
Abstract
The liver has a central role in the regulation of systemic glucose and lipid fluxes during feeding and fasting and also relies on these substrates for its own energy needs. These parallel requirements are met by coordinated control of carbohydrate and lipid fluxes into and out of the Krebs cycle, which is highly tuned to nutrient availability and heavily regulated by insulin and glucagon. During progression of type 2 diabetes, hepatic carbohydrate and lipid biosynthesis fluxes become elevated, thus contributing to hyperglycaemia and hypertriacylglycerolaemia. Over this interval there are also significant fluctuations in hepatic energy state. To date, it is not known to what extent abnormal glucose and lipid fluxes are causally linked to altered energy states. Recent evidence that the glucose-lowering effects of metformin appear to be mediated by attenuation of hepatic energy generation places an additional spotlight on the interdependence of hepatic biosynthetic and oxidative fluxes. The transition from fasting to feeding results in a significant re-direction of hepatic glucose and lipid fluxes and may also incur a temporary hepatic energy deficit. At present, it is not known to what extent these variables are additionally modified by type 2 diabetes and/or non-alcoholic fatty liver disease. Thus, there is a compelling need to measure fluxes through oxidative, gluconeogenic and lipogenic pathways and determine their relationship with hepatic energy state in both fasting and fed conditions. New magnetic resonance-based technologies allow these variables to be non-invasively studied in animal models and humans. This review summarises a presentation given at the symposium entitled ‘The liver in focus’ at the 2015 annual meeting of the EASD. It is accompanied by two other reviews on topics from this symposium (by Kenneth Cusi, DOI: 10.1007/s00125-016-3952-1, and by Hannele Yki-Järvinen, DOI: 10.1007/s00125-016-3944-1) and a commentary by the Session Chair, Michael Roden (DOI: 10.1007/s00125-016-3911-x).
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Relationship between hepatic glucose, lipid, and energy metabolism
As the major site in the body for carbohydrate and lipid biosynthesis, the liver has a central role in the regulation of systemic glucose and lipid fluxes during feeding and fasting. At the same time, it is also dependent to a considerable degree on the oxidation of these substrates for its own energy needs. Under normal conditions, these activities are highly attuned to carbohydrate and lipid inflow from feeding, and uptake of NEFA derived from adipose tissue lipolysis during fasting. In the setting of type 2 diabetes and non-alcoholic fatty liver disease (NAFLD), control of glucose and lipid biosynthesis is loosened, contributing to excessive systemic glucose and lipid levels. These changes in glucose and lipid fluxes are also accompanied by alterations in hepatocellular energy levels. The benign early stages of NAFLD are associated with an elevated energy state, while the later more severe stages of both NAFLD and type 2 diabetes are associated with a depressed energy state [1]. A key question is: how are the changes in hepatic energy state related to those of hepatic glucose and lipid metabolic fluxes?
Hepatic Krebs cycle as the central link for carbohydrate, lipid and energy metabolism
The liver has the capacity to sustain high rates of both carbohydrate and lipid biosynthesis. In addition to diverting carbons from energy production, these pathways also consume energy and reducing equivalents. The hepatic energy balance is therefore in part determined by the partitioning of carbons between carbohydrate and lipid biosynthesis pathways and oxidation. Under fasting conditions, the profiles of hepatic oxidative and biosynthetic fluxes are relatively well understood. In essence, there is a high flux through fatty acid β-oxidation, sustained by the inflow of NEFA from peripheral adipose tissue lipolysis, with the acetyl-CoA product being fully oxidised to CO2 by the Krebs cycle. This process generates an abundance of ATP and reducing equivalents that sustain the conversion of pyruvate and other anaplerotic substrates into glucose via gluconeogenesis. Under conditions where acetyl-CoA generation from β-oxidation exceeds the oxidative capacity of the Krebs cycle, the acetyl-CoA overflow is diverted into ketone body synthesis. Based on the known rates of hepatic fatty acid oxidation, oxidative Krebs cycle fluxes and gluconeogenesis, and considering that the overwhelming fraction of acetyl-CoA is derived from β-oxidation, the overall process has a positive energy balance. Moreover, the carbon flows are conceptually straightforward, with the catabolic inflow of fatty acid carbons into the Krebs cycle driving the gluconeogenic outflow of anaplerotic carbons. This metabolic model has been confirmed in both fasted humans and animal models in many different laboratories [2–9]. In the setting of mild hepatic insulin resistance, it explains how elevated rates of fasting glucose production are energetically sustainable through systematic increases in both anaplerotic and oxidative Krebs cycle fluxes [5, 10].
If the generation of mitochondrial ATP and reducing equivalents is restricted, then gluconeogenesis from pyruvate is also restrained. Thus, the antihyperglycaemic actions of metformin have been explained through blockade of respiration redox reactions via inhibition of mitochondrial glycerol-3-phosphate dehydrogenase [11] and by direct inhibition of complex I and ATP synthase [12]. Compounds or formulations that reduce energy generation through mild mitochondrial uncoupling have also been shown to improve glucose intolerance and reverse steatosis [13, 14] and have been proposed as a new class of glucose-lowering medications. In this setting, uncoupling agents have an additional advantage over respiration blockers in that they also accelerate fatty acid oxidation, thereby promoting the clearance of excess fat as well as reducing gluconeogenesis.
Measuring hepatic Krebs cycle fluxes under basal fasting conditions
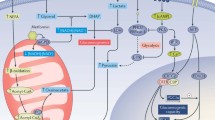
The hepatic Krebs cycle is of high importance in both the manifestation of abnormal hepatic glucose and lipid fluxes and as a target for therapeutic interventions. To date, hepatic Krebs cycle fluxes have been assayed by measuring the flow and distribution of carbon-labelled tracers through its constituent metabolite pools and into biosynthetic products such as glucose. Initial studies using 14C tracers and laborious metabolite carbon-by-carbon degradation methods for obtaining positional labelling information were fundamental in describing the principal carbon flows through the hepatic Krebs cycle [15–17] and provided the foundations for developing stable isotope 13C tracers whose distributions can be analysed more conveniently and in greater detail by MS and NMR methods. While positron emission tomography (PET) has also been used to measure hepatic fatty acid oxidation in humans using [11C]palmitate [18], it does not provide sufficient labelling information for de-convoluting oxidative and anaplerotic Krebs cycle fluxes. With 13C-enriched substrates, the required positional labelling information can be non-invasively acquired by two complementary approaches. In vivo 13C NMR spectroscopy permits direct real-time assessment of positional 13C-enrichment of hepatic Krebs cycle carbon skeletons through the observation of hepatic glutamate and glutamine, whose carbon skeletons are exchanged with that of α-ketoglutarate [19–21]. Ex vivo analysis involves 13C NMR or gas chromatography-mass spectrometry (GC-MS) measurements of circulating or excreted metabolites that can be traced back to the hepatic Krebs cycle (Fig. 1). These include plasma glucose, hepatic uridine diphosphate glucose (UDP-glucose) and hepatic glutamine—the latter two being non-invasively sampled in urine following conjugation with paracetamol and phenylacetic acid, respectively [3, 5, 8, 10, 22–25]. To date, only a very small number of studies describing hepatic Krebs cycle fluxes in individuals with type 2 diabetes or NAFLD have been published. In NAFLD patients with elevated levels of intrahepatic triacylglycerol but relatively mild hepatic insulin resistance, ex vivo 13C-isotopomer and 2H-enrichment analysis of plasma glucose following administration of [3,4-13C2]glucose, [U-13C]propionate and 2H2O revealed systematically higher rates of hepatic oxidative Krebs cycle and gluconeogenic fluxes compared with individuals with normal hepatic triacylglycerol levels [5]. These findings were recapitulated in mouse models of high-fat feeding and hepatic insulin resistance [10]. In overnight-fasted individuals with type 2 diabetes who had moderately increased rates of fasting endogenous glucose production over healthy controls, no significant differences in oxidative and anaplerotic Krebs cycle fluxes were found [3]. More such studies are required to better understand the evolution of hepatic Krebs cycle fluxes with NAFLD and type 2 diabetes pathogenesis and to investigate how they are modulated by medications that target mitochondrial energy generation, such as metformin. Stable isotope tracer methodologies can in principle be integrated with in vivo 31P magnetic resonance measurements of hepatic high-energy phosphate metabolites [26] to precisely determine the relationship between hepatic energy state and Krebs cycle fluxes.

Coverage of the principal catabolic, anaplerotic and biosynthetic fluxes of hepatic carbohydrate and lipid metabolism using deuterated water (2H2O), shown in blue, and 13C-enriched substrates, shown in red. OAA, oxaloacetate; -P, phosphate; PEP, phosphoenolpyruvate
Hepatic carbohydrate, lipid and Krebs cycle fluxes in the fed state
The fed state presents a more complex and perhaps more challenging setting for hepatic control of glucose, lipid and energy metabolism. Under these conditions, the liver receives a range of different nutrients via the portal vein, including simple sugars, amino acids and short-chain fatty acids. Some, such as fructose, glycerol, butyrate and propionate, are efficiently extracted and rapidly incorporated into hepatic intermediary metabolism. Others, such as glucose and lactate, are only partially cleared from the circulation. The abundance of these substrates in portal vein blood, in conjunction with high levels of insulin resulting from beta cell stimulation, promote hepatic glycogen synthesis and de novo lipogenesis, while fatty acid oxidation and endogenous glucose production are suppressed [27]. Ex vivo analysis of plasma glucose, hepatic lipid, and hepatic glycogen deuterium (2H) enrichment from deuterated water (2H2O) provides information on their fractional synthesis or appearance rates and can also provide details of the metabolic carbon sources that sustain these activities (Fig. 1). In rodent models, these analyses reveal that gluconeogenesis contributes a significant fraction of both plasma glucose and hepatic glycogen appearance under natural feeding conditions [28, 29] or following a glucose tolerance test [30]. When the rodent feed was supplemented with sucrose in the drinking water, glucose and glycogen synthesis via anaplerotic fluxes were largely replaced by synthesis from fructose [31]. Analysis of hepatic triacylglycerol or plasma VLDL enrichment from 2H2O [32] as well as 13C-enriched precursors provides information on fractional de novo lipogenesis [33–35], as well as fractional elongation and desaturation rates of the fatty acid moiety and turnover of the glycerol moiety [32, 36]. In animal models of diet-induced NAFLD and insulin resistance, these activities are highly dependent on the constituents of the diet. Fractional lipogenic rates are promoted by diets high in sugar [37] but suppressed by diets high in fat [36, 38]. In humans, elevated lipogenic rates were found in insulin-resistant NAFLD patients [39, 40], as well as in healthy individuals following both acute intake as well as longer term consumption of diets made hyperenergetic with excess fructose [41–43].
Collectively, glycogenic and lipogenic biosynthetic activities consume a significant proportion of mitochondrially generated ATP and reducing equivalents. In addition, ATP can be sequestered during the initial metabolic processing of nutrients, most notably by fructose. At the same time, NEFA availability is reduced, thereby curtailing hepatocyte energy production from NEFA oxidation. Under these conditions, pyruvate dehydrogenase (PDH) is activated via its dephosphorylation, thereby promoting acetyl-CoA generation from glycolytic substrates. The relatively low levels of mitochondrial acetyl-CoA also attenuate pyruvate carboxylase (PC) activity, hence, the ratio of PDH:PC flux—which is very low during fasting—is increased. Once incorporated into citrate, acetyl-CoA may be oxidised or made available for de novo lipogenesis via the citrate shuttle. Despite the central role of these fluxes in rebalancing hepatic lipid, carbohydrate and energy fluxes during the transition from fasting to feeding, very few in situ measurements of Krebs cycle fluxes have been performed under fed conditions in either animal models or humans. Alves et al measured fractional PDH flux contributions to hepatic Krebs cycle fluxes in rats under both fasting state and under hyperinsulinaemic–hyperglycaemic clamp conditions, where the availability of glucose as a substrate is maximal [44]. Their studies confirmed the very low PDH contribution to Krebs cycle fluxes during fasting. While the PDH contribution was significantly increased under the clamp conditions, it accounted for less than half of oxidative Krebs cycle flux, and in rats fed high-fat diets, this contribution fell to about 20% [44]. Magnusson et al measured Krebs cycle fluxes in overnight-fasted healthy volunteers infused with glucose and glucagon [2]. They found that PDH accounted for 40–60% of oxidative Krebs cycle flux under these conditions compared with ∼5% in 60-h-fasted individuals.
Given that, under fed conditions, glycolytic substrates appear to contribute at most ∼50% of the acetyl-CoA requirements for oxidation and that NEFA oxidation is efficiently repressed by malonyl-CoA, are there alternative substrates that could contribute to mitochondrial energy generation? Short-chain fatty acids (SCFAs) are derived from intestinal fermentation of complex carbohydrates and their mitochondrial uptake is independent of carnitine palmitoyltransferase 1a (CPT-1a) and therefore unaffected by cytosolic malonyl-CoA levels. It is not known to what extent the liver is dependent on these or other substrates as alternative energy sources during feeding. Acetate, typically the most abundant intestinally generated SCFA, can also be converted to acetyl-CoA in the cytosol via acetyl-CoA synthase [45, 46], thereby potentially competing for de novo lipogenesis against mitochondrially generated acetyl-CoA. Recent studies suggest that altered intestinal SCFA production is an important link between intestinal microbiome dysbiosis and hepatic insulin resistance and NAFLD [47, 48].
Conclusions
The onset and progression of type 2 diabetes/NAFLD is associated with a progressive impairment in hepatic control of glucose and lipid fluxes. Recent studies also suggest that these changes are also accompanied by alterations in hepatocellular energy state. These alterations in carbon flows and energy status are linked by the hepatic Krebs cycle. To date, there have been very few measurements of human hepatic Krebs cycle fluxes in type 2 diabetes/NAFLD and none have examined the relationship between these variables and hepatic energy state. The transition from fasting to feeding also transforms carbohydrate and lipid carbon fluxes through the Krebs cycle and, depending on the nutrient composition, the hepatic energy state can be acutely compromised, even in healthy individuals. Currently, we have no knowledge of hepatic Krebs cycle fluxes in this setting for either healthy or type 2 diabetic/NAFLD individuals. Finally, the actions of glucose-lowering medications such as metformin appear to be based on compromising the capacity for hepatocellular mitochondrial energy generation. How these effects interplay with acute hepatic energy depletion induced by nutrients such as fructose, as well as underlying impairments of hepatic mitochondrial function in type 2 diabetes/NAFLD, are not well understood. The development of non-invasive tracer approaches that provide comprehensive coverage of hepatic glucose, lipid and Krebs cycle fluxes coupled with non-invasive measurements of hepatocellular energy state should improve our understanding of hepatic glucose and lipid metabolism in healthy humans and better explain the metabolic derangements associated with diabetes and NAFLD.
Abbreviations
- NAFLD:
-
Non-alcoholic fatty liver disease
- PC:
-
Pyruvate carboxylase
- PDH:
-
Pyruvate dehydrogenase
- SCFA:
-
Short-chain fatty acid
References
Koliaki C, Roden M (2013) Hepatic energy metabolism in human diabetes mellitus, obesity and non-alcoholic fatty liver disease. Mol Cell Endocrinol 379:35–42
Magnusson I, Schumann WC, Bartsch GE et al (1991) Noninvasive tracing of Krebs cycle metabolism in liver. J Biol Chem 266:6975–6984
Diraison F, Large V, Brunengraber H, Beylot M (1998) Non-invasive tracing of liver intermediary metabolism in normal subjects and in moderately hyperglycaemic NIDDM subjects. Evidence against increased gluconeogenesis and hepatic fatty acid oxidation in NIDDM. Diabetologia 41:212–220
Large V, Brunengraber H, Odeon M, Beylot M (1997) Use of labeling pattern of liver glutamate to calculate rates of citric acid cycle and gluconeogenesis. Am J Physiol 272:E51–E58
Sunny NE, Parks EJ, Browning JD, Burgess SC (2011) Excessive hepatic mitochondrial TCA cycle and gluconeogenesis in humans with nonalcoholic fatty liver disease. Cell Metab 14:804–810
Jin ES, Burgess SC, Merritt ME, Sherry AD, Malloy CR (2005) Differing mechanisms of hepatic glucose overproduction in triiodothyronine-treated rats vs Zucker diabetic fatty rats by NMR analysis of plasma glucose. Am J Physiol Endocrinol Metab 288:E654–E662
Burgess SC, Jeffrey FMH, Storey C et al (2005) Effect of murine strain on metabolic pathways of glucose production after brief or prolonged fasting. Am J Physiol Endocrinol Metab 289:E53–E61
Jones JG, Solomon MA, Cole SM, Sherry AD, Malloy CR (2001) An integrated 2H and 13C NMR study of gluconeogenesis and TCA cycle flux in humans. Am J Physiol Endocrinol Metab 281:E848–E851
Jones JG, Solomon MA, Sherry AD, Jeffrey FMH, Malloy CR (1998) 13C NMR measurements of human gluconeogenic fluxes after ingestion of [U-13C]propionate, phenylacetate, and acetaminophen. Am J Physiol Endocrinol Metab 275:E843–E852
Satapati S, Sunny NE, Kucejova B et al (2012) Elevated TCA cycle function in the pathology of diet-induced hepatic insulin resistance and fatty liver. J Lipid Res 53:1080–1092
Madiraju AK, Erion DM, Rahimi Y et al (2014) Metformin suppresses gluconeogenesis by inhibiting mitochondrial glycerophosphate dehydrogenase. Nature 510:542–546
Bridges HR, Jones AJY, Pollak MN, Hirst J (2014) Effects of metformin and other biguanides on oxidative phosphorylation in mitochondria. Biochem J 462:475–487
Tao H, Zhang Y, Zeng X, Shulman GI, Jin S (2014) Niclosamide ethanolamine-induced mild mitochondrial uncoupling improves diabetic symptoms in mice. Nat Med 20:1263–1269
Perry RJ, Zhang D, Zhang X-M, Boyer JL, Shulman GI (2015) Controlled-release mitochondrial protonophore reverses diabetes and steatohepatitis in rats. Science 347:1253–1256
Landau BR, Schumann WC, Chandramouli V, Magnusson I, Kumaran K, Wahren J (1993) 14C-labeled propionate metabolism in vivo and estimates of hepatic gluconeogenesis relative to Krebs cycle flux. Am J Physiol 265:E636–E647
Esenmo E, Chandramouli V, Schumann WC, Kumaran K, Wahren J, Landau BR (1992) Use of 14CO2 in estimating rates of hepatic gluconeogenesis. Am J Physiol 263:E36–E41
Schumann WC, Magnusson I, Chandramouli V, Kumaran K, Wahren J, Landau BR (1991) Metabolism of [2-14C]acetate and its use in assessing hepatic Krebs cycle activity and gluconeogenesis. J Biol Chem 266:6985–6990
Iozzo P, Bucci M, Roivainen A et al (2010) Fatty acid metabolism in the liver, measured by positron emission tomography, is increased in obese individuals. Gastroenterology 139:846–856
Vogt JA, Yarmush DM, Yu YM et al (1997) TCA cycle flux estimates from NMR- and GC-MS-determined 13C glutamate isotopomers in liver. Am J Phys 272:C2049–C2062
Jucker BM, Lee JY, Shulman RG (1998) In vivo 13C NMR measurements of hepatocellular tricarboxylic acid cycle flux. J Biol Chem 273:12187–12194
Befroy DE, Perry RJ, Jain N et al (2014) Direct assessment of hepatic mitochondrial oxidative and anaplerotic fluxes in humans using dynamic 13C magnetic resonance spectroscopy. Nat Med 20:98–102
Diraison F, Large V, Maugeais C, Krempf M, Beylot M (1999) Noninvasive tracing of human liver metabolism: comparison of phenylacetate and apoB-100 to sample glutamine. Am J Physiol Endocrinol Metab 277:E529–E536
Yang DW, Previs SF, Fernandez CA et al (1996) Noninvasive probing of citric acid cycle intermediates in primate liver with phenylacetylglutamine. Am J Physiol Endocrinol Metab 270:E882–E889
Weis BC, Margolis D, Burgess SC et al (2004) Glucose production pathways by 2H and 13C NMR in patients with HIV-associated lipoatrophy. Magn Reson Med 51:649–654
Sunny NE, Kalavalapalli S, Bril F et al (2015) Cross-talk between branched-chain amino acids and hepatic mitochondria is compromised in nonalcoholic fatty liver disease. Am J Physiol Endocrinol Metab 309:E311–E319
Szendroedi J, Chmelik M, Schmid AI et al (2009) Abnormal hepatic energy homeostasis in type 2 diabetes. Hepatology 50:1079–1086
Randle PJ (1998) Regulatory interactions between lipids and carbohydrates: the glucose fatty acid cycle after 35 years. Diabetes Metab Rev 14:263–283
Soares AF, Viega FJ, Carvalho RA, Jones JG (2009) Quantifying hepatic glycogen synthesis by direct and indirect pathways in rats under normal ad libitum feeding conditions. Magn Reson Med 61:1–5
Soares AF, Carvalho RA, Veiga FJ et al (2012) Restoration of direct pathway glycogen synthesis flux in the STZ-diabetes rat model by insulin administration. Am J Physiol Endocrinol Metab 303:E875–E885
Delgado TC, Barosa C, Nunes PM, Cerdan S, Geraldes CFGC, Jones JG (2012) Resolving the sources of plasma glucose excursions following a glucose tolerance test in the rat with deuterated water and [U-13C]glucose. PLoS ONE 7, e34042
Delgado TC, Martins FO, Carvalho F et al (2013) 2H enrichment distribution of hepatic glycogen from 2H2O reveals the contribution of dietary fructose to glycogen synthesis. Am J Physiol Endocrinol Metab 304:E384–E391
Lee JJ, Lambert JE, Hovhannisyan Y et al (2015) Palmitoleic acid is elevated in fatty liver disease and reflects hepatic lipogenesis. Am J Clin Nutr 101:34–43
Diraison F, Pachiaudi C, Beylot M (1997) Measuring lipogenesis and cholesterol synthesis in humans with deuterated water: use of simple gas chromatographic mass spectrometric techniques. J Mass Spectrom 32:81–86
Diraison F, Pachiaudi C, Beylot M (1996) In vivo measurement of plasma cholesterol and fatty acid synthesis with deuterated water: determination of the average number of deuterium atoms incorporated. Metab: Clin Exp 45:817–821
Parks EJ, Hellerstein MK (2006) Recent advances in liver triacylglycerol and fatty acid metabolism using stable isotope labeling techniques. J Lipid Res 47:1651–1660
Duarte JAG, Carvalho F, Pearson M et al (2014) A high-fat diet suppresses de novo lipogenesis and desaturation but not elongation and triglyceride synthesis in mice. J Lipid Res 55:2541–2553
Martins F, Delgado TC, Viegas J et al (2015) Mechanisms by which the thiazolidinedione troglitazone protects against sucrose-induced hepatic fat accumulation and hyperinsulinemia. Br J Pharmacol 173:267–278
Delgado TC, Pinheiro D, Caldeira M et al (2009) Sources of hepatic triglyceride accumulation during high-fat feeding in the healthy rat. NMR Biomed 22:310–317
Donnelly KL, Smith CI, Schwarzenberg SJ, Jessurun J, Boldt MD, Parks EJ (2005) Sources of fatty acids stored in liver and secreted via lipoproteins in patients with nonalcoholic fatty liver disease. J Clin Investig 115:1343–1351
Diraison F, Moulin P, Beylot M (2003) Contribution of hepatic de novo lipogenesis and reesterification of plasma non esterified fatty acids to plasma triglyceride synthesis during non-alcoholic fatty liver disease. Diabetes Metab 29:478–485
Parks EJ, Skokan LE, Timlin MT, Dingfelder CS (2008) Dietary sugars stimulate fatty acid synthesis in adults. J Nutr 138:1039–1046
Faeh D, Minehira K, Schwarz JM, Periasami R, Seongsu P, Tappy L (2005) Effect of fructose overfeeding and fish oil administration on hepatic de novo lipogenesis and insulin sensitivity in healthy men. Diabetes 54:1907–1913
Schwarz J-M, Noworolski SM, Wen MJ et al (2015) Effect of a high-fructose weight-maintaining diet on lipogenesis and liver fat. J Clin Endocrinol Metab 100:2434–2442
Alves TC, Befroy DE, Kibbey RG et al (2011) Regulation of hepatic fat and glucose oxidation in rats with lipid-induced hepatic insulin resistance. Hepatology 53:1175–1181
Knowles SE, Jarrett IG, Filsell OH, Ballard FJ (1974) Production and utilization of acetate in mammals. Biochem J 142:401–411
Sone H, Shimano H, Sakakura Y et al (2002) Acetyl-coenzyme A synthetase is a lipogenic enzyme controlled by SREBP-1 and energy status. Am J Physiol Endocrinol Metab 282:E222–E230
Vos MB (2014) Nutrition, nonalcoholic fatty liver disease and the microbiome: recent progress in the field. Curr Opin Lipidol 25:61–66
De Vadder F, Kovatcheva-Datchary P, Goncalves D et al (2014) Microbiota-generated metabolites promote metabolic benefits via gut-brain neural circuits. Cell 156:84–96
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Funding
The author receives financial support from Portuguese Foundation for Science and Technology research grant EXCL/DTP/0069/2012. Also, structural funding for the Center for Neurosciences and UC-NMR facility is supported in part by FEDER – European Regional Development Fund through the COMPETE Programme and the Portuguese Foundation for Science and Technology through grants PEst-C/SAU/LA0001/2011; REEQ/481/QUI/2006, RECI/QEQ-QFI/0168/2012, CENTRO-07-CT62-FEDER-002012, and Rede Nacional de Ressonância Magnética Nuclear.
Duality of interest
The author declares that there is no duality of interest associated with this manuscript.
Contribution statement
The author is the sole contributor to this paper.
Rights and permissions
About this article

Cite this article
Jones, J.G. Hepatic glucose and lipid metabolism. Diabetologia 59, 1098–1103 (2016). https://doi.org/10.1007/s00125-016-3940-5
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00125-016-3940-5