed in Supplemental Table 1, prior to cloning into the complementation vector.
Reverse transcriptase PCR
Reverse transcriptase PCR (RT-PCR) experiments were performed to determine if all 11 genes in the proposed type IV pilus-like operon were co-transcribed. Primers (Supplemental Table 3) were designed such that a sequence linking two neighboring genes across the intergenic region would be amplified if the genes were co-transcribed. RNA template was extracted from wild-type cells using an RNeasy Mini Kit (Qiagen Inc. Canada Mississauga, ON) with optional DNase digestion (Qiagen Inc.) as per the manufacturer’s protocol. cDNA was amplified using a One-Step RT-PCR kit (Qiagen Inc.) in accordance with the supplied protocol. In addition to using cDNA as template, PCR reactions were done using the same primer combinations with the purified RNA without the RT step as template as a control for possible DNA contamination of the RNA samples. PCR reactions were also run with genomic DNA as template to ensure the primers amplified the predicted size fragments.
The same strategy was employed to examine the possible co-transcription of the ATPase and conserved pilus membrane component genes.
Electron microscopy
Cells were grown overnight and washed with 50 mM MgSO4 prior to treatment with 2 % phosphotungstic acid to negatively stain the samples. Cells were examined on formvar-coated gold grids and imaged under a Hitachi 7,000 electron microscope operating at an accelerating voltage of 75 kV.
Results
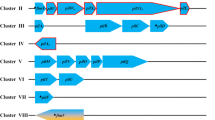
A type IV pili-like locus encoding 11 potential genes (mmp0231–mmp0241) was initially identified by Szabo et al. (2007) (Fig. 1a). We recently demonstrated the essential involvement of the three pilin-like genes (epdA, epdB and epdC) of the locus in the normal assembly of surface pili (Ng et al. 2011), while eppA was previously shown to encode the prepilin peptidase (Szabo et al. 2007). However, the possible involvement of the remaining genes in the operon in pili biosynthesis remained to be determined. These predicted proteins do not show homology to any bacterial type IV pilus genes or to pili genes in S. solfataricus. The proteins encoded by these genes were analyzed by various bioinformatics programs (PRED-SIGNAL, PSORT, TmPred and InterProScan), which at times gave conflicting predictions (Table 1). Most of the proteins are annotated as conserved hypothetical proteins and most have significant BLAST matches to only archaeal proteins and sometimes only to Methanococcus proteins. Most lack the presence of motifs that would be useful in deciphering a possible function (Table 1). Two are predicted to have signal peptides (MMP0235 and MMP0238), while MMP0241 contains a domain of unknown function (DUF2341) that is found in various bacterial proteins that form proton channels (MotA, TolQ, ExbB) or act as transport proteins. Other proteins are predicted to be cytoplasmic or membrane located (refer to Table 1).

Analysis of the major type IV pilus locus. a The major type IV pilus locus showing all 11 genes with the three pilin-like genes (epdA, epdB and epdC) and the prepilin peptidase (eppA) indicated. b RT-PCR experiment indicating co-transcription of all 11 genes. For each pair of adjacent genes, the triplet of lanes represent (1) Standard PCRs using Mm900 genomic DNA as template and the respective RT primers which amplify across the intergenic regions to indicate the expected amplicon size and primer specificity; (2) RT-PCR run using total RNA extracted from Mm900 cells as template with the same RT primers. The RT lanes which have bands at the same size as the DNA lanes indicate the co-transcription of the indicated genes; (3) Standard PCRs performed using total RNA that did not undergo reverse transcription as template to rule out possible DNA contamination of the RNA sample
RT-PCR experiments
All 11 genes in the type IV pilus locus are oriented in the same direction, and the intergenic region between each adjoining gene is very small (ranging from overlapping to 52 nucleotides), with the exception of the distance between mmp0233 and mmp0234 which is 126 nucleotides. Thus, the possibility of the entire region comprising a single operon was analyzed by RT-PCR. Agarose gel electrophoresis (Fig. 1b) shows amplified PCR products of the correct predicted length in all the lanes representing the 10 intergenic region between the 11 genes when the RNA was first reverse transcribed, suggesting that all 11 genes form a single operon. These PCR products were the same size as those obtained using genomic DNA as template. No PCR products were obtained when the RT step was omitted (Fig. 1b), indicating the RNA samples were free of contaminating genomic DNA.
Targeted internal gene deletions of pili operon genes
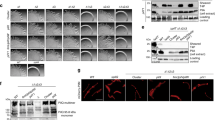
To determine whether each of the genes found in the pili locus was essential for piliation, an internal deletion of each gene was attempted (Moore and Leigh 2005; Ng et al. 2011). These deletions were all created in a flaK deletion strain of M. maripaludis. This strain lacks the signal peptidase necessary to process archaellins (Bardy and Jarrell 2002, 2003) making the cells nonarchaellated and leaving only the less numerous and thinner pili as the sole surface appendages (Ng et al. 2011). In spite of repeated attempts, an internal deletion of mmp0231 was not successful; however, deletions were created in all the remaining genes. Deletion strains were identified among the transformants by whole-cell PCR using gene-specific primers that amplified across the deleted region, resulting in a smaller PCR product in a mutant strain when compared to the products obtained with the same primer pair using the wild-type cells as template (Fig. 2). Electron microscopic examination of each mutant revealed that deletion of any of the genes, with the exception of mmp0238 and mmp0235, led to nonpiliated cells (Fig. 3). For all the deletions that led to nonpiliated cells, at least 50 cells of each mutant were examined. EM examination of the mmp0235 and mmp0238 deletion strains, on the other hand, revealed a wild-type number and appearance of pili demonstrating that neither of these genes was essential for piliation (Fig. 3). For these two mutants as well as the parent flaK deletion strain, the number of pili per cell was small (flaK 3.1 ± 2.5 pili per cell, N = 16; mmp0235 2.5 ± 1.8 pili per cell, N = 27; mmp0238 4.0 ± 2.8 pili per cell, N = 19). Even in the flaK cells, not all cells were piliated (35/39 or 83 %). All of the gene deletions that resulted in loss of piliation were complemented with plasmid-borne wild-type versions of the appropriate gene. In all cases, complementation of the deletion mutants resulted in a return to piliation (Fig. 4), although in the case of ∆mmp0232, ∆mmp0234 and ∆mmp0239, only a small percentage (4 %, N = 75) of complemented cells were piliated.

Confirmation of the deletions of the targeted pili locus genes by PCR. Washed whole cells of each deletion strain as well as wild-type cells were used as template for the PCR confirmation of each deletion, using corresponding sequencing primers (Supplemental Table 1). Results for each indicated deletion are shown in pairs with the first lane showing the PCR amplicon using wild-type cells as template and the second lane representing the amplicon from the gene deletion strain. The amplicon size in each lane is the predicted size

Electron micrographs of strains carrying the indicated pili locus gene deletion showing its effect on piliation. All deletion strains are nonpiliated except for ∆mmp0235 and ∆mmp0238. The parent strain, ∆flaK, is shown for comparison. Enlargements of a section of a cell are presented to more easily visualize the thin pili, when present. Samples were negatively stained with 2 % phosphotungstic acid (pH 7.0). Bar, 200 nm

Electron micrographs of deletion strains complemented with a plasmid-borne wild-type copy of the deleted gene. All strains were returned to a piliated state following complementation, although only a small percentage of cells were piliated in the complementations of mmp0232, mmp0234 and mmp0239. Enlargements of a section of a cell are presented to more easily visualize the thin pili. Samples were negatively stained with 2 % phosphotungstic acid (pH 7.0). Bar, 200 nm
Identification of type IV pilin ATPase and conserved membrane protein genes
Type IV pili systems in both Bacteria and archaea require at least a single ATPase to provide energy for the assembly process. Most have two ATPases, one for extension and one for retraction of the pili, enabling a type of surface motility called twitching (Burrows 2012; Mattick 2002). In addition, type IV pili systems have a conserved membrane component that acts as a platform for assembly and can interact with the ATPases (Burrows 2012; Takhar et al. 2013). The type IV pili-like locus in M. maripaludis does not contain homologues of a pilus ATPase or conserved membrane component gene. However, two potential type IV pilus ATPase genes, mmp0040 and mmp0281, were found outside the locus upon examination of the annotated sequenced M. maripaludis genome and a search for homologues to pilus ATPases from Pseudomonas aeruginosa using the BLAST algorithm. Of the two genes, mmp0040 had a higher sequence similarity to the P. aeruginosa pilus ATPases. It was also found to be in a gene cluster containing two genes encoding type IV pili-conserved membrane components (type 2 secretion system protein F, PilC in P. aeruginosa) (Fig. 5a). These three genes were shown by RT-PCR to be co-transcribed as part of a five gene operon (Fig. 5b), along with genes for transcription initiation factor B (mmp0041) and H/ACA RNA–protein complex component Gar1 (mmp0042). When the MMP0040 protein sequence was used as a query to search P. aeruginosa genomes in a BLAST search (http://blast.ncbi.nlm.nih.gov/Blast.cgi), it retrieved matches to TadA ATPases (4e-60; 61 % coverage) with less significant alignments to PilB (1e-08) and PilT (3e-08). Using the protein sequence of MMP0281 in a similar search retrieved significant alignments to trb conjugation ATPase (2e-04) and GspE (0.006) with coverage of <30 %. When the protein sequence of P. aeruginosa TadA was used as query to search the M. maripaludis genomes, it resulted in hits to MMP0040 (3e-62, 79 % coverage) as well as FlaI (3e-38, 78 % coverage) and MMP0281 (2e-07, 31 % coverage). FlaI is the ATPase for archaella assembly and rotation (Banerjee et al. 2012; Thomas et al. 2002). When the protein sequences of P. aeruginosa PilT or PilB were used as queries in a similar search, it returned significant alignments to FlaI (1e-10, 31 % coverage for PilT and 1e-10, 23 % coverage for PilB). The PilB search also identified significant alignment to MMP0040 but with low coverage (1e-07, 12 % coverage). Deletions were created in each of the two potential ATPase genes and each was examined for the presence of pili (Fig. 5c). The mmp0040 deletion strain was shown to be nonpiliated (Fig. 6), while the mmp0281 deletion strain remained piliated (Fig. 6) to a similar extent as the flaK cells (for the mmp0281 deletion strain 3.1 ± 1.9 pili per cell, N = 22). Internal deletions were created in each of the two pilus platform genes as well (Fig. 5c), and each of these deletion strains was examined for the presence of pili. Interestingly, both mmp0038 and mmp0039 were required for piliation (Fig. 6). While both MMP0038 and MMP0039 are type II secretion system protein F domain proteins, the amino acid similarity between the two proteins is very low. Complementation of the mmp0038, mmp0039 and mmp0040 deletion strains all resulted in a return to the piliated state, confirming their involvement in piliation Fig. 6).

The five gene locus containing genes for the pili assembly ATPase and two copies of the platform (pilC-like) protein. a Organization of the genetic locus. b RT-PCR experiment indicating co-transcription of all five genes. For each pair of adjacent genes, the triplet of lanes represent (1) Standard PCRs using Mm900 genomic DNA as template and the respective RT primers which amplify across the intergenic regions to indicate the expected amplicon size and primer specificity; (2) RT-PCR run using total RNA extracted from Mm900 cells as template with the same RT primers. The RT lanes which have bands at the same size as the DNA lanes indicate the co-transcription of the indicated genes; (3) Standard PCRs performed using total RNA that did not undergo reverse transcription as template to rule out possible DNA contamination of the RNA sample. (c) Confirmation of in-frame deletions of the targeted pili locus genes by PCR. Washed whole cells of each deletion strain as well as wild-type cells were used as template for the PCR confirmation of each in-frame deletion, using corresponding sequencing primers (Supplemental Table 1). Results for each indicated deletion are shown in pairs with the first lane showing the PCR amplicon using wild-type cells as template and the second lane representing the amplicon from the gene deletion strain. In all cases, the obtained PCR products were of the predicted size

Electron micrographs of strains deleted for the type IV pili ATPase and platform protein genes and their complemented strains. All deletion strains were nonpiliated, except ∆mmp0281, but were returned to a piliated state following complementation. In the case of mmp0040, the complementation returned cells to a wild-type level of piliation, while for the complementations of both mmp0038 and mmp0039, only a small percentage of cells were piliated. Samples were negatively stained with 2 % phosphotungstic acid (pH 7.0). Bar, 200 nm
A summary of the effects of all the studied gene deletions and complementations on piliation, including our previous results on the pilus structural genes (Ng et al. 2011), is presented in Table 2.
Discussion
The 11 gene type IV pili-like locus (mmp00231–mmp0241) includes a gene for a prepilin peptidase EppA (mmp0232) and three type IV pilin-like genes (mmp0233 [epdA], mmp0236 [epdB] and mmp0237 [epdC]) (Szabo et al. 2007). However, the possible involvement of the remaining genes in pili formation has not been previously addressed and was a major focus of this work.
The results of RT-PCR experiments demonstrated that all 11 genes formed a single transcriptional unit. With the exception of the prepilin peptidase and the pilins, the seven other genes in the locus represent novel genes necessary for assembly of type IV pili-like structures; they bear no homology to known type IV pili genes in bacteria or to ones so far identified in other archaea such as the Aap and Ups pili of S. solfataricus and S. acidocaldarius (Frols et al. 2008; Henche et al. 2012a). Some of the genes appear to be unique to Methanococcus and encode proteins predicted to be located in the cytoplasm, cytoplasmic membrane or to be secreted. Elucidation of the functions of these gene products will be a future challenge. Even in well studied bacterial type IV pili systems, the functions of many conserved gene products remain a mystery with the type IVa and type IVb (along with the tad/flp subgrouping), all having unique components with unknown functions (Burrows 2012). It could be speculated that all the proteins encoded within this operon may be needed in relatively small numbers and so found in an operon separate from the gene encoding the major pilin subunit, MMP1685. The latter would be needed in significantly higher numbers than the minor pilins EpdA, EpdB and EpdC and may be transcribed from a much stronger promoter. This may be how M. maripaludis obtains the optimal amounts of the different structural proteins present in significantly different stoichiometries in the final assembled pili.
Electron microscopic analysis of cells containing deletions in each of seven other genes of this major pilus operon indicated that all of the pilus locus genes, except mmp0235 and mmp0238, were essential for piliation. Successful complementation of all gene deletions indicated that the effects on piliation observed were due to the specific deleted gene and not from polar effects generated in the creation of each gene deletion. Not all complementations, however, returned the mutants to wild-type levels of piliation. In some examples, the complementation only restored piliation to a small percentage of cells. In the P. aeruginosa type IV system (Giltner et al. 2010), overexpression of minor pilins can be detrimental to pilus biosynthesis, and it has been suggested that the overabundant minor pilins may titrate out other key chromosomally encoded pilus proteins. In the cases of complementations that did not return mutant cells to a wild-type state of piliation in our experiments, it is possible that gene products were produced at above-normal levels, and this may have interfered with normal pilus assembly.
The two genes in the locus not found to be necessary for pili formation, namely mmp0235 and mmp0238, might still have a role to play in pili function, such as attachment. Adhesins have been identified in certain bacterial type IV pili systems, including Neisseria species. Here, PilC1 has been identified as an adhesin located at the tip of the pili (Rudel et al. 1995). PilC1 is made initially with a signal peptide (Morand et al. 2001), and pili can be formed without the adhesin present (Rudel et al. 1995). Both MMP0235 and MMP0238 are also made with a predicted signal peptide, and if either of these two proteins is the adhesin, then their absence may affect pili function but not its assembly.
Of the nonpilin genes studied in the major pilus locus, only mmp0232 (eppA) has an assigned function as encoding the prepilin peptidase, required for processing of all the pilin-like proteins prior to their incorporation into the pilus structure (Szabo et al. 2007). As expected, deletion of this gene led to nonpiliated cells. Among the studied archaea, Methanococcus appears unique in possessing two prepilin peptidases involved in assembly of two different surface appendages. FlaK was the first identified prepilin peptidase in archaea, and its essential role in the processing of pre-archaellins for archaella biosynthesis in Methanococcus sp. has been well established (Bardy and Jarrell 2002, 2003). Clearly, FlaK cannot compensate for the loss of EppA, and each enzyme is restricted in its substrates to either pilins or archaellins despite the similarities in the signal peptide and N-termini of the two substrates (Szabo et al. 2007). Other archaea, such as S. solfataricus and H. volcanii, possess a variety of proteins with class three signal peptides (including archaellins and pilins) but these all appear to be processed by a single enzyme, designated PibD, that possesses broad substrate specificity, (Albers and Pohlschroder 2009; Lassak et al. 2012a; Tripepi et al. 2010). Interestingly, S. solfataricus PibD was shown to be able to process M. voltae archaellin in in vitro assays (Ng et al. 2009).
Examination of the 11 gene pili cluster indicated that genes encoding conserved type IV pili ATPases and the membrane component were missing. In most type IV pili systems, an ATPase (PilB) is needed for incorporation of new subunits into the structure for pilus extension, while a separate ATPase (PilT) removes subunits from the base of the structure leading to retraction of the pilus (Burrows 2012, 2005). This extension and retraction of the pilus leads to the movement of cells across a solid surface in a process called twitching (Burrows 2012). The conserved membrane protein appears to interact with the ATPases to form a platform for assembly/disassembly of the pilus (Burrows 2012; Crowther et al. 2004). Archaeal species have not been shown to twitch, and in archaea, only homologues to PilB have been identified (Peabody et al. 2003). In S. acidocaldarius, at least two different type IV pili, produced under different growth conditions, are already known. A single ATPase and a single homologue of the conserved membrane protein have been identified in both Ups and Aap pili systems in a locus that also contains genes for pilins. Deletion of either the ATPase or the conserved membrane protein gene in either pilus system led to the inability of those mutants to assemble that particular pilus type (Henche et al. 2012a; van Wolferen et al. 2013). Deletion analysis identified a small locus in the M. maripaludis genome containing a single pilus ATPase homologue adjacent to two homologues of the conserved pilus membrane protein that were all essential for piliation. Based on this evidence, M. maripaludis pili are predicted to be unable to retract unless MMP0040 is able to perform both polymerization and removal of subunits from the structure, an idea already considered for the single ATPase Sulfolobus pili systems (Albers and Pohlschroder 2009). The presence of two divergent copies of the conserved membrane component gene is relatively rare in bacterial type IV pili systems, but found in the tad pili systems of P. aeruginosa and Aggregatibacter (previously Actinobacillus) actinomycetemcomitans (Burrows 2012). In the latter case, both membrane component genes (tadB and tadC) are required for piliation (Kachlany et al. 2000). Why two versions of this protein are required for in the Tad pili system and here in M. maripaludis is not known but it has been suggested that perhaps the single ATPase of the Tad system may interact with one version of the conserved membrane proteins in pilin addition and with the other in pilin removal (Burrows 2012). In P. aeruginosa, recent evidence led to speculation that the platform protein PilC is likely a dimer which might reside within the lumen of the hexameric ATPase (Takhar et al. 2013). The polymerization and retraction ATPases might then interact with the two cytoplasmic domains of PilC with the PilB ATPase interacting with the N-terminal domain and PilT ATPase with the C-terminal domain (Takhar et al. 2013). In systems with two platform proteins as in M. maripaludis, it is possible that a heterodimer is formed and the single ATPase interacts with one member of the dimer for extension but with the other for retraction (Burrows 2012).
The results presented in this report, coupled to our earlier investigations (Nair et al. 2013; Ng et al. 2011), indicate that genes responsible for the type IV-like pili in M. maripaludis are spread around the genome, in sharp contrast to pili loci in Sulfolobus (Lassak et al. 2012a). The 11 gene operon investigated here contains three minor pilins, the prepilin peptidase necessary for prepilin processing as well as numerous genes that have no obvious counterpart in either bacterial or other archaeal pili systems. The gene for the major structural protein MMP1685 (Ng et al. 2011) is located at an entirely different genetic locus as is the gene for an additional minor pilin MMP1283 (Nair et al. 2013), and as shown here, the genes encoding the highly conserved assembly ATPase and membrane proteins are located at a fourth distinct locus. With the involvement of five structural proteins (EpdA, EpdB, EpdC, MMP1283 and MMP1685) already determined to be necessary for normal piliation, the structure of pili of M. maripaludis seems to be more complex than Sulfolobus systems where only two pilins have been reported (Frols et al. 2008; Henche et al. 2012a; van Wolferen et al. 2013). The pili of M. maripaludis are usually observed in small numbers under routine growth conditions and, unlike in Sulfolobus (Orell et al. 2013b; Reimann et al. 2012; Vassart et al. 2012), no studies on their possible regulation have been reported. The initial publication on the 11 gene pilus locus designated mmp0233 as eppA to denote a novel subclass of a euryarchaeal type IV prepilin peptidase (Szabo et al. 2007). The pilin genes were designated epdA (mmp0233), epdB (mmp0236) and epdC (mmp0237) since they were dependent on EppA for signal peptide removal and hence EppA-dependent proteins. Since these are the only known pili in M. maripaludis and the structures are currently lacking a specific designation, we feel they could rightly be called Epd pili since the pili themselves are EppA-dependent. Since we have identified a number of genes essential for Epd pili formation in this and prior publications (Nair et al. 2013; Ng et al. 2011), we propose all of these genes now be given epd designations. For the pilins MMP1283 and MMP1685, they are given the designations EpdD and EpdE, respectively. The other genes whose deletions led to nonpiliated cells are designated as follows: mmp0234:epdF; mmp0239: epdG; mmp0240: epdH; mmp0241:epdI; mmp0038: epdJ; mmp0039: epdK and mmp0040:epdL. Delineating the roles of the novel gene products involved in M. maripaludis pili biosynthesis represent immediate challenges for the field.
References
Ajon M, Fröls S, van Wolferen M, Stoecker K, Teichmann D, Driessen AJ, Grogan DW, Albers SV, Schleper C (2011) UV-inducible DNA exchange in hyperthermophilic archaea mediated by type IV pili. Mol Microbiol 82:807–817
Albers SV, Pohlschroder M (2009) Diversity of archaeal type IV pilin-like structures. Extremophiles 13:403–410
Albers SV, Elferink MG, Charlebois RL, Sensen CW, Driessen AJM, Konings WN (1999) Glucose transport in the extremely thermoacidophilic Sulfolobus solfataricus involves a high-affinity membrane-integrated binding protein. J Bacteriol 181:4285–4291
Altschul SF, Gish W, Miller W, Myers EW, Lipman DJ (1990) Basic local alignment search tool. J Mol Biol 215:403–410
Bagos PG, Tsirigos KD, Plessas SK, Liakopoulos TD, Hamodrakas SJ (2009) Prediction of signal peptides in archaea. Protein Eng Des Sel 22:27–35
Balch WE, Fox GE, Magrum LJ, Woese CR, Wolfe RS (1979) Methanogens: reevaluation of a unique biological group. Microbiol Rev 43:260–296
Banerjee A, Ghosh A, Mills DJ, Kahnt J, Vonck J, Albers SV (2012) FlaX, a unique component of the crenarchaeal archaellum, forms oligomeric ring-shaped structures and interacts with the motor ATPase FlaI. J Biol Chem 287:43322–43330
Bardy SL, Jarrell KF (2002) FlaK of the archaeon Methanococcus maripaludis possesses preflagellin peptidase activity. FEMS Microbiol Lett 208:53–59
Bardy SL, Jarrell KF (2003) Cleavage of preflagellins by an aspartic acid signal peptidase is essential for flagellation in the archaeon Methanococcus voltae. Mol Microbiol 50:1339–1347
Blank CE, Kessler PS, Leigh JA (1995) Genetics in methanogens: transposon insertion mutagenesis of a Methanococcus maripaludis nifH gene. J Bacteriol 177:5773–5777
Burrows LL (2005) Weapons of mass retraction. Mol Microbiol 57:878–888
Burrows LL (2012) Pseudomonas aeruginosa twitching motility: type IV pili in action. Annu Rev Microbiol 66:493–520
Chaban B, Ng SY, Kanbe M, Saltzman I, Nimmo G, Aizawa SI, Jarrell KF (2007) Systematic deletion analyses of the fla genes in the flagella operon identify several genes essential for proper assembly and function of flagella in the archaeon, Methanococcus maripaludis. Mol Microbiol 66:596–609
Crowther LJ, Anantha RP, Donnenberg MS (2004) The inner membrane subassembly of the enteropathogenic Escherichia coli bundle-forming pilus machine. Mol Microbiol 52:67–79
Esquivel RN, Xu R, Pohlschroder M (2013) Novel archaeal adhesion pilins with a conserved N terminus. J Bacteriol 17:3808–3818
Frols S, Ajon M, Wagner M, Teichmann D, Zolghadr B, Folea M, Boekema EJ, Driessen AJ, Schleper C, Albers SV (2008) UV-inducible cellular aggregation of the hyperthermophilic archaeon Sulfolobus solfataricus is mediated by pili formation. Mol Microbiol 70:938–952
Gardner WL, Whitman WB (1999) Expression vectors for Methanococcus maripaludis: overexpression of acetohydroxyacid synthase and beta-galactosidase. Genetics 152:1439–1447
Ghosh A, Albers SV (2011) Assembly and function of the archaeal flagellum. Biochem Soc Trans 39:64–69
Giltner CL, Habash M, Burrows LL (2010) Pseudomonas aeruginosa minor pilins are incorporated into type IV pili. J Mol Biol 398:444–461
Henche AL, Ghosh A, Yu X, Jeske T, Egelman E, Albers SV (2012a) Structure and function of the adhesive type IV pilus of Sulfolobus acidocaldarius. Environ Microbiol 14:3188–3202
Henche AL, Koerdt A, Ghosh A, Albers S- (2012b) Influence of cell surface structures on crenarchaeal biofilm formation using a thermostable green fluorescent protein. Environ Microbiol 14:779–793
Hofmann K, Stoffel W (1993) TMbase—a database of membrane spanning proteins segments. Biol Chem Hoppe Seyler 374:166
Jarrell KF, Albers SV (2012) The archaellum: an old motility structure with a new name. Trends Microbiol 20:307–312
Jarrell KF, McBride MJ (2008) The surprisingly diverse ways that prokaryotes move. Nat Rev Microbiol 6:466–476
Jarrell KF, Bayley DP, Kostyukova AS (1996) The archaeal flagellum: a unique motility structure. J Bacteriol 178:5057–5064
Jarrell KF, VanDyke DJ, Wu J (2009) Archaeal flagella and pili. In: Jarrell KF (ed) Pili and flagella: Current research and future trends. Caister Academic Press, Norfolk, pp 215–234
Jarrell KF, Stark M, Nair DB, Chong JPJ (2011) Flagella and pili are both necessary for efficient attachment of Methanococcus maripaludis to surfaces. FEMS Microbiol Lett 319:44–50
Jarrell KF, Ding Y, Nair DB, Siu S (2013) Surface appendages of Archaea: structure, function, genetics and assembly. Life 3:86–117
Kachlany SC, Planet PJ, Bhattacharjee MK, Kollia E, DeSalle R, Fine DH, Figurski DH (2000) Nonspecific adherence by Actinobacillus actinomycetemcomitans requires genes widespread in bacteria and archaea. J Bacteriol 182:6169–6176
Kessler PS, Leigh JA (1999) Genetics of nitrogen regulation in Methanococcus maripaludis. Genetics 152:1343–1351
Lassak K, Ghosh A, Albers SV (2012a) Diversity, assembly and regulation of archaeal type IV pili-like and non-type-IV pili-like surface structures. Res Microbiol 163:630–644
Lassak K, Neiner T, Ghosh A, Klingl A, Wirth R, Albers SV (2012b) Molecular analysis of the crenarchaeal flagellum. Mol Microbiol 83:110–124
Lie TJ, Wood GE, Leigh JA (2005) Regulation of nif expression in Methanococcus maripaludis: roles of the euryarchaeal repressor NrpR, 2-oxoglutarate, and two operators. J Biol Chem 280:5236–5241
Mattick JS (2002) Type IV pili and twitching motility. Annu Rev Microbiol 56:289–314
Moore BC, Leigh JA (2005) Markerless mutagenesis in Methanococcus maripaludis demonstrates roles for alanine dehydrogenase, alanine racemase, and alanine permease. J Bacteriol 187:972–979
Morand PC, Tattevin P, Eugene E, Beretti JL, Nassif X (2001) The adhesive property of the type IV pilus-associated component PilC1 of pathogenic Neisseria is supported by the conformational structure of the N-terminal part of the molecule. Mol Microbiol 40:846–856
Muller DW, Meyer C, Gurster S, Kuper U, Huber H, Rachel R, Wanner G, Wirth R, Bellack A (2009) The Iho670 fibers of Ignicoccus hospitalis: a new type of archaeal cell surface appendage. J Bacteriol 191:6465–6468
Nair DB, Chung DKC, Schneider J, Uchida K, Aizawa SI, Jarrell KF (2013) Identification of an additional minor pilin essential for piliation in the archaeon Methanococcus maripaludis. PLoS One 8:e83961
Ng SY, Chaban B, Jarrell KF (2006) Archaeal flagella, bacterial flagella and type IV pili: a comparison of genes and posttranslational modifications. J Mol Microbiol Biotechnol 11:167–191
Ng SYM, Zolghadr B, Driessen AJM, Albers SV, Jarrell KF (2008) Cell surface structures of archaea. J Bacteriol 190:6039–6047
Ng SY, VanDyke DJ, Chaban B, Wu J, Nosaka Y, Aizawa S, Jarrell KF (2009) Different minimal signal peptide lengths recognized by the archaeal prepilin-like peptidases FlaK and PibD. J Bacteriol 191:6732–6740
Ng SYM, Wu J, Nair DB, Logan SM, Robotham A, Tessier L, Kelly JF, Uchida K, Aizawa S-, Jarrell KF (2011) Genetic and mass spectrometry analysis of the unusual type IV-like pili of the archaeon Methanococcus maripaludis. J Bacteriol 193:804–814
Orell A, Fröls S, Albers SV (2013a) Archaeal biofilms: the great unexplored. Annu Rev Microbiol 67:337–354
Orell A, Peeters E, Vassen V, Jachlewski S, Schalles S, Siebers B, Albers SV (2013b) Lrs14 transcriptional regulators influence biofilm formation and cell motility of Crenarchaea. ISME J 7:1886–1898
Peabody CR, Chung YJ, Yen MR, Vidal-Ingigliardi D, Pugsley AP, Saier MH Jr (2003) Type II protein secretion and its relationship to bacterial type IV pili and archaeal flagella. Microbiology 149:3051–3072
Pohlschroder M, Gimenez MI, Jarrell KF (2005) Protein transport in Archaea: sec and twin arginine translocation pathways. Curr Opin Microbiol 8:713–719
Pohlschroder M, Ghosh A, Tripepi M, Albers SV (2011) Archaeal type IV pilus-like structures-evolutionarily conserved prokaryotic surface organelles. Curr Opin Microbiol 14:1–7
Quevillon E, Silventoinen V, Pillai S, Harte N, Mulder N, Apweiler R, Lopez R (2005) InterProScan: protein domains identifier. Nucleic Acids Res 33:W116–W120
Reimann J, Lassak K, Khadouma S, Ettema TJ, Yang N, Driessen AJ, Klingl A, Albers SV (2012) Regulation of archaella expression by the FHA and von Willebrand domain-containing proteins ArnA and ArnB in Sulfolobus acidocaldarius. Mol Microbiol 86:24–36
Rudel T, Scheurerpflug I, Meyer TF (1995) Neisseria PilC protein identified as type-4 pilus tip-located adhesin. Nature 373:357–359
Szabo Z, Albers SV, Driessen AJM (2006) Active-site residues in the type IV prepilin peptidase homologue PibD from the archaeon Sulfolobus solfataricus. J Bacteriol 188:1437–1443
Szabo Z, Stahl AO, Albers SV, Kissinger JC, Driessen AJM, Pohlschroder M (2007) Identification of diverse archaeal proteins with class III signal peptides cleaved by distinct archaeal prepilin peptidases. J Bacteriol 189:772–778
Takhar HK, Kemp K, Kim M, Howell PL, Burrows LL (2013) The platform protein is essential for type IV pilus biogenesis. J Biol Chem 288:9721–9728
Thomas NA, Bardy SL, Jarrell KF (2001) The archaeal flagellum: a different kind of prokaryotic motility structure. FEMS Microbiol Rev 25:147–174
Thomas NA, Mueller S, Klein A, Jarrell KF (2002) Mutants in flaI and flaJ of the archaeon Methanococcus voltae are deficient in flagellum assembly. Mol Microbiol 46:879–887
Tripepi M, Imam S, Pohlschroder M (2010) Haloferax volcanii flagella are required for motility but are not involved in PibD-dependent surface adhesion. J Bacteriol 192:3093–3102
Tumbula DL, Makula RA, Whitman WB (1994) Transformation of Methanococcus maripaludis and identification of a PstI-like restriction system. FEMS Microbiol Lett 121:309–314
van Wolferen M, Ajon M, Driessen AJ, Albers SV (2013) Molecular analysis of the UV-inducible pili operon from Sulfolobus acidocaldarius. Microbiol Open 2:928–937
VanDyke DJ, Wu J, Ng SY, Kanbe M, Chaban B, Aizawa SI, Jarrell KF (2008) Identification of putative acetyltransferase gene, mmp0350, which affects proper assembly of both flagella and pili in the archaeon Methanococcus maripaludis. J Bacteriol 190:5300–5307
Vassart A, Van Wolferen M, Orell A, Hong Y, Peeters E, Albers SV, Charlier D (2012) Sa-Lrp from Sulfolobus acidocaldarius is a versatile, glutamine-responsive, and architectural transcriptional regulator. Microbiol Open 2:75–93
Wang YA, Yu X, Ng SYM, Jarrell KF, Egelman EH (2008) The structure of an archaeal pilus. J Mol Biol 381:456–466
Yu NY, Wagner JR, Laird MR, Melli G, Rey S, Lo R, Dao P, Sahinalp SC, Ester M, Foster LJ, Brinkman FS (2010) PSORTb 3.0: improved protein subcellular localization prediction with refined localization subcategories and predictive capabilities for all prokaryotes. Bioinformatics 26:1608–1615
Yu X, Goforth C, Meyer C, Rachel R, Schröder GF, Egelman EH (2012) Filaments from Ignicoccus hospitalis show diversity of packing in proteins containing N-terminal type IV pilin helices. J Mol Biol 422:274–281
Zolghadr B, Weber S, Szabo Z, Driessen AJM, Albers SV (2007) Identification of a system required for the functional surface localization of sugar binding proteins with class III signal peptides in Sulfolobus solfataricus. Mol Microbiol 64:795–806
Zolghadr B, Klingl A, Rachel R, Driessen AJ, Albers SV (2011) The bindosome is a structural component of the Sulfolobus solfataricus cell envelope. Extremophiles 15:235–244
Acknowledgments
This research was funded by a Discovery Grant from the Natural Sciences and Engineering Research Council of Canada (NSERC) to KFJ.
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by Erko Stackebrandt.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
About this article
Cite this article
Nair, D.B., Uchida, K., Aizawa, SI. et al. Genetic analysis of a type IV pili-like locus in the archaeon Methanococcus maripaludis . Arch Microbiol 196, 179–191 (2014). https://doi.org/10.1007/s00203-014-0956-4
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00203-014-0956-4