
Abstract
Aims/hypothesis
Increased renal mast cells have been detected in diabetic nephropathy. However, only a few patients have been examined. Evidence of the involvement of mast cells in diabetic nephropathy is still scarce, and no observation of mast cells during the development of diabetic nephropathy has yet been reported in humans. Here, we examined changes in renal mast cells in patients at different stages of diabetic nephropathy and related these to the development of the disease.
Methods
Eighty patients at different clinical stages of diabetic nephropathy and 16 normal kidney donors were recruited. Immunohistochemical staining for tryptase, chymase, TGF-β1, renin and TNF-α was done on renal sections from patients and control participants. Changes in mast cell number, degranulation, subtype and phenotype were examined. Correlation between mast cells and patients’ clinical and pathological indices was analysed.
Results
With progression of diabetic nephropathy, the number and degranulation level of mast cells increased. Increase in mast cell number and degranulation level correlated significantly with tubular interstitial injury. Almost all renal mast cells in patients with diabetic nephropathy were found to produce chymase, renin, TGF-β1 and TNF-α. The level of TNF-α in mast cells increased with progression of diabetic nephropathy.
Conclusions/interpretation
This study suggests that mast cells are involved in development of diabetic nephropathy. Through release of bioactive substances, such as tryptase, chymase, TGF-β1, renin and TNF-α, into the tubular interstitium by degranulation, mast cells could promote renal inflammation and fibrosis, and thus contribute to diabetic nephropathy.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Diabetic nephropathy is a multistage clinical syndrome characterised by thickening of the glomerular basement membrane and mesangial expansion with progression into glomerulosclerosis, tubular necrosis and interstitial fibrosis, which ultimately result in renal failure. Although a lot of investigations have been carried out, the pathogenic mechanism involved in diabetic nephropathy is not completely understood and no satisfactory therapeutic option is available to treat this insidious disease [1].
Mast cells (MCs) are a group of multifunctional cells. Knowledge obtained in the past two decades indicates that MCs are not only the key effector cells in anaphylaxis, but also regulators of innate and adaptive immune responses [2, 3]. The pathogenic roles of MCs have been extended to allergic diseases and helminth infection, and other diseases such as autoimmune diseases, inflammatory diseases, carcinogenesis and fibrosis [3–10]. With regards to the production of tryptase and chymase, two subtypes of MCs have been reported in humans: the MCT subtype is believed to produce tryptase only, whereas the MCTC subtype is believed to produce both tryptase and chymase.
MCs have been detected in a lot of renal diseases [11–22]; however, only a few cases were investigated in each kind of renal disease in most of the studies reported. Observation of MCs in the development of renal disease is still scarce. Evidence provided by previous studies suggests that MCs contribute to renal diseases mainly by promoting tubular interstitial injury; however, their exact role remains poorly understood. Increased numbers of MCs have been reported in diabetic nephropathy [11, 13, 17], but, again, only a few patients were examined. Evidence of the involvement of MCs in diabetic nephropathy is still scarce, and no observation has been made of MCs during the development of diabetic nephropathy in humans. The aim of the present study was to examine changes in MC number, degranulation level, subtype and phenotype in the renal tissues of patients at different stages of diabetic nephropathy and to relate this to the development of disease.
Methods
Study population
Eighty type 2 diabetic patients with biopsy-proven diabetic nephropathy, who were hospitalised at the clinical unit of the nephrology centre of Jingling Hospital from 2003 to 2008, were recruited. All these patients met the World Health Organization criteria for type 2 diabetes mellitus, and renal biopsy consistent with the diagnosis of diabetic nephropathy and exclusion for other concomitant renal diseases. According to their clinical characteristics (regardless of duration of diabetes), patients were classified into microalbuminuria (urinary albumin excretion rate [UAE] >30 mg/24 h and <300 mg/24 h, n = 19), proteinuria (UAE ≥300 mg/24 h and estimated glomerular filtration rate [eGFR, according to the equation of Modification of Diet in Renal Disease] ≥60 ml min−1 1.73 m−2, n = 32) and renal insufficiency (eGFR <60 ml min−1 1.73 m−2, n = 29) stage groups. Patients with hypertension were given angiotensin-receptor blockers such as irbesartan or valsartan. Other antihypertensive agents (calcium-channel blockers, ACE inhibitors, diuretics, etc.) were also used as needed. The target blood pressure was the same (a systolic blood pressure of 135 mmHg or less, and a diastolic blood pressure of 85 mmHg or less). Sixteen normal renal tissues from mismatched kidney donors served as normal control participants (normal control group). Informed consent was obtained from participants. All research work with human participants was in accordance with the ethical standards of the responsible committee on human experimentation and with the Declaration of Helsinki. Table 1 shows relevant clinical and pathological characteristics of the patients.
Histology
For light microscopy, renal biopsy specimens were fixed in 10% neutral buffered formalin, embedded in paraffin and sectioned at 2 μm thickness. Haematoxylin–eosin, periodic acid–Schiff’s reagent, Masson’s trichrome and periodic acid methenamine silver staining were carried out. Glomerular, tubular interstitial and vascular lesions in biopsies were recorded, classified and scored according to criteria described by Tervaert et al. [23]. Immunofluorescence included the use of antibodies against IgA, IgG, IgM, C3, C1q, and κ and λ light chains to rule out other renal diseases. Electron microscopy was also performed to confirm the diagnosis.
Neutrophils, eosinophils and plasma cells were identified according to the morphological characteristics of these cells on haematoxylin and eosin-stained sections, and the number of these cells in the tubular interstitium was counted by two pathologists. Immunohistochemical staining for CD3, CD20 and CD68 was performed to label T lymphocytes, B lymphocytes and macrophages respectively. The numbers of CD3, CD20 and CD68 positive cells in the tubular interstitium were counted. For each section, at least ten randomly selected cortical regions were examined, and the area of the selected regions in each section was measured using NIS Element BR3.4 software (Nikon, Shinagaw-ku, Tokyo, Japan). The number of each kind of inflammatory cell (including neutrophils, eosinophils, plasma cells, T lymphocytes, B lymphocytes and macrophages) was presented as the number of the cells per mm2 of renal cortex area.
The relative interstitial volume (RIV) was measured according to methods described by Okon et al. [17, 24]. For each section, ten randomly selected cortical regions were measured and the average value was taken as the RIV of the section.
Evaluation of the degranulation level of MCs
Renal sections were stained with Toluidine Blue and examined under a high power field by two different researchers using a single blind approach. Degranulated MCs were determined by the presence of extruded granule contents [25]. At least 20 randomly selected fields were examined for each section. The total numbers of Toluidine Blue staining-positive cells (total MCs) and of degranulated MCs were counted. The degranulation level of MCs in each section was presented as the ratio of degranulated MCs to total MCs.
Immunohistochemistry
Immunohistochemical staining was performed using routine methods. Briefly, formalin-fixed sections were deparaffinised with xylene and rehydrated with graded ethanols. The sections were autoclaved for antigen repair. Endogenous peroxidase was blocked with hydrogen peroxide. For reduction of background labelling, the sections were blocked for 30 min in 10% fetal calf serum. The samples were rinsed in PBS and then incubated overnight at 4°C with different first antibodies, such as mouse anti-human MC tryptase monoclonal antibody (Chemicon, Temicon, CA, USA, 1:2,000), mouse anti-human MC chymase monoclonal antibody (Chemicon, 1:1,000), rabbit anti-human renin polyclonal antibody (Santa Cruz Biotechnology, CA, USA, 1:200), mouse anti-human TNF-α monoclonal antibody (Santa Cruz Biotechnology, 1:100), rabbit anti-human TGF-β1 polyclonal antibody (Santa Cruz Biotechnology, 1:200), mouse anti-human CD3 monoclonal antibody (Novocastra, UK, 1:200), mouse anti-human CD20 monoclonal antibody (Dako, Glostrup, Denmark, 1:100) and mouse anti-human CD68 monoclonal antibody (Dako, 1:100). After washing with PBS, the tissues were incubated with Envision reagent (Dako) for 45 min, then the sections were washed and developed with diaminobenzidine. Each section was counterstained with haematoxylin. Normal homologous serum was used to replace the first antibody as a negative control.
Measurement of renal MCs
Immunohistochemical staining for tryptase was carried out to mark all the renal MCs. The cortex part of each renal section was fully scanned and photographed under a light microscope by a CCD (Nikon DS-Ri1, Nikon), which was connected to a personal computer. Then, the number of tryptase-positive MCs in the cortex of each section was counted by two independent researchers using a single blind approach. The renal tissue area of the same region was measured using NIS Element BR software. The number of renal MCs in each section was presented as the number of MCs per mm2 of renal cortex area (MCs density).
Analysis of the subtype of MCs
Immunohistochemical staining for tryptase and chymase was carried out on serial renal sections. The MCs that were positive for tryptase staining and negative for chymase staining were believed to be MCT subtype. The MCs that were positive for both tryptase and chymase staining were believed to be MCTC subtype. To measure the ratio of each subtype, at least ten randomly selected high power fields were examined. The number of each subtype of MC was counted and the ratio was calculated for each section.
Semi-quantitative analysis of TNF-α in renal MCs
The TNF-α level in MCs was analysed quantitatively using Image-pro Plus 6.0 software (Media Cybernetics, Bethesda, MD, USA). Serial renal sections were used in the assay. For each pair of serial sections, one section was stained for tryptase to mark MCs, and the other was stained for TNF-α. At least 15 randomly selected MCs were analysed from each section of the diabetic nephropathy patients. As only a few MCs were found in each section of the normal control participants, all MCs found in all sections were pooled together and analysed.
Statistical analysis
All data were analysed using SPSS version 10.0 (SPSS, Chicago, IL, USA). Because the data involved a small number of participants (for each group) or were determined to be non-normal, all the data are presented as median (interquartile range), and multiple comparisons were performed using Kruskal–Wallis ANOVA, with pairwise comparison performed using Mann–Whitney U test with Bonferroni correction. Correlation analysis was carried out using Spearman coefficients with Bonferroni correction to adjust for the multiple testing. p < 0.05 was considered statistically significant.
Results
Distribution of MCs in renal tissues of patients with diabetic nephropathy
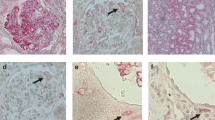
MCs were detected in the cortex and medulla of renal sections from diabetic nephropathy patients by immunohistochemical staining for tryptase, which is the specific molecular marker of MCs. Most MCs were found to be distributed diffusely in the interstitium, especially in the peritubular, perivascular and periglomerular regions (Fig. 1b). Occasionally, the MCs were found to be infiltrated in the wall of atrophied tubules. Few intraglomerular MCs were detected in our tissue samples. MCs were mostly located on the periphery rather than within areas where inflammatory cells aggregated.

Distribution of MCs in the renal tissues of patients with diabetic nephropathy. MCs (shown by black arrows) were found to be distributed diffusely in the interstitium. a Negative control of immunohistochemistry (original magnification ×200). Scale bar, 100 μm. b Occasionally, the MCs were found to be infiltrated in the wall of atrophied tubules (original magnification ×200). Scale bar, 100 μm. c Immunohistochemical staining for tryptase was used to mark the MCs. To show the renal structure better, haematoxylin–eosin staining was performed after immunohistochemical staining (original magnification ×400). Scale bar, 50 μm
Renal MCs increased as the disease progressed
In the normal control group, only a few MCs were found. The density of MCs in the normal control group was 2.2 (0.3–3.7) MCs/mm2. However, increased numbers of MCs were found in the diabetic nephropathy patients and these paralleled the progression of the disease (Fig. 2).

Renal MCs (shown by black arrows) increased with development of diabetic nephropathy. Sections of renal biopsies from 80 patients with diabetic nephropathy (including the microalbuminuria stage group [n = 19], proteinuria stage group [n = 32] and renal insufficiency stage group [n = 29]) and 16 normal control participants were used in the assay. Immunohistochemical staining for tryptase was carried out to mark renal MCs. The number of tryptase-positive cells in the cortex part of each section was counted. The area of the same part was measured and the density of MCs in renal cortex was calculated. Multiple comparisons were performed using Kruskal–Wallis ANOVA, and pairwise comparison was performed using Mann–Whitney U test, with Bonferroni correction. a Normal control group. b Microalbuminuria stage group. c Proteinuria stage group. d Renal insufficiency stage group. Scale bars for (a–d) 100 μm. e Results of statistical analysis, **p < 0.01. White circles represent extreme values where the extremes are defined as cases with values more than three box lengths from the upper or lower edge of the box. The box length is the interquartile range. MG, microalbuminuria stage group; NG, normal control group; PG, proteinuria stage group; RIG, renal insufficiency stage group
Degranulation level of MCs increased with the development of diabetic nephropathy
The degranulation level of MCs was analysed using Toluidine Blue staining. In the normal control group, degranulation was found in only a few MCs (about one in ten). However, with the development of diabetic nephropathy, the degranulation level of MCs increased significantly (Fig. 3).

Increased degranulation level of renal MCs (shown by black arrows) was found with development of diabetic nephropathy. Sections of renal biopsies from 80 patients with diabetic nephropathy (including microalbuminuria stage group [n = 19], proteinuria stage group [n = 32] and renal insufficiency stage group [n = 29]) and 16 normal control participants were used in the assay. For the diabetic nephropathy patients, at least 20 randomly selected high power fields were examined and the ratio of degranulated MCs was calculated for each section. As only a few MCs were found in each section from the normal controls, all MCs found in all sections from the normal controls were pooled together and the ratio of degranulated MCs was calculated (three degranulated MCs were found in 31 MCs). Multiple comparisons were performed using Kruskal–Wallis ANOVA, and pairwise comparison was performed using Mann–Whitney U test, with Bonferroni correction. a MCs without degranulation (note that the MC is intact and no granule was found outside the cell). b Degranulated MCs (note that some granules have been released outside the cell). Scale bars for (a) and (b) 10 μm. c Results of statistical analysis: the horizontal line was represents the average degranulation level of the normal control group, **p < 0.01. White circles represent extreme values where the extremes are defined as cases with values more than three box lengths from the upper or lower edge of the box. The box length is the interquartile range. MG, microalbuminuria stage group; NG, normal control group; PG, proteinuria stage group; RIG, renal insufficiency stage group
Correlation of renal MCs with clinical and pathological indices
As shown in Table 2, density of renal MCs was found to be correlated significantly with eGFR, serum creatinine, serum cystatin C, urine retinol-binding protein (RBP), urine lysozyme, urine N-acetyl-β-glucosaminidase (NAG), urine C3, urine α2 microglobulin and 24 h urine protein level. Significant correlation was also found between the density of renal MCs and the RIV. Moreover, MCs were found to be significantly correlated with the number of renal tubular interstitial inflammatory cells, including neutrophils, eosinophils, macrophages and T lymphocytes. However, plasma cells and B lymphocytes were not found to be correlated with MCs.
Most MCs were found to be MCTC subtype and no change in MC subtype was found during the development of diabetic nephropathy
To explore whether MCs at different stages of diabetic nephropathy have different subtypes and functions, production of tryptase and chymase in the renal MCs was investigated. Immunohistochemical results showed that more than 80% of the tryptase-positive MCs in the sections of diabetic nephropathy patients and normal control participants were chymase-positive (Fig. 4). No obvious change was found in subtype of MCs in different groups of diabetic nephropathy patients.

Most of the tryptase-positive renal MCs (shown by black arrows) of normal controls and patients with diabetic nephropathy were found to be chymase-positive. Serial renal sections from 30 diabetic nephropathy patients (including microalbuminuria stage group [n = 10], proteinuria stage group [n = 10] and renal insufficiency stage group [n = 10]) and ten normal controls were used in the assay. For the diabetic nephropathy patients, the percentage of MCTC MCs was calculated for each section. As only a few MCs were found in each section from the normal controls, all the MCs found in all sections from the normal controls were pooled together and analysed. Multiple comparisons were performed using Kruskal–Wallis ANOVA, and pairwise comparison was performed using Mann–Whitney U test, with Bonferroni correction. a Normal control group, immunohistochemically stained for tryptase. b Normal control group, immunohistochemically stained for chymase. c Microalbuminuria stage group, immunohistochemically stained for tryptase. d Microalbuminuria stage group, immunohistochemically stained for chymase. e Proteinuria stage group, immunohistochemically stained for tryptase. f Proteinuria stage group, immunohistochemically stained for chymase. g Renal insufficiency stage group, immunohistochemically stained for tryptase. h Renal insufficiency stage group, immunohistochemically stained for chymase. i Negative control of immunohistochemistry. Scale bars (a–i) 100 μm. j Results of statistical analysis: the horizontal line represents the average ratio of MCTC MCs in the normal control group. White circles represent extreme values where the extremes are defined as cases with values more than three box lengths from the upper or lower edge of the box. The box length is the interquartile range. a and b, c and d, e and f, g and h are serial sections. MG, microalbuminuria stage group; NG, normal control group; PG, proteinuria stage group; RIG, renal insufficiency stage group
Expression changes of TGF-β1, renin and TNF-α in renal MCs during the development of diabetic nephropathy
TGF-β1 and renin were detected in the MCs of normal control participants and diabetic nephropathy patients. No obvious change in these two molecules was found in the MCs of the diabetic nephropathy patients compared with the normal control participants (Fig. 5). Only very weak immunohistochemical staining for TNF-α was found in the MCs of normal controls; however, immunohistochemical staining for TNF-α increased in the MCs of diabetic nephropathy patients. The immunohistochemical staining intensity of TNF-α increased gradually from the microalbuminuria stage to the proteinuria stage and renal insufficiency stage (Fig. 6).

TGF-β1 and renin were detected in MCs (shown by black arrows) of normal controls and patients with diabetic nephropathy. No obvious change in these two molecules was found in the MCs of diabetic nephropathy patients compared with those of the normal controls. a Normal controls, immunohistochemically stained for tryptase. b Normal controls, immunohistochemically stained for renin. c Patients with diabetic nephropathy, immunohistochemically stained for tryptase. d Patients with diabetic nephropathy, immunohistochemically stained for renin. e Normal controls, immunohistochemically stained for tryptase. f Normal controls, immunohistochemically stained for TGF-β1. g Patients with diabetic nephropathy, immunohistochemically stained for tryptase. h Patients with diabetic nephropathy, immunohistochemically stained for TGF-β1. i Negative control of immunohistochemistry. Scale bars (a–i) 50 μm. a and b, c and d, e and f, g and h are serial sections

TNF-α in renal MCs (shown by black arrows) was found to be increased with the development of diabetic nephropathy. Serial sections of renal biopsies from 30 patients with diabetic nephropathy (including microalbuminuria stage group [n = 10], proteinuria stage group [n = 10] and renal insufficiency stage group [n = 10]) and ten normal controls were used in the assay. At least 15 randomly selected MCs were analysed in each section of diabetic nephropathy patients. As only a few MCs were found in each section from the normal controls, all the MCs found in all sections from the normal controls were pooled together and analysed (in total, 21 MCs were found and analysed). Multiple comparisons were performed using Kruskal–Wallis ANOVA, and pairwise comparison was performed using Mann–Whitney U test, with Bonferroni correction. a Normal control group, immunohistochemically stained for tryptase. b Normal control group, immunohistochemically stained for TNF-α. c Microalbuminuria stage group, immunohistochemically stained for tryptase. d Microalbuminuria stage group, immunohistochemically stained for TNF-α. e Proteinuria stage group, immunohistochemically stained for tryptase. f Proteinuria stage group, immunohistochemically stained for TNF-α. g Renal insufficiency stage group, immunohistochemically stained for tryptase. h Renal insufficiency stage group, immunohistochemically stained for TNF-α. Scale bars (a–h) 50 μm. i Results of quantitative analysis, **p < 0.01. a and b, c and d, e and f, g and h are serial sections. MG, microalbuminuria stage group; NG, normal control group; PG, proteinuria stage group; RIG, renal insufficiency stage group
Discussion
By using renal sections from diabetic nephropathy patients at different stages of disease, the present study investigated changes in MCs occurring during development of the disease and the association between renal MCs and the progression of diabetic nephropathy. Increased renal MCs were found at all stages of diabetic nephropathy and these paralleled the progression of the disease. As early as the microalbuminuria stage, MCs began to increase. As diabetic nephropathy progressed, renal MCs increased continually. Interestingly, in the present study, the degranulation level of MCs was also found to be increased with the development of diabetic nephropathy. MCs are rich in growth factors, proteases and inflammatory mediators. Once activated, MCs can release various kinds of pathogenic substances such as TGF-β, chymase, tryptase, histamine, renin and TNF-α by degranulation [26–31]. Certainly, release of these kinds of substances can cause renal injury. Therefore, the increase in the number and degranulation level of MCs suggests that MCs could be involved in the development of diabetic nephropathy. In accordance with this, correlation analysis revealed that MCs correlated significantly with serum creatinine, serum cystatin C and the decline of GFR.
Previous studies suggested that MCs contributed to renal diseases mainly through promotion of tubular interstitial injury [13, 15–17, 32–34]. Most investigations focused on the roles of MCs on renal fibrosis. Similar to previous studies [16], in the present study, MCs were found to be distributed mainly in the tubular interstitium, but rarely in the glomerulus. Furthermore, the present study showed that the increase in MC number and degranulation level occurred mainly at the later stages of diabetic nephropathy (in the proteinuria and renal insufficiency stage groups) when tubular interstitial injury became obvious. The involvement of MCs in tubular interstitial injury was also supported by the results of correlation analysis. MCs were found to be correlated significantly with urine RBP, urine lysozyme, urine NAG and RIV. In addition, renal MCs correlated significantly with the tubular interstitial inflammatory cells, including neutrophils, eosinophils, macrophages and T lymphocytes. The correlation between MCs and inflammatory cells suggests that MCs play a role in promoting renal inflammation in diabetic nephropathy.
MCs can be activated in several ways. The well-known, classical pathway of MC activation is through IgE–FcεR cross-linking. But MCs can also be activated through alternative pathways, such as the complement pathway and the microbial pattern recognition receptors (toll-like receptors) pathway. In vitro studies have demonstrated that C3a is the most potent chemoattractant and activator of MCs [35, 36]. Activation of complement, which can lead to the generation of C3a, has also been demonstrated to increase in diabetic nephropathy [37]. Thus, we suggest that increased complement activation might contribute to the recruitment and activation of MCs.
Tryptase and chymase are two major proteinases in MCs. Studies have shown that these two enzymes play very important roles in the function of MCs [21]. According to the production of the two molecules, human MCs are classified into two subtypes [16]. The MCTC MCs are believed to produce both tryptase and chymase, whereas the MCT MCs are believed to produce tryptase only. The MCT MCs are believed to be T cell-dependent and the MCTC MCs are believed to be T cell-independent [38]. In human renal tissue, different results have been reported on MC subtype [11, 15, 17, 32–34]. MC subtype is believed to be disease-dependent [16]. In diabetic nephropathy, Rüger et al. reported that most MCs of diabetic nephropathy patients (about 80.3%) contained both tryptase and chymase [11]. However, in another study, Okon and Stachura reported a much lower ratio of chymase-positive MCs in diabetic nephropathy patients (about 52.9%) [17]. In the present study, approximately 84% of the tryptase-positive MCs in diabetic nephropathy patients were found to be chymase-positive, and no obvious change in subtype was found in different groups of diabetic nephropathy patients. This result supports the idea that MCTC is the major subtype of renal MCs in diabetic nephropathy patients. Unexpectedly, in the present study, 80.5% of the MCs in normal control participants were found to be MCTC subtype. The result was quite different from what had been reported by Okon and Stachura [17] and Tóth et al. [33] (about 14.5% and 20.1% of MCs, respectively, in normal control participants were found to be MCTC subtype in their reports), but similar to the result of Rüger et al. [11] (about 62.5% of MCs in normal controls were found to be MCTC subtype in their report). Is MCTC the major subtype of renal MCs in healthy human beings? As only a few MCs could be found in each section of the normal control participants, investigations based on many more specimens are required.
In the present study, we also investigated the expression of TGF-β1, renin and TNF-α in MCs. As an important profibrotic factor, TGF-β has been proven to play important roles in the pathogenesis of diabetic nephropathy [39–42]. As the key enzyme in the process of angiotensin production, renin participates in the regulation of extracellular volume, arterial vasoconstriction and blood pressure. Also, it has been reported that renin can upregulate TGF-β expression [43, 44]. Recently, renin was reported to be able to induce renal inflammation through binding and activating prorenin receptors [45]. An increased renin level was reported in patients with diabetic nephropathy and treatment with renin inhibitor attenuated the development of the disease [46]. TNF-α is a pleiotropic cytokine. As an important proinflammatory molecule, TNF-α could promote the expression of adhesion molecules and the recruitment and activation of inflammatory cells [47, 48]. In addition, TNF-α could increase production of reactive oxygen species [49]. For the first time, we demonstrated that TGF-β, renin and TNF-α are produced by human renal MCs. Interestingly, the level of TNF-α in MCs was found to be increased with the development of diabetic nephropathy. These findings further support the involvement of MCs in diabetic nephropathy, and also provide us with some clues for deeper understanding of the mechanisms through which MCs could exert their roles: (a) through synthesising and releasing TGF-β and renin, MCs may contribute to tubular interstitial fibrosis; (b) through synthesising and releasing renin and TNF-α, MCs may initiate and (or) promote tubular interstitial inflammation. These clues are helpful not only for understanding the mechanisms of diabetic nephropathy, but also for developing treatment strategies. For example, as a novel renin inhibitor, aliskiren may protect against diabetic nephropathy not only through blocking the renin–angiotensin system, but also through inhibiting MC degranulation-associated renal inflammation.
It should be pointed out that in the present study we could not identify the resident MCs and the infiltrated MCs. Thus, we could not differentiate their pathogenic roles in the development of diabetic nephropathy. MCs are highly heterogeneous. The microenvironment (for example the presence of different kinds of cytokines) in which the MCs reside determines the gene expression and phenotype development of the cells. With the variation of microenvironmental conditions, the phenotypes (for example, the expression level of TNF-α, as shown in the present study) and pathogenic roles of MCs may differ to some degree during the development of diabetic nephropathy. The present study did not differentiate the pathogenic roles of MCs at microalbuminuria stage, macroalbuminuria stage and renal insufficiency stage. Also, the present study did not observe the influence of agents that antagonise the renin–angiotensin system on MC number. In a study based on a 5/6 nephrectomy rat model, Jones et al. reported that ACE inhibition led to a reduction in renal MC number and reduced renal injury [50]. As a possible new mechanism of renin–angiotensin system blockage therapy, it is interesting. Further exploration of these questions may provide helpful information for better understanding of the mechanisms of diabetic nephropathy and the pathogenic roles of MCs.
In summary, increased MCs and degranulation level were found at all stages of diabetic nephropathy and these paralleled the development of the disease. Increased MCs correlated significantly with the indices indicating tubular and interstitial injury. As MCs are multifunctional, the mechanisms of MC involvement in diabetic nephropathy are likely to be complex. However, as MCs have been proven to contain tryptase, chymase, TGF-β, renin and TNF-α, theoretically they can contribute to diabetic nephropathy through synthesis and release of these molecules. Taken together, results from the present study suggest that MCs play important roles in the development of diabetic nephropathy, especially in tubular interstitial injury. However, further work is necessary to prove this.
Abbreviations
- eGFR:
-
Estimated glomerular filtration rate
- MC:
-
Mast cell
- NAG:
-
N-acetyl-β-glucosaminidase
- RBP:
-
Retinol-binding protein
- RIV:
-
Relative interstitial volume
- UAE:
-
Urinary albumin excretion rate
References
Balakumar P, Arora MK, Ganti SS, Reddy J, Singh M (2009) Recent advances in pharmacotherapy for diabetic nephropathy: current perspectives and future directions. Pharmacol Res 60:24–32
Kawakami T (2009) A crucial door to the mast cell mystery knocked in. J Immunol 18:6863–6881
Galli SJ, Kalesnikoff J, Grimbaldeston MA, Piliponsky AM, Williams CM, Tsai M (2005) Mast cells as “tunable” effector and immunoregulatory cells: recent advances. Annu Rev Immunol 23:749–786
Dawicki W, Marshall JS (2007) New and emerging roles for mast cells in host defence. Curr Opin Immunol 19:31–38
Sayed BA, Christy A, Quirion MR, Brown MA (2008) The master switch: the role of mast cells in autoimmunity and tolerance. Annu Rev Immunol 26:705–739
Malaviya R, Abraham SN (2001) Mast cell modulation of immune responses to bacteria. Immunol Rev 179:16–24
Galli SJ, Nakae S, Tsai M (2005) Mast cells in the development of adaptive immune responses. Nat Immunol 6:135–142
Levick SP, McLarty JL, Murray DB, Freeman RM, Carver WE, Brower GL (2009) Cardiac mast cells mediate left ventricular fibrosis in the hypertensive rat heart. Hypertension 53:1041–1047
Sakamoto-Ihara T, Suzuki Y, Kurusu A, Yamashita M, Horikoshi S, Tomino Y (2007) Possible involvement of mast cells in renal fibrosis in patients with IgA nephropathy. Inflamm Res 56:421–427
Melillo RM, Guarino V, Avilla E et al (2010) Mast cells have a protumorigenic role in human thyroid cancer. Oncogene 29:6203–6215
Rüger BM, Hasan Q, Greenhill NS, Davis PF, Dunbar PR, Neale TJ (1996) Mast cells and type VIII collagen in human diabetic nephropathy. Diabetologia 39:1215–1222
Hiromura K, Kurosawa M, Yano S, Naruse T (1998) Tubulointerstitial mast cell infiltration in glomerulonephritis. Am J Kidney Dis 32:593–599
Roberts IS, Brenchley PE (2000) Mast cells: the forgotten cells of renal fibrosis. J Clin Pathol 53:858–862
Kondo S, Kagami S, Kido H, Strutz F, Müller GA, Kuroda Y (2001) Role of mast cell tryptase in renal interstitial fibrosis. J Am Soc Nephrol 12:1668–1676
Goto E, Honjo S, Yamashita H, Shomori K, Adachi H, Ito H (2002) Mast cells in human allografted kidney: correlation with interstitial fibrosis. Clin Transplant 16(Suppl 8):7–11
Ehara T, Shigematsu H (2003) Mast cells in the kidney. Nephrology (Carlton) 8:130–138
Okon K, Stachura J (2007) Increased mast cell density in renal interstitium is correlated with relative interstitial volume, serum creatinine and urea especially in diabetic nephropathy but also in primary glomerulonephritis. Pol J Pathol 58:193–197
Holdsworth SR, Summers SA (2008) Role of mast cells in progressive renal diseases. J Am Soc Nephrol 19:2254–2261
Welker P, Krämer S, Groneberg DA et al (2008) Increased mast cell number in human hypertensive nephropathy. Am J Physiol Renal Physiol 295:F1103–F1109
Silva GE, Costa RS, Ravinal RC, dos Reis MA, Dantas M, Coimbra TM (2008) Mast cells, TGF-beta1 and alpha-SMA expression in IgA nephropathy. Dis Markers 24:181–190
Balakumar P, Reddy J, Singh M (2009) Do resident renal mast cells play a role in the pathogenesis of diabetic nephropathy? Mol Cell Biochem 330:187–192
Togawa H, Nakanishi K, Shima Y et al (2009) Increased chymase-positive mast cells in children with crescentic glomerulonephritis. Pediatr Nephrol 24:1071–1075
Tervaert TW, Mooyaart AL, Amann K et al (2010) Pathologic classification of diabetic nephropathy. J Am Soc Nephrol 21:556–563
Okon K, Szumera A, Kuzniewski M (2003) Are CD34+ cells found in renal interstitial fibrosis? Am J Nephrol 23:409–414
Singh LK, Boucher W, Pang X et al (1999) Potent mast cell degranulation and vascular permeability triggered by urocortin through activation of corticotropin-releasing hormone receptors. J Pharmacol Exp Ther 288:1349–1156
Metz M, Grimbaldeston MA, Nakae S, Piliponsky AM, Tsai M, Galli SJ (2007) Mast cells in the promotion and limitation of chronic inflammation. Immunol Rev 217:304–328
Nakae S, Suto H, Berry GJ, Galli SJ (2007) Mast cell-derived TNF can promote Th17 cell-dependent neutrophil recruitment in ovalbumin-challenged OTII mice. Blood 109:3640–3648
Grimbaldeston MA, Nakae S, Kalesnikoff J, Tsai M, Galli SJ (2007) Mast cell-derived interleukin 10 limits skin pathology in contact dermatitis and chronic irradiation with ultraviolet B. Nat Immunol 8:1095–1104
Moon TC, St Laurent CD, Morris KE et al (2010) Advances in mast cell biology: new understanding of heterogeneity and function. Mucosal Immunol 3:111–128
Silver RB, Reid AC, Mackins CJ et al (2004) Mast cells: a unique source of renin. Proc Natl Acad Sci USA 101:13607–13612
Helske S, Syväranta S, Kupari M et al (2006) Possible role for mast cell-derived cathepsin G in the adverse remodelling of stenotic aortic valves. Eur Heart J 27:1495–1504
Ehara T, Shigematsu H (1998) Contribution of mast cells to the tubulointerstitial lesions in IgA nephritis. Kidney Int 54:1675–1683
Tóth T, Tóth-Jakatics R, Jimi S, Takebayashi S (2000) Increased density of interstitial mast cells in amyloid A renal amyloidosis. Mod Pathol 13:1020–1028
Yamada M, Ueda M, Naruko T et al (2001) Mast cell chymase expression and mast cell phenotypes in human rejected kidneys. Kidney Int 59:1374–1381
Nilsson G, Johnell M, Hammer CH et al (1996) C3a and C5a are chemotaxins for human mast cells and act through distinct receptors via a pertussis toxin-sensitive signal transduction pathway. J Immunol 157:1693–1698
Hartmann K, Henz BM, Krüger-Krasagakes S et al (1997) C3a and C5a stimulate chemotaxis of human mast cells. Blood 89:2863–2870
Hansen TK, Forsblom C, Saraheimo M et al (2010) Association between mannose-binding lectin, high-sensitivity C-reactive protein and the progression of diabetic nephropathy in type 1 diabetes. Diabetologia 53:1517–1524
Irani AM, Craig SS, DeBlois G, Elson CO, Schechter NM, Schwartz LB (1987) Deficiency of the tryptase positive, chymase negative mast cell type in gastrointestinal mucosa of patients with defective T lymphocyte function. J Immunol 138:4381–4386
Chen S, Jim B, Ziyadeh FN (2003) Diabetic nephropathy and transforming growth factor-beta: transforming our view of glomerulosclerosis and fibrosis build-up. Semin Nephrol 23:532–543
Border WA, Noble NA (1997) TGF-beta in kidney fibrosis: a target for gene therapy. Kidney Int 51:1388–1396
Border WA, Noble NA (1998) Evidence that TGF-beta should be a therapeutic target in diabetic nephropathy. Kidney Int 54:1390–1391
Sharma K, Jin Y, Guo J, Ziyadeh FN (1996) Neutralization of TGF-beta by anti-TGF-beta antibody attenuates kidney hypertrophy and the enhanced extracellular matrix gene expression in STZ-induced diabetic mice. Diabetes 45:522–530
Huang Y, Noble NA, Zhang J, Xu C, Border WA (2007) Renin-stimulated TGF-beta1 expression is regulated by a mitogen-activated protein kinase in mesangial cells. Kidney Int 72:45–52
He M, Zhang L, Shao Y et al (2009) Inhibition of renin/prorenin receptor attenuated mesangial cell proliferation and reduced associated fibrotic factor release. Eur J Pharmacol 606:155–161
Balakumar P, Jagadeesh G (2010) Cardiovascular and renal pathologic implications of prorenin, renin, and the (pro)renin receptor: promising young players from the old renin-angiotensin-aldosterone system. J Cardiovasc Pharmacol 56:570–579
Kelly DJ, Zhang Y, Moe G, Naik G, Gilbert RE (2007) Aliskiren, a novel renin inhibitor, is renoprotective in a model of advanced diabetic nephropathy in rats. Diabetologia 50:2398–2404
Holtmann MH, Neurath MF (2004) Differential TNF-signaling in chronic inflammatory disorders. Curr Mol Med 4:439–444
Matera MG, Calzetta L, Cazzola M (2010) TNF-alpha inhibitors in asthma and COPD: we must not throw the baby out with the bath water. Pulm Pharmacol Ther 23:121–128
Koike N, Takamura T, Kaneko S (2007) Induction of reactive oxygen species from isolated rat glomeruli by protein kinase C activation and TNF-alpha stimulation, and effects of a phosphodiesterase inhibitor. Life Sci 80:1721–1728
Jones SE, Kelly DJ, Cox AJ, Zhang Y, Gow RM, Gilbert RE (2003) Mast cell infiltration and chemokine expression in progressive renal disease. Kidney Int 64:906–913
Acknowledgements
This study was supported by the Jingling Hospital Fund (2010 M027). We thank J. P. Wang and C. H. Zen, who are affiliated to the Research Institute of Nephrology, Jingling Hospital, for their help with histological analysis. We thank X. D. Zhu and M. C. Zhang, who are affiliated to the Research Institute of Nephrology, Jingling Hospital, for their help with immunohistochemical analysis. We also thank Y. X. Liu and M. L. Zhou, who are affiliated to the Data and Statistics Centre of Jingling Hospital for their help with data analysis.
Contribution statement
JMZ conceived and designed the study, drafted the article, participated in the revision of the manuscript and gave final approval of the version to be published. GHY, ZC, RW and ZHL participated in the conception of the study, the analysis and interpretation of data and revising the article, and gave final approval of the version to be published.
Duality of interest
The authors declare that there is no duality of interest associated with this manuscript.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Zheng, J.M., Yao, G.H., Cheng, Z. et al. Pathogenic role of mast cells in the development of diabetic nephropathy: a study of patients at different stages of the disease. Diabetologia 55, 801–811 (2012). https://doi.org/10.1007/s00125-011-2391-2
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00125-011-2391-2