
Abstract
Three kinds of representative sediments were obtained from a macrophyte-dominated bay (East Lake Taihu) and two algae-dominated regions (Western Lake Taihu and Meiliang Bay). Physiological responses of Vallisneria asiatica to these sediments were compared. Results from 20 days exposures showed no obvious differences in malondialdehyde (MDA) in roots, while the MDA content in leaves of plants exposed to Western Lake Taihu sediment was significantly (p < 0.05) higher than those exposed to the other two sediments. In comparison to the other two sediments, plants exposed to Western Lake Taihu sediment showed significantly lower (p < 0.05) superoxide dismutase in roots and leaves on the 10th and 40th day. On the 40th day, root catalase (CAT) activities in V. asiatica from Western Lake Taihu and Meiliang Bay sediments were lower than that from East Lake Taihu sediment, while leaf CAT activity in V. asiatica from Western Lake Taihu sediment was higher than that from East Lake Taihu sediment (p < 0.05). Western Lake Taihu sediment caused more serious oxidative stress in V. asiatica than East Lake Taihu sediment. Results indicated eutrophic sediment was a contributing factor in the disappearance of V. asiatica in Western Lake Taihu.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Submersed macrophytes can improve the water quality by inhibiting the growth of algae (Nakai et al. 2000), absorb excessive nutrients (Lesage et al. 2007) and prevent resuspension of sediment (Madsen et al. 2001). Thus, they play an important role in the ecological structure and function of a healthy lake (Engel 1998) by improving the self-purification capacity of a lake ecosystem. Submerged macrophytes experience adverse effects when changes in environmental factors (including temperature, light, and the concentrations of pollutants and nutrients) exceed their required ranges within which the plants can grow and breed normally (Foyer and Noctor 1999). According to previous studies, adverse conditions, such as high concentrations of nutrients and metals, will cause visible damage or growth reduction of submerged macrophytes. Additionally, contaminants can also induce a series of antioxidant responses in cells because of the excessive reactive oxygen species (ROS) induced by adversities (Dučić and Polle 2005; Wang et al. 2009). Eutrophic sediment will also induce either a massive production of ROS or destroy the scavenging system of ROS, resulting in an accumulation of ROS (Wang et al. 2009). Plants are equipped with a defense system to remove excessive ROS, thereby repairing damage caused by ROS. Antioxidant enzymes play an important role in this detoxification process. Among the antioxidant enzymes, superoxide dismutase (SOD) acts as the first line of defense against ROS by degrading superoxide to H2O2 (Bowler et al. 1992). Thereafter, catalase (CAT) can decompose H2O2 to water and oxygen (Pflugmacher 2004). Lipid peroxidation will occur when the lipid of a cell membrane is attacked by ROS because of a deficient antioxidant defense, and as a consequence, this damage to cell membranes may lead to a decline in growth rate and biomass (Kabala et al. 2008). Malondialdehyde (MDA), as a secondary end product of lipid peroxidation, is commonly used to detect the degree of lipid peroxidation and oxidative damage in roots and leaves of plants (Peutherta and Pflugmacher 2010).
Eutrophication of shallow lakes, such as that of Lake Taihu in China, has been a global environmental issue in recent years (Codd 1995). A field investigation in Lake Taihu showed Vallisneria asiatica and other submerged macrophytes started to decrease in 2000, coinciding with increased eutrophication (Zhu 2008). Currently, considerable coverage of submerged macrophytes only occurs in East Lake Taihu, with few emergent plants distributed in Western Lake Taihu, and no aquatic macrophytes present in Meiliang Bay. V. asiatica is a common species of submerged macrophytes in Lake Taihu, but the population is currently in decline. In order to determine whether the eutrophic sediment is a contributing factor in the disappearance of V. asiatica in Western Lake Taihu and Meiliang Bay, it is important to investigate responses of V. asiatica to the eutrophic sediment of Lake Taihu prior to the transplantation of this species to the lake. The objective of this study was to compare the antioxidant responses of V. asiatica to the sediments from algae-dominated regions and the macrophyte-dominated region.
Methods and Materials
The submerged macrophyte, V. asiatica, was collected in Onga River, Fukuoka, Japan, and then cultured in dechlorinated tap water for 5 days to remove attached sediments and old branches were additionally removed. Only plants showing healthy growth and of a uniform size were chosen for the experiment. Surface sediment was collected from Lake Taihu using a Van Veen grab operated from a small boat at the sites shown in Fig. 1. Site 1 is located in East Lake Taihu, which is a region characterized by high coverage of submerged macrophytes (Qu et al. 2001). Site 2 is located along Western Lake Taihu, which is polluted and regularly covered by thick algal blooms from late spring into autumn because of the developed agriculture and forestry in that area (Qu et al. 2001). Site 3 is located in Meiliang Bay, one of most polluted zones in Lake Taihu (Qu et al. 2001). Basic characteristics of the sediment are summarized in Table 1. Compared with East Lake Taihu sediment, the sediment of Western Lake Taihu was enriched with organic matter (OM), nutrients, and heavy metals, and the sediment of Meiliang Bay contained high contents of total nitrogen (TN) and total phosphorus (TP).

Sampling sites of sediments in Lake Taihu, China
The three sediments were sieved (0.2 cm) to remove coarse debris, homogenized, and then placed into beakers (500 mL). Sediment was placed in beakers to inhibit or weaken the release of inorganic forms of N, P, Mn, Cu, Fe, etc. and difficult to degrade organic pollutants from sediments to water. For each type of sediment, three beakers containing sediment were placed in a strengthened glass tank with a volume of 40 L. A total of 18 healthy even-sized plants (approximately 80 g) were randomly planted in each beaker. Each tank (30 × 30 × 45 cm3) was filled with 30 L of dechlorinated tap water (containing 0.72 mg L−1 NO3 −–N and 0.02 mg L−1 PO4 3−–P). Three tanks were placed in a controlled environment (temperature = 25°C; illumination intensity = 3400 lux; photoperiod = 12 h:12 h). Three plants from each tank (one plant per beaker) were randomly selected for the biochemical analysis during days 10, 20, and 40.
The pH in the sediment was measured using portable electrodes. Sediment particles smaller than 100 μm were used for all analyses and acid digestion. The proportion of organic matter (OM) in the sediment can be reflected by weight loss through combustion (450°C, 3 h) (Kleeberg et al. 2010). To determine the metal and nutrient contents, sediment samples were digested by H2SO4 and H2O2 (Kleeberg et al. 2010). Major elements such as P and metals were determined by Inductively Coupled Plasma Optical Emission Spectrometer (ICP-OES, Optima 5300DV). Nitrogen was measured by the TN analysis method (Hach 2002).
Plant samples were ground and homogenized in 8 mL of phosphate buffer (50 mM, pH 7.0), containing 3 mM EDTA-2Na and 0.5 % (w/v) PVPP and then centrifuged at 12,000×g for 10 min to obtain the supernatant as extractions for analyzing the MDA content and enzyme activity. MDA content determination was based on Heath and Packer (1968). Briefly, 2 mL of extract was mixed with 2 mL of 0.6 % thiobarbituric acid in 20 % trichloroacetic acid (TCA). The mixture was heated in boiling water for 20 min, after which it was immediately cooled, and centrifuged at 12,000 ×g for 10 min. The supernatant was measured by the UV–visible spectrophotometer (UV-1600PC), and the MDA content was calculated from the difference in the absorbance at 532 and 600 nm. SOD activity was assayed by monitoring the inhibition of photochemical reduction of nitro-blue tetrazolium (NBT) (Beauchamp and Fridovich 1971). The reagent for SOD activity estimation was prepared by mixing 1.1 mL phosphorus buffer (pH 7.8), 0.1 mL of 8 mM hydroxylammonium chloride, 0.1 mL of 20 % extraction, and 0.1 mL of 8 mM xanthine in a tube. Then, 0.1 mL xanthine oxidase (5.87 U/mL) was added to the tube every 10 s. A total of 1 mL of 20 mM sulfanilic acid was added in each test tube to stop the reaction every 10 s after the incubation in water for 40 min at 30°C. A total of 1 mL of 10 mM N-(1-naphthyl) ethylenediamine dihydrochloride was added. After 20 min, the absorbance at 530 nm was measured. One unit of the enzyme activity is defined as the amount of enzyme required to generate a 50 % inhibition of the rate of NBT reduction measured at 530 nm (Li et al. 2002). CAT was determined by measuring the yield of oxygen in the reaction between extractions and H2O2 at the temperature of 24°C and ordinary pressure within 1 min (Greenfield and Price 1954).
Data were reported as mean ± standard deviation (n = 3). Statistical analysis was performed by using one-way ANOVA followed by the LSD-test, at p < 0.05. Different letters indicate significant differences (p < 0.05) between three types of sediment by a one-way ANOVA test.
Results and Discussion
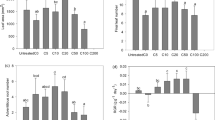
In Fig. 2, no differences in root MDA activity among the three kinds of sediment were observed. Low MDA contents indicated the membrane structure of root cells was not damaged, possibly in part because of the activated oxidative enzymes. Additionally, the roots potentially possess alternative means (e.g., translocation or complexation) of coping with high metal levels to protect from oxidative damage (Dučić and Polle 2005). The 20 and 40 days MDA content in V. asiatica leaves from Western Lake Taihu sediment was significantly higher than that growing on East Lake Taihu sediment. Furthermore, for the sediment from Western Lake Taihu, 40 days leaf MDA content was considerably higher than that at 20 days. This indicated V. asiatica growing on Western Lake Taihu sediment were under oxidative stress, compared with those growing on East Lake Taihu sediment. The oxidative stress response in V. asiatica to Western Lake Taihu sediment included the increased MDA content after 10 days exposure. Fertile sediment typically has higher N and P content (Ye et al. 2009) and is considered to be an important nutrient source for rooted submerged macrophytes (Ye et al. 2009). Compared with brown clay encountered in other studies, Lake Taihu sediment was enriched in organic matter and nutrients (Ye et al. 2009; Yu et al. 2010). Elements such as Cu, Mn, and Zn are essential nutrients necessary for normal growth and development of plants (Dučić and Polle 2005). Therefore, fertile sediment plays a positive role for the growth of V. asiatica because of abundant nutrients. However, it is believed that the growth of submerged macrophytes is negatively affected by the sediment when nutrient concentrations in sediment exceed the plant’s tolerance limits (Zhang et al. 2008). Compared with East Lake Taihu sediment, Western Lake Taihu sediment contained much more N, P, Mn, Cu and Fe, therefore, this sediment resulted in greater oxidative stress possibly because nutrient and metal (Mn, Cu and Fe) contents exceeded V. asiatica tolerance limits. Many researchers also have reported that high concentrations of Cu and Zn can cause oxidative damage to aquatic plants, expressed as low contents of chlorophyll and protein and high MDA content (Wang et al. 2009; Xing et al. 2010). However, for the Meiliang Bay sediment, there was no increase in MDA content in V. asiatica, although this sediment contained higher nutrients than that of East Lake Taihu sediment. The sediment from different sites differed in numerous constituents. Therefore, it is difficult to analyze all the constituents. In this study, only the main variables listed in Table 1 were analyzed, because it is well known that sediments in eutrophic Lake Taihu contain excessive nutrients (especially N and P), as well as toxic metals because large amounts of untreated effluent from surrounding industries and agriculture are discharged into Lake Taihu (Qu et al. 2001). Thus, the high content of N, P, Mn, Cu and Fe in Western Lake Taihu sediment is just one of the important factors which resulted in oxidative stress to V. asiatica. Compared with Western Lake Taihu sediment, Meiliang Bay sediment contained relative low contents of OM, TN and metals. High concentration of TP in Meiliang Bay sediment cannot result in the oxidative damage for V. asiatica. For Western Lake Taihu sediment, oxidative damage possibly occurred because of the high contents of nutrients and metals. V. asiatica can induce antioxidant defense against oxidative stress caused by the high contents of nutrients in the Meiliang Bay sediment.

Changes of MDA contents in a roots and b leaves of Vallisneria asiatica. Different letters indicate significant differences at the <0.05 level
In this study, the exposure of sediment from algae-dominated regions to V. asiatica led to oxidative stress and antioxidant defense, but the response in roots and leaves of V. asiatica were different. Roots of V. asiatica responded to the eutrophic sediments during the early period of the experiment. Results showed the SOD was suppressed when V. asiatica were exposed to the Western Lake Taihu sediment for 10 days (Fig. 3). When superoxide radical generation exceeds the elimination ability of SOD, superoxide radicals, as well as other oxyradicals, can inactivate the enzyme SOD (Yin et al. 2008). Excessive ROS can induce the antioxidant responses of plants to oxidative stresses, including the enhancement or the suppression of antioxidant enzyme activities (Jayakumar et al. 2006). The suppressed SOD in turn caused oxidative damage to V. asiatica because of the resulting low ROS elimination capacity. During the first period, the roots activated a defense system in cells which could cope with the eutrophic sediment. For example, compared with East Lake Taihu sediment, the CAT activity in leaves of V. asiatica exposed to eutrophic sediments from algae-dominated regions was higher on day 10, and then it became lower on day 40 (Fig. 3). Compared with Western Lake Taihu sediment, Meiliang Bay sediment cannot suppress the SOD based solely on the high concentration of nutrients. The induction of antioxidant enzyme CAT indicated the ongoing detoxification process in V. asiatica to resist oxidative stress from the eutrophic sediment (Pflugmacher 2004). However, the detoxification ability of CAT decreased with time expressed as the decreased CAT for the eutrophic sediment from algae-dominated regions on day 40, compared with East Lake Taihu sediment. In addition, at later times, there were no differences in the SOD activity in roots of V. asiatica among the three kinds of sediment in Lake Taihu.

Changes of a SOD and b CAT activities in roots of Vallisneria asiatica. Different letters indicate significant differences at the <0.05 level
In contrast to roots, the defense system of leaves responded to the stress from eutrophic sediments during the later period. As a consequence, compared with East Lake Taihu sediment, decreased SOD activity in V. asiatica growing on Western Lake Taihu sediment was evident on day 40 and the CAT activity for Western Lake Taihu sediment was higher than other sediments (Fig. 4). Furthermore, low SOD activities resulted in injury to V. asiatica because of resulting reduced ability to scavenge excessive ROS (Blokhina et al. 2003). Low SOD activities were also found in leaves of submerged macrophytes in Lake Taihu (Zhang et al. 2011). If the cellular antioxidant defense is deficient, excessive ROS are able to rapidly attack membrane lipids resulting in lipid peroxidation (Peutherta and Pflugmacher 2010). Lipid peroxidation was reflected by the increased MDA content in leaves during the later period. Negative effects of excessive ROS induced by the eutrophic sediment on V. asiatica mainly include the inactivation of the enzyme SOD through the oxidation of the tryptophan residues near the active-site metals (Fe, Cu and Zn) by hydrogen peroxide, and the lipid peroxidation through the degradation of polyunsaturated fatty acids in the membrane lipids undergoing the stress (Bowler et al. 1992).

Changes of a SOD and b CAT activities in leaves of Vallisneria asiatica. Different letters indicate significant differences at the <0.05 level
In conclusion, compared with East Lake Taihu sediment and Meiliang Bay sediment, the oxidative damage in V. asiatica caused by Western Lake Taihu sediment was more serious because of high concentrations of nutrients and metals. High contents of nutrients for Meiliang Bay sediment cannot induce the lipid peroxidation of membranes compared with East Lake Taihu. V. asiatica can survive in East Lake Tahu sediment by adjusting its antioxidant enzymes. Results also indicated that roots of V. asiatica responded to the eutrophic sediment immediately, because they were in direct contact with sediment. Leaves of V. asiatica exhibited their responses to the eutrophic sediment during the later period of the experiment after the transportation of elements in sediment from roots to leaves.
References
Beauchamp C, Fridovich I (1971) Superoxide dismutases: improved assays and an assay applicable to acrylamide gels. Anal Biochem 44:276–287
Blokhina O, Virolainen E, Fagerstedt KV (2003) Antioxidants, oxidative damage and oxygen deprivation stress: a review. Ann Bot-Lond 91:179–194
Bowler C, Montagu MV, Inze D (1992) Superoxide dismutase and stress tolerance. Annu Rev Plant Biol 43:83–116
Codd GA (1995) Cyanobacterial toxins: occurrence, properties and biological significance. Water Sci Technol 32:149–156
Dučić T, Polle A (2005) Transport and detoxification of manganese and copper in plants. Braz J Plant Physiol 17:103–112
Engel S (1998) The role and interaction of submerged macrophytes in a shallow Wisconsin Lake. J Freshw Ecol 4:329–341
Foyer CH, Noctor G (1999) Leaves in the dark see the light. Science 284:599–601
Greenfield RE, Price VE (1954) Liver catalase Ι: a. manometric determination of catalase activity. J Biol Chem 209:355–361
Hach (2002) Water analysis handbook of DR 2400 portable spectrophotometer. HACH Company, Loveland
Heath RL, Packer L (1968) Photoperoxidation in isolated chloroplasts Ι. Kinetics and stoichiometry of fatty acid peroxidation. Arch Biochem Biophys 125:189–198
Jayakumar AR, Panickar KS, Murthy CR, Norenberg MD (2006) Oxidative stress and mitogen-activated protein kinase phosphorylation mediate ammonia-induced cell swelling and glutamate uptake inhibition in cultured astrocytes. J Neurosci 26:4774–4784
Kabala KM, Janicka-Russak M, Burzynski M, Klobus G (2008) Comparison of heavy metal effect on the proton pumps of plasma membrane and tonoplast in cucumber root cells. J Plant Physiol 165:278–288
Kleeberg A, Herzog C, Jordan S, Hupfer M (2010) What drives the evolution of the sedimentary phosphorus cycle? Limnologica 40:102–113
Lesage E, Mundia C, Rousseau DPL, Van de Moortel AMK, Laing GD, Meers E, Tack FMG, De Pauw N, Verloo MG (2007) Sorption of Co, Cu, Ni and Zn from industrial effluents by the submerged aquatic macrophyte Myriophyllum spicatum L. Ecol Eng 30:320–325
Li JC, Maezawa S, Nakano K (2002) Determination of superoxide by nitrite ion method. Hort Res 1:279–282 (in Japanese)
Madsen JD, Chambers PA, James WF, Koch EW, Westlake DF (2001) The interaction between water movement, sediment dynamics and submersed macrophytes. Hydrobiologia 444:71–84
Nakai S, Inoue Y, Hosomi M, Murakami A (2000) Growth inhibition of blue–green algae (Microcystis aeruginasa) by Myriophyllum spicatum-releasing four polyphenols. Water Res 34:3026–3032
Peutherta A, Pflugmacher S (2010) Influence of the cyanotoxin microcystin-LR on tocopherol in Alfalfa seedlings (Medicago sativa). Toxicon 56:411–417
Pflugmacher S (2004) Promotion of oxidative stress in the aquatic macrophyte Ceratophyllum demersum during biotransformation of the cyanobacterial toxin microcystin-LR. Aquat Toxicol 70:169–178
Qu WC, Mike D, Wang SM (2001) Multivariate analysis of heavy metal and nutrient concentrations in sediments of Taihu Lake, China. Hydrobiologia 450:83–89
Wang C, Zhang SH, Wang PF, Qian J, Hou J, Zhang WJ, Lu J (2009) Excess Zn alters the nutrient uptake and induces the antioxidative responses in submerged plant Hydrilla verticillata (L.f.) Royle. Chemosphere 76:938–945
Xing W, Huang WM, Liu GH (2010) Effect of excess iron and copper on physiology of aquatic plant Spirodela polyrrhiza (L.) Schleid. Environ Toxicol 25:103–112
Ye C, Yu HC, Kong HN, Song XF, Zou GY, Xu QJ, Liu J (2009) Community collocation of four submerged macrophytes on two kinds of sediments in Lake Taihu, China. Ecol Eng 35:1656–1663
Yin Y, Wang XR, Sun YY, Guo HY, Yin DQ (2008) Bioaccumulation and oxidative stress in submerged macrophyte Ceratophyllum demersum L. upon exposure to pyrene. Environ Toxicol 23:328–336
Yu HC, Ye C, Song XF, Liu J (2010) Comparative analysis of growth and physio-biochemical responses of Hydrilla verticillata to different sediments in freshwater microcosms. Ecol Eng 36:1285–1289
Zhang Z, Rengel Z, Meney K (2008) Interactive of N and P on growth but not on resource allocation of Canna indicain wetland microcosms. Aquat Bot 89:317–323
Zhang M, Wang ZQ, Xu J, Liu YQ, Ni LY, Cao T, Xie P (2011) Ammonium, microcystins, and hypoxia of blooms in eutrophic water cause oxidative stress and C–N imbalance in submersed and floating-leaved aquatic plants in Lake Taihu, China. Chemosphere 82:329–339
Zhu GW (2008) Eutrophic status and causing factors for a large, shallow and subtropical Lake Taihu, China. J Lake Sci 20:21–26 (in Chinese)
Acknowledgments
This research was supported by the Research Institute for East Asia Environments, Kyushu University, and financed by Mitsubishi Corporation in Japan.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
Kang, C., Kuba, T., Hao, A. et al. Antioxidant Responses of Vallisneria asiatica to Eutrophic Sediments in Lake Taihu, China. Bull Environ Contam Toxicol 95, 194–199 (2015). https://doi.org/10.1007/s00128-015-1579-4
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00128-015-1579-4