
Abstract
Hydrogen oxidation and electron transport were studied in the chlorobenzene-utilizing anaerobe Dehalococcoides sp. strain CBDB1. While Cu2+ and Hg2+ ions irreversibly inhibited hydrogenase activity in intact cells, Ni2+ ions inhibited reversibly. About 80% of the initial hydrogenase activity was inactivated within 30 s when the cells were exposed to air. In contrast, hydrogenase was active at a redox potential of +10 mV when this redox potential was established anoxically with a redox indicator. Viologen dyes served both as electron acceptor for hydrogenase and electron donor for the dehalogenase. A menaquinone analogue, 2,3-dimethyl 1,4-naphthoquinone, served neither as electron acceptor for the hydrogenase nor as electron donor for the dehalogenase. In addition, the menaquinone antagonist 2-n-heptyl-4-hydroxyquinoline-N-oxide had no effect on dechlorination catalyzed by cell suspensions or isolated membranes with hydrogen as electron donor, lending further support to the notion that menaquinone is not involved in electron transport. The ionophores tetrachlorosalicylanilide and carbonylcyanide m-chlorophenylhydrazone did not inhibit dechlorination by cell suspensions, indicating that strain CBDB1 does not require reverse electron transport. The ATP-synthase inhibitor N,N′-dicyclohexylcarbodiimide inhibited the dechlorination reaction with cell suspensions; however, the latter effect was partially relieved by the addition of tetrachlorosalicylanilide. 1,2,3,4-Tetrachlorobenzene strongly inhibited dechlorination of other chlorobenzenes by cell suspensions with hydrogen as electron donor, but it did not interfere with either hydrogenase or dehalogenase activity.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
Dehalococcoides sp. strain CBDB1 is able to grow using hydrogen and chlorobenzenes (Adrian et al. 2000; Jayachandran et al. 2003) or chlorinated dioxins (Bunge et al. 2003) as sole energy source. Based on this ability, it was deduced that energy conservation must occur via a respiratory mechanism in which a proton-motive force (pmf) across the membrane is generated by electron transport through membrane-bound electron transport components. This pmf can be used by ATP-synthase to couple proton transfer back across the membrane for the synthesis of ATP. The electrochemical potential difference ΔE0′ between H2 (E0′ of H+/H2 = −414 mV) and the chlorobenzene couples (e.g., E0′ of HCB/PeCB = +478 mV) (Dolfing and Harrison 1992) is around 0.9 V.
Energy metabolism and electron transport have been studied in the PCE-dechlorinating strains Dehalobacter restrictus (Schumacher and Holliger 1996) and Sulfurospirillum multivorans (Miller et al. 1997) and the 3-chlorobenzoate dechlorinating bacterium Desulfomonile tiedjei DCB-1 (Louie and Mohn 1999). In Dehalococcoides sp. strain CBDB1, it was shown that the reductive dehalogenase is membrane bound, and previous experiments suggest that it is oriented to the outside of the cell. The reductive dehalogenase itself was not very sensitive to oxygen and was able to accept electrons from methyl and benzyl viologen (Hölscher et al. 2003). An in vitro assay of dehalogenase using various artificial electron donors showed that only compounds having redox potentials of ≤−360 mV were used by the enzyme (Hölscher et al. 2003). This suggested that a low-redox-potential electron donor is required for the reductive dechlorination of chlorobenzenes, although the standard redox potential of the chlorobenzene couples is always positive (e.g., E0′ of HCB/PeCB = +478 mV).
The results of Miller et al. (1997) indicated that reverse electron transport was required for the reductive dechlorination of PCE and that quinones were not involved in the electron transport chain of S. multivorans. In contrast, in another anaerobe, Dehalobacter restrictus, which is capable of dehalorespiration with PCE and hydrogen, menaquinone was involved in PCE reduction (Schumacher and Holliger 1996). Here we describe biochemical properties of hydrogenase and electron transport in a bacterium of a different phylogenetic group, Dehalococcoides sp. strain CBDB1.
Materials and methods
Inoculum and culture conditions
Strain CBDB1 was grown under strictly anaerobic conditions. The cells were cultivated in 120-ml flasks containing 60 ml medium and 60 ml gas phase. Crystals of PeCB (20 μg ml−1) or HCB (50 μg ml−1) were added as electron acceptors as described previously (Jayachandran et al. 2003). According to the method of Adrian et al. (2000), a synthetic mineral medium with vitamins and trace elements was reduced with 0.8 mM titanium(III) citrate, and 5 mM acetate as carbon source was added. The flasks were sealed with Teflon-lined butyl-rubber septa (Macherey and Nagel, Düren, Germany) and aluminum crimp caps. The headspace of each flask was exchanged with N2–CO2 (4:1, v/v) using an automatic gas exchanger. A nominal concentration of 7.5 mM hydrogen (10 ml) was added with a syringe to the headspace of the cultures using glass syringes previously flushed with sterile water. The standard inoculation volume was 200 μl active pure culture per flask. Cultures were incubated at 29°C in the dark without shaking.
Preparation of cell fractions
Cultures were harvested when they reached about 1 μg total protein ml−1 , which was approximately after 10 days of incubation. All handlings were done under anoxic conditions. If necessary, the culture was concentrated by centrifugation. Cell suspensions were flushed with nitrogen to remove hydrogen and lower chlorinated benzenes from the cultivation. Cells were disrupted with a small-scale ball mill (Ritsch Mixer Mill MM 200, Haan, Germany) using a volume ratio between culture and glass beads of 0.61 and a shaking time of 12.7 min; these were found to be the best parameters for maintenance of hydrogenase activity. The crude extract obtained after cell disruption was centrifuged at 10,000g for 20 min to remove intact cells. The supernatant was filled in a tube in an anoxic chamber and closed with an airtight Teflon-lined cap. After centrifugation at 120,000g for 60 min, the pellet was considered as the membrane fraction and the supernatant as the soluble fraction. The soluble fraction was further concentrated using either Sephadex G 25 coarse (1 g/5 ml) (Pharmacia Biotech, Uppsala, Sweden) or Centricon Plus-20 Centrifugal Filter Units (Millipore, Billerica, Mass., USA) with a molecular mass cut off of 5 kDa.
Analytical procedures
Chlorobenzene concentrations were quantified by gas chromatography from dichloromethane extracts as described previously (Jayachandran et al. 2003). Hydrogenase activity was assayed in a butyl-rubber stoppered glass cuvette measuring the hydrogen-dependent reduction of 1 mM benzyl viologen (ɛ=9.75 mM−1 cm−1) at 546 nm (Schneider and Schlegel 1976). The reaction mixture contained 1 mM benzyl viologen in anoxic 100 mM Tris–HCl at pH 7.5. A volume of 800 μl of assay solution was filled in a 1.6-ml anoxic cuvette in an anoxic chamber containing 5% H2. The reaction was started by the addition of 50-μl cell suspension or cellular fraction. The increase in A546 was monitored with a spectrophotometer (Uvikon 923 Double beam UV/VIS, Milan, Italy). Hydrogenase activity was expressed in nkat (nmol products per second at 25°C). Alternatively, 1 mM methyl viologen (ɛ=9.7 mM−1cm−1 at 578 nm), 1 mM ethyl viologen (ɛ=10 mM−1cm−1 at 578 nm), 1 mM AQDS (ɛ=1.8 mM−1cm−1 at 450 nm) or 1 mM DMN (ɛ=15.2 mM−1cm−1 at 270–290 nm) was used as electron acceptor for the hydrogenase. To study the effect of 1,2,3,4-TeCB on hydrogenase activity, AQDS, which is not oxidized by the dehalogenase, was used as electron acceptor. Diaphorase (NADH: benzyl viologen oxidoreductase) activity in the soluble fraction was assayed in a butyl-rubber-stoppered glass cuvette by monitoring the reduction of benzyl viologen (1 mM oxidized benzyl viologen was added) at 546 nm using 1 mM NADH as electron donor in the presence of nitrogen in the head space (Schneider and Schlegel 1976).
Cells were directly counted using a phase-contrast microscope (Axioskop 2 plus, Zeiss, Jena, Germany). Total protein was quantified by a fluorescent test (NanoOrange-kit, Molecular Probes, Leiden, The Netherlands). Fluorescence measurements were made on a RF-5001PC-fluorometer (Shimadzu, Tokyo, Japan).
Stability of hydrogenase activity in the presence of oxygen was investigated by stirring 5 ml of culture oxically. Redox potential was measured with a platinum–Ag/AgCl electrode (Pt 4800 M5-S7/80; Mettler-Toledo, Steinbach, Germany). A volume of 150 μl anoxic aqueous solution of resazurin (5 mg ml−1) was added to alter the redox potential of the culture anoxically. After a defined incubation time, titanium(III) citrate was added until the A546 of the samples increased to about 0.4 due to the reduction of benzyl viologen and hydrogenase activity was determined.
Dehalogenase activity was measured with reduced methyl viologen as electron donor as described by Hölscher et al. (2003). Tests for electron transport inhibitors contained 200 μl cell suspensions with about 5×107 cells ml−1, 50 μM 1,2,3-TCB, 2 mM titanium(III) citrate and inhibitors or ionophores in 100 mM Tris–HCl buffer at pH 7.5. The assay solution was filled in 2 ml vials in an anoxic chamber containing 5% H2. The vials were stoppered with Teflon-lined butyl-rubber septa and aluminum crimp caps. The tests were incubated overnight at 25°C and the dechlorinated products were analyzed by gas chromatography as described previously. Where indicated, 1 μM HOQNO, 0.5 mM DCCD, 1 μM TCS, 1 μM CCCP or 10 μM 1,2,3,4-TeCB was added.
Source of chemicals
All chlorobenzenes were obtained from Aldrich (Steinheim, Germany), Merck–Schuchardt (Hohenbrunn, Germany), Arcos (NJ, USA) or Fluka (Buchs, Switzerland). Titanium chloride (synthesis-grade solution) was obtained from Merck–Schuchardt. Titanium(III) citrate [0.1 M with respect to Ti(III)] was prepared as described by Zehnder and Wuhrmann (1976). Benzyl viologen, methyl viologen, ethyl viologen, AQDS and DMN were obtained from Aldrich, TCS from Arcos. All other chemicals used were of analytical grade and were purchased from Sigma (Deisenhofen, Germany) or Merck. Gases, N2, H2 (99.999%, v/v) and CO2 (99.8%, v/v) were obtained from Messer Griesheim (Berlin, Germany) and traces of oxygen were removed by a reduction column (Ochs, Göttingen, Germany).
Results
Electron donors and acceptors for hydrogenase and dehalogenase activities
Hydrogenase activity was detected with whole cells of strain CBDB1. Among the compounds tested as electron acceptors with hydrogen as the electron donor, the highest activity was detected with benzyl viologen (Table 1). Whole cells of strain CBDB1 also reduced methyl viologen, ethyl viologen and AQDS but not DMN (Table 1). Electron acceptors were only reduced if the headspace of the cuvettes contained hydrogen. Among the redox compounds tested as artificial electron donors for the 1,2,3-dehalogenase, only viologens were effective electron donors (Table 1). The highest enzyme activity was measured with methyl viologen (E0′ = −450 mV). The quinone analogue AQDS and the menaquinone derivative DMNH2 did not serve as electron donors for the 1,2,3-TCB dehalogenase reaction. No dechlorination was observed in the absence of any added electron donor.
Localization and oxygen sensitivity of the hydrogenase activity
Hydrogenase activity was detected in the membrane as well as in the soluble fraction after fractionation of cell components (Table 2). Total activity of both fractions together, however, amounted to only about 20% of the activity found with whole cells. In the soluble fraction, in addition to hydrogenase activity, diaphorase activity was detected. Diaphorase activity was about 2.5-fold higher than hydrogenase activity (Table 2). Whole-cell hydrogenase activity was irreversibly inhibited by 75 μM Cu2+ or Hg2+. The inhibitory effect of Ni2+ on hydrogenase activity of both membrane and soluble fractions could be reversed by increasing concentrations of benzyl viologen (Fig. 1).

Competitive inhibition of hydrogenase by Ni2+. Double-reciprocal plots of the rate of hydrogen oxidation catalyzed by a membrane and b soluble fractions of strain CBDB1 in the presence (filled squares) and absence (open squares) of 75 μM Ni2+. The reaction mixture contained 100 mM Tris–HCl buffer at pH 7.5 in the presence of a nominal concentration of 2 mM H2. S, concentration of the substrate benzyl viologen
Hydrogenase activity was not detected in cell suspensions of strain CBDB1 in the absence of titanium(III) citrate even under strictly anoxic conditions. This suggested that the presence of reducing agent is necessary to maintain the stability of the enzyme. Early experiments in assessing the oxygen sensitivity of enzymes from strain CBDB1 showed that hydrogenase activity of intact cells was completely lost within 5 min of storage in the presence of oxygen. In contrast, dehalogenase activity of strain CBDB1 could still be detected after 20 h of storage in the presence of oxygen (Hölscher et al. 2003). About 80% of hydrogenase activity was inactivated irreversibly within 30 s when cells were exposed to air. During this time, the redox potential of the medium increased from −190 mV to −100 mV (Fig. 2). However, samples with a redox potential of about +10 mV, established anoxically with the redox dye resazurin, showed about 80% of the hydrogenase activity found with control sample without resazurin (134 nkat/mg protein with resazurin; 168 nkat/mg protein without resazurin). This showed that hydrogenase of strain CBDB1 stayed active at a positive redox potential in the absence of oxygen. However, after 30 min, hydrogenase activity was lost in the sample with resazurin as well as in the control, yielding redox potentials of +470 mV and +110 mV, respectively.

Hydrogenase activity measured with whole cells of strain CBDB1 at various redox potentials in the presence of oxygen. Hydrogenase activity and redox potential were measured with samples taken every 30 s while the culture was stirred oxically as described in “Materials and methods”. (filled squares) Redox potential, (open squares) hydrogenase activity. Values are means of two independent experiments
Electron transport
Isolated membrane fractions reductively dechlorinated 1,2,3-TCB to 1,3-DCB (6.3±0.4 μM after overnight incubation) with hydrogen as the electron donor. This demonstrated that all components needed for oxidation of hydrogen, transport of electrons and reduction of chlorobenzenes were associated with the membrane. The dechlorination rate was not increased after the addition of soluble fraction of strain CBDB1. Also, the soluble fraction alone did not catalyze dechlorination.
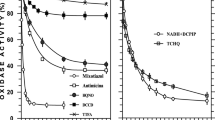
HOQNO, an antagonist of menaquinone-dependent redox reactions (Zhao and Weiner 1998), did not inhibit dechlorination with H2 as the electron donor in intact cell suspensions (Fig. 3) or isolated membranes (6.06±0.4 μM 1,3-DCB after overnight incubation). Both TCS and CCCP, protonophores that dissipate membrane potential and proton gradient, did not inhibit dechlorination of 1,2,3-trichlorobenzene by cell suspensions (Fig. 3). DCCD inhibits ATPase by blocking the proton channels. As a result, an influx of protons into the cell is prevented (White 1995). In the presence of 0.5 mM DCCD, dechlorination of 1,2,3-TCB by intact cells was almost completely inhibited (Fig. 3). The same concentration of DCCD did not have any effect on either hydrogenase or dehalogenase activity of strain CBDB1 when the enzyme activities were measured separately with benzyl viologen as electron acceptor or donor, respectively. The inhibitory effect of DCCD on dechlorination by whole cells was partially released when 1 μM TCS was added (Fig. 3).

Effect of CCCP (1 μM), TCS (1 μM), HOQNO (1 μM) and DCCD (0.5 mM) on reductive dechlorination of 1,2,3-TCB by cell suspensions of strain CBDB1 in the presence of a nominal concentration of 2 mM H2 as electron donor
In the dehalogenase activity test with reduced methyl viologen as electron donor, the highest specific activity was found when 1,2,3,4-TeCB was used as electron acceptor (Jayachandran et al. 2003; Hölscher et al. 2003). However, strain CBDB1 did not grow with 1,2,3,4-TeCB as electron acceptor. In fact, the addition of a saturating concentration of 1,2,3,4-TeCB prevented growth on other chlorobenzenes (Jayachandran et al. 2003). The dechlorination of 1,2,3-TCB, 1,2,4,5-TeCB, 1,2,3,5-TeCB or PeCB catalyzed by whole cells of strain CBDB1 with hydrogen as electron donor was completely inhibited by 10 μM 1,2,3,4-TeCB (Table 3). Neither 1,2,3,5-TeCB nor 1,2,4,5-TeCB inhibited dehalogenation of 1,2,3-TCB, or PeCB by cell suspensions at this concentration. Significant inhibition of 1,2,3-TCB dechlorination was observed even at 1 μM 1,2,3,4-TeCB (Table 3).
The effect of 1,2,3,4-TeCB on hydrogenase activity was measured using AQDS as electron acceptor. AQDS was reduced by the hydrogenase but the dehalogenase could not use the reduced AQDS as electron donor for reduction of 1,2,3,4-TCB. Therefore, AQDS accumulation could be measured. With hydrogen as an electron donor 10 μM 1,2,3,4-TeCB did not interfere with AQDS reduction catalyzed by whole cells. 1,2,3,4-TeCB at 50 μM inhibited the hydrogenase activity of strain CBDB1 by about 25%.
Discussion
Cu2+ and Hg2+ have been described to inactivate [Ni–Fe] hydrogenases irreversibly (Llama et al. 1979; Cypionka and Dilling 1986). Competitive inhibition by Ni2+ has been demonstrated with the [Ni–Fe] hydrogenase of the purple sulfur bacterium Thiocapsa roseopersicina (Zadvorny et al. 2000). The inhibition effects found with hydrogenase activity of strain CBDB1 suggest that this enzyme is a [Ni–Fe] containing hydrogenase. While dehalogenase in crude extract of strain CBDB1 was still active even after 20 h of storage in the presence of oxygen (Hölscher et al. 2003), hydrogenase activity was highly sensitive towards oxygen. The instability of hydrogenase towards oxygen could be the main reason that the strictly hydrogenotrophic strain almost instantly loses its viability when exposed to air.
Putative electron mediators
Both hydrogenase and dehalogenase of strain CBDB1 were demonstrated to use artificial electron donors or acceptors. However, we were not able to identify the in vivo redox compound(s) that mediate electron transport between the two enzymes. Schumacher and Holliger (1996) observed the involvement of menaquinone in PCE reduction by D. restrictus. In contrast, in another PCE-dehalogenating anaerobe, S. multivorans, menaquinone seemed not to participate in electron transport (Miller et al. 1997). The lack of inhibition of dechlorination by the menaquinone antagonist HOQNO indicated that menaquinone is not a respiratory component in strain CBDB1. Failure of the menaquinone analogue DMNH2 to support 1,2,3-TCB dechlorination also supported the exclusion of menaquinone in the electron transport of strain CBDB1.
Miller et al. (1997) proposed a reaction mechanism for the PCE reductive dehalogenase of S. multivorans, according to which a reverse electron flow driven by an electrochemical proton potential is involved to reduce cob(II)alamin to cob(I)alamin. Recently, the co-factor of the PCE reductive dehalogenase of S. multivorans has been shown to be a norpseudovitamin B12 (Kräutler et al. 2003). In strain CBDB1, however, a proton potential was not required for chlorobenzene dechlorination, as was shown with both protonophores TCS and CCCP which did not inhibit dechlorination. Also, the detection of dechlorination activity with isolated membrane fractions and hydrogen as the electron donor demonstrated that a reversed electron flow is not necessary for dechlorination. Similar results were found for the PCE reductive dehalogenase of Dehalococcoides ethenogenes strain 195 (Nijenhuis 2002). The significant differences between S. multivorans and Dehalococcoides sp. could be due to differences in the electron transport chain, the localization of the dehalogenases, or the co-factors.
Inhibition of electron transport by 1,2,3,4-TeCB
In the presence of 1,2,3,4-TeCB, the hydrogen-dependent reductive dechlorination of other chlorobenzenes by cell suspensions is abolished. Since 1,2,3,4-TeCB does not inhibit hydrogenase or dehalogenase activity directly, it must somehow interfere with electron transport in strain CBDB1. The inhibition by 1,2,3,4-TeCB seems to be specific because a significant inhibitory effect was observed at a concentration of 1 μM. It might be that the chemical structure of 1,2,3,4-TeCB resembles one of the electron-mediating components and competes with this putative mediator at its binding site. It will be of interest to see whether 1,2,3,4-TeCB inhibits dehalorespiration in organisms other than strain CBDB1.
Topology of respiratory enzymes involved in energy metabolism
In strain CBDB1, dehalogenase, hydrogenase and all electron mediating components are associated with the membrane. Reduced methyl viologen, a monovalent membrane non-permeable cationic radical (Jones and Garland 1977), acted as artificial electron donor for the reduction of 1,2,3-TCB with intact cells, suggesting that the active site of dehalogenase is oriented to the outside of the cytoplasmic membrane (Hölscher et al. 2003). Analogously, inhibition of hydrogenase activity with intact cells by membrane non-permeable Cu2+ ions (Cypionka and Dilling 1986) suggested that the active site of hydrogenase is also oriented to the outside.
Since fermentative growth is unknown with H2, strain CBDB1 must derive its energy by a respiratory process in which a pmf is generated across the membrane. A pmf can be used for ATP synthesis by ATPase. The involvement of ATPase in the respiration of strain CBDB1 was demonstrated in this study, showing inhibition of the dehalogenation of chlorobenzenes by the ATPase inhibitor DCCD. Because no protons can flow into the cell via ATPase in the presence of DCCD, the respiration chain leading concomitantly to the export of protons from the cytoplasm also comes to a halt. This model is supported by the result that TCS relieves the inhibition by DCCD, because TCS would allow protons to flow into the cell independently from inactive ATPase, and the respiration chain could resume proton export, however, of course without ATP generation.
Theoretically, the potential difference ΔE0′ = +892 mV between H2 and HCB (Dolfing and Harrison 1992) could generate 2–3 ATP/Cl− by a chemiosmotic mechanism, adapting the estimation done by Kröger et al. (2002) for Wolinella succinogenes. During HCB respiration by strain CBDB1 using hydrogen as electron donor and acetate as carbon source, a total cell protein yield of at most 2.1±0.24 g cell protein/mol Cl− released was determined (Jayachandran et al. 2003). Assuming that the protein content accounts for 50% of the dry cell weight (Mackiewicz and Wiegel 1998), a maximum of 4.2 g dry cell mass/mol Cl− can be expected. With a molar growth yield (YATP) of about 7 g cell dry mass per mol ATP, a value that was also used by Miller et al. (1997) for the PCE-respiring S. multivorans following Loubiere and Lindely (1991), only about 0.6 mol of ATP per mol Cl− released would be estimated for strain CBDB1. This difference between the theoretical yield of 2–3 ATP/Cl− and the observed yield of about 0.6 ATP/Cl− must have biochemical reasons. A reverse electron-transport process, which would consume energy and lead to a lower cell yield, was ruled out by using the protonophores. An explanation that cannot currently be ruled out is that the structure of the electron transport chain is very simple, leading to loss of the majority of the available energy as heat. This large increase in entropy would allow the reaction to proceed even if the chlorinated electron acceptors are available at only very low concentration, a condition common in natural environments.
Abbreviations
- AQDS:
-
Anthraquinone-2,6-disulfonate
- CCCP:
-
Carbonylcyanide m-chlorophenylhydrazone
- DCCD:
-
N,N′dicyclohexylcarbodiimide
- DMN:
-
2,3-Dimethyl 1,4-naphtoquinone
- HCB:
-
Hexachlorobenzene
- HOQNO:
-
2-n-Heptyl-4-hydroxyquinoline-N-oxide
- PCE:
-
Tetrachloroethene
- PeCB:
-
Pentachlorobenzene
- TeCB:
-
Tetrachlorobenzene
- TCS:
-
3,3′,4′,5-Tetrachlorosalicylanilide
- TCE:
-
Trichloroethene
References
Adrian L, Szewzyk U, Wecke J, Görisch H (2000) Bacterial dehalorespiration with chlorinated benzenes. Nature 408:580–583
Bunge M, Adrian L, Kraus A, Opel M, Lorenz WG, Andreesen JR, Görisch H, Lechner U (2003) Reductive dehalogenation of chlorinated dioxins by an anaerobic bacterium. Nature 421:357–360
Cypionka H, Dilling W (1986) Intracellular localization of the hydrogenase in Desulfotomaculum orientis. FEMS Microbiol Lett 36:257–260
Dolfing J, Harrison BK (1992) The Gibbs free energy of formation of halogenated aromatic compounds and their potential role as electron acceptors in anaerobic environments. Environ Sci Technol 26:2213–2218
Hölscher T, Görisch H, Adrian L (2003) Reductive dehalogenation of chlorobenzene congeners in cell extracts of Dehalococcoides sp. strain CBDB1. Appl Environ Microbiol 69:2999–3001
Jayachandran G, Görisch H, Adrian L (2003) Dehalorespiration with hexachlorobenzene and pentachlorobenzene by Dehalococcoides sp. strain CBDB1. Arch Microbiol 180:411–416
Jones RW, Garland PB (1977) Sites and specificity of the reaction of bipyridylium compounds with anaerobic respiratory enzymes of Escherichia coli: effects of permeability barriers imposed by the cytoplasmic membrane. Biochem J 1641:99–211
Kräutler B, Fieber W, Ostermann S, Fasching M, Ongania KH, Gruber K, Kratky C, Mikl C, Siebert A, Diekert G (2003) The cofactor of tetrachloroethene reductive dehalogenase of Dehalospirillum multivorans is norpseudo-B12, a new type of a natural corrinoid. Helv Chim Acta 86:3698–3716
Kröger A, Biel S, Simon J, Gross R, Unden G, Lancaster CRD (2002) Fumarate respiration of Wolinella succinogenes: enzymology, energetics and coupling mechanism. Biochim Biophys Acta 1553:23–38
Llama MJ, Serra JL, Rao KK, Hall DO (1979) Isolation and characterization of the hydrogenase activity from the non-heterocystous cyanobacterium Spirulina maxima. FEBS Lett 98:342–346
Loubiere P, Lindely ND (1991) The use of acetate as additional co-substrate improves methylotropic growth of the acetogenic anaerobe Eubacterium limosum when CO2 fixation is rate-limiting. J Gen Microbiol 137:2247–2251
Louie TM, Mohn WW (1999) Evidence for a chemiosmotic model of dehalorespiration in Desulfomonile tiedjei DCB-1. J Bacteriol 181:40–46
Mackiewicz M, Wiegel J (1998) Comparison of energy and growth yields for Desulfitobacterium dehalogenans during utilization of chlorophenol and various traditional electron acceptors. Appl Environ Microbiol 64:352–355
Miller E, Wohlfarth G, Diekert G (1997) Studies on tetrachloroethene respiration in Dehalospirillum multivorans. Arch Microbiol 166:379–387
Nijenhuis I (2002) Dehalorespiration by Dehalococcoides ethenogenes strain 195 and relatives. Ph.D thesis, Cornell University
Schneider K, Schlegel HG (1976) Purification and properties of soluble hydrogenase from Alcaligenes eutrophus H16. Biochim Biophys Acta 452:66–80
Schumacher W, Holliger C (1996) The proton/electron ratio of the menaquinone-dependent electron transport from dihydrogen to tetrachloroethene in “Dehalobacter restrictus”. J Bacteriol 178:2328–2333
White D (1995) The physiology and biochemistry of prokaryotes. Oxford University Press, New York
Zadvorny OA, Zorin NA, Gogotov IN (2000) Influence of metal ions on hydrogenase from the purple sulfur bacterium Thiocapsa roseopersicina. Biochemistry (Mosc) 65:1287–1291
Zehnder AJB, Wuhrmann K (1976) Titanium(III) citrate as a non toxic oxidation–reduction buffering system for the culture of obligate anaerobes. Science 194:1165–1166
Zhao Z, Weiner JH (1998) Interaction of 2-n-heptyl-4-hydroxyquinoline N-oxide with dimethlyl sufoxide reductase of Escherichia coli. J Biol Chem 273:20758–20763
Acknowledgment
This work was supported by the DAAD and the DFG, grant AD178/1.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Jayachandran, G., Görisch, H. & Adrian, L. Studies on hydrogenase activity and chlorobenzene respiration in Dehalococcoides sp. strain CBDB1. Arch Microbiol 182, 498–504 (2004). https://doi.org/10.1007/s00203-004-0734-9
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00203-004-0734-9