
Abstract
Soybean [Glycine max (L.) Merr.] was one of the most important legume crops in the world in 2010. Japanese beetles (JB; Popillia japonica, Newman) in the US were an introduced and potentially damaging insect pest for soybean. JBs are likely to spread across the US if global warming occurs. Resistance to JB in soybean was previously reported only in plant introductions. The aims here were to identify loci underlying resistance to JB herbivory in recombinant inbred lines (RILs) derived from the cross of Essex × Forrest cultivars (EF94) and to correlate those with loci with factors that confer insect resistance in soybean cultivars. The RIL population was used to map 413 markers, 238 satellite markers and 177 other DNA markers. Field data were from two environments over 2 years. Pest severity (PS) measured defoliation on a 0–9 scale. Pest incidence (PI) was the percentage of plants within each RIL with beetles on them. Antibiosis and antixenosis data were from feeding assays with detached leaves in petri plates. Five QTL were detected for the mean PS field trait (16% < R 2 < 27%). The loci were within the intervals Satt632–A2D8 on linkage group (LG) A2 (chromosome 8); Satt583–Satt415 on LG B1 (11); Satt009–Satt530 on LG N (3); and close to two markers OB02_140 (LG E; 20 cM from Satt572) and OZ15_150 LG (19 cM from Satt291 C2). Two QTL were detected for the mean PI field trait (16% < R 2 < 18%) close to Satt385 on LG A1 and Satt440 on LG I. The no choice feeding studies detected three QTL that were significant; two for antixenosis (22% < R 2 < 24%) between Satt632–A2D8 on LG A2 (8) and Sat_039–Satt160 on LG F (13); and a major locus effect (R 2 = 54%) for antibiosis on LG D2 (17) between Satt464–Satt488. Therefore, loci underlying resistance to JB herbivory were a mixture of major and minor gene effects. Some loci were within regions underlying resistance to soybean cyst nematode (LGs A2 and I) and root knot nematode (LG F) but not other major loci underlying resistance to nematode or insect pests (LGs G, H and M).
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Japanese beetle (JB; Popillia japonica, Newman) is of the Order Coleoptera and Family Scarabaeidae (Potter and Held 2002). The Japanese beetle is native to the main island of Japan where it is not a damaging pest. However, in the US and Canada, JBs represent an introduced scarab that causes losses for both crops and ornamental plants. It was first found in the US in 1916 at a nursery near Riverton, New Jersey. The beetle is currently found in coastal and adjacent midwest states from Maine to Alabama with small infestations westward beyond the Mississippi River. Three infestations of this pest have been eradicated from California and several Western state remained JB free by 2009.
Japanese beetles are voracious feeders with over 300 host plant species (Ladd 1987, 1989). Feeding typically removes all interveinal cells leaving a network. The chemicals released from plant materials by feeding are attractants to other JBs so swarms develop on particular plants. Feeding attractants include simple sugars and volatiles with a fruity or flowery smell such as terpenoids, aliphatics, and aromatics (Loughrin et al. 1996). Feeding deterrents found in non-host plants appear to be anti-nutrients that reduce longevity and fecundity, since JBs do not avoid or learn to avoid known toxins. Feeding peaks from early morning to mid-afternoon with females leaving the swarms in mid-afternoon to oviposit on the roots or feed in solitary.
The larval stages, commonly called white grubs, were especially damaging to roots (Potter and Held 2002). The larvae feed on roots for a year, overwintering there. They pupate in Spring and emerge as adults in early summer. Adult females alternately feed, mate with multiple partners and return to the soil to oviposit so that populations of JBs can increase dramatically in 1 year following infestation. The extensive intermating suggests JBs have evolved under positive selection for increased recombination and the resultant diversity (Ladd 1987).
JBs threaten sustainable soybean–corn rotation production in the US as both crops are favored hosts. However, JBs may avoid fields close to non-host species such as sorghum (Smith et al. 1988; Bohlen and Barrett 1990). Further, threat derives from their invasive nature, high rates of inter-mating and tendency of larger swarms to form each year (Yesudas 2007; Tigreros and Switzer 2009). No-till and reduced till productions favor JB grub survival. Therefore, damage may be predicted to increase as farming practices change and global warming accelerates.
By 2009, JBs were often a significant pest of soybean in the Midwest and eastern US (Delucia et al. 2008; Zavala et al. 2009). However, the damage to cost of control threshold was rarely reached because it was predicted to be 30% defoliation before bloom, factoring in the cost of insecticides (Gould 1963). However, the threshold was somewhat arbitrary and did not account for the root damage caused by the white grubs nor the effect on the rotational crop the next year. Indeed, the testing of insecticidal seed treatments has shown major yield effects that cast doubts over previous estimates of the seed yield reductions ascribed to insect pests (Bradshaw et al. 2008).
Innate genetic resistance in soybean to JBs would be a low cost control method. JB-resistant soybean germplasm releases have been developed from a plant introduction (P.I. 171451; Hammond et al. 2001). The discovery was serendipitous in that the germplasm was selected for Mexican bean beetle, but also showed resistance to Japanese beetles in field (Hammond and Cooper 1989). The genetic basis of the resistance that was introgressed from P.I. 171451 is probably located on LG M linked to Satt463, Satt220, and Satt536. The gene underlying this QTL had not been reported by 2010. However, increased insect susceptibility in CO2 enrichment was attributed to decreased cysteine protease inhibitors (Zavala et al. 2009; Casteel et al. 2008).
Forced feeding allows the detection of antibiosis and antixenosis by soybean leaves and the chemicals within them (Rector et al. 2000). Natural products found in soybean, such as coumarin, can be shown to act as deterrents for JBs when added to artificial diets (Patton et al. 1997b). Soybean plants contain many chemicals in roots, shoots and seed that deter the feeding by many insect pests (Narvel et al. 2001). Isoflavones (phaseol, afromosin, coumestrol, daidzein, and glyceollins) and other natural products may confer resistance to soybean (Rector et al. 2000; Patton et al. 1997a). The amount of each chemical required for resistance varies among cultivars and insect pests, and is partly determined by the genotype, developmental stage and organ ingested (Chiari et al. 2004; Kassem 2004, 2006; Primomo et al. 2006; Afzal et al. 2009). Conversely, protein, oil, and sugars were known soybean components that would increase JB fecundity and longevity. However, separating the effects of attractants or nutrients from anti-nutrients or feeding deterrents can be complex.
Studies of insect resistance in soybean have concentrated on plant introduction-derived resistances to corn ear worm, aphids, cutworm and leaf hoppers sometimes combined with Bt-derived resistances (Killen and Lambert 1986; Narvel et al. 2001; Baur and Boethel 2004; Nakazawa 2005). Specific insect pest resistance has been found in P.I. 229358 (‘Sodendaizu’), P.I. 1171451 (‘Kosamame’), and P.I. 227687 (‘Miyako White’). Both open field scores and caged or trapped feeding has been used. In traps, weight gains of insect pests and/or lethality (Komatsu et al. 2005) was an indicator resistance. In the field, feeding preference was evidenced by the extent of feeding (Rector et al. 2000). By 2008, about 30 major insect-resistant QTL had been identified, many provide a broad resistance to several insect pests (Narvel et al. 2001). Broad resistance suggests certain soybean genotypes actively synthesize and accumulate natural products to stop (antixenosis) or deter (antibiosis) insect feeding.
Here, the feeding damage caused by JB was analyzed on a group of 96 recombinant inbred lines (RILs) lacking major gene resistance. Swarming damage was measured as pest severity (PS), cultivar choice was measured as pest incidence (PI). Forced feeding allowed measurement of antibiosis and antixenosis. Genetic analysis with DNA markers allowed loci underlying partial resistance to JBs to be identified.
Materials and methods
Seed material
In 2005, the EF94 RIL seed material (at the F5:15 generation) for this study was acquired from the SIUC seed store from the 2001 increase of the released EF94 population (Lightfoot et al. 2005). Briefly, the population was 94 RILs derived at the F5 by single seed descent and bulked for the nine generations since. Near isogenic lines (NILs) derived from individual RILs were also used. The Essex cultivar parent was partially susceptible with trait means of 1.07 ± 0.12 beetle deaths (MNANTIX) and 0.025 ± 0.002 g weight gain per beetle (MNANTIB) in forced feeding; and 25.5 ± 0.9% MNPI and 3.51 units MNPS in field experiments of cultivar choice. The Forrest cultivar parent was partially resistant with trait means of 1.81 ± 0.12 beetle deaths (MNANTIX), 0.011 ± 0.002 g (MNANTIB), 16.2 ± 0.8% MNPI, 2.62 ± 0.05 units of MNPS. The seeds of the RILs, NILs and parents were grown in 2005 and 2006 at the SIUC, Agronomy Research Center (ARC) fields 19 and 26 at Carbondale, Illinois. Fields were planted to corn the previous year. Field border crops were pasture, sorghum and corn (2 sides) in 2005 and woodland, sorghum and corn (2 sides) in 2006. The 2006 seed planted were those harvested in 2005 (at the F5:16 generation).
Field plots
In 2005, the RILs were planted as four row plots. The rows were 16 lines deep and 36 lines wide and the outer two rows were planted with RIL ExF85, a highly disease susceptible line (Iqbal et al. 2005). The plots were 3 m long and had a 2 m alley way. In 2006, the rows were 24 deep and 48 wide, 2 row plots. Individual plots were 2 m long and had a 1.3 m alley way. Herbicide treatments for the planting year 2005 had a pre- and a post-application. The pre-application was done on 06 June 2005 using Valor (flumioxazin-VALENT-EPA Reg. No. 59639-99 or 59639-98), sprayed at 140 ml/ha (2 oz. per acre) and sprayed for pre-broadleaf and grass suppression in soybean. Prowl (pendimethalin-Dow 19 AgriSci. EPA Reg. No. 68156-62719) was mixed with water at 1.68 l/ha and sprayed for pre-grass. The post-application was done on 08 July 2005. Stellar (flumiclorac + gal lactofen-VALENT-EPA Reg. No. 59639-92) was sprayed at 490 ml/ha for broadleaf and Select AE (Clethodim-VALENT-EPA Reg. No. 59639-3) was used as 560 ml/ha used for annual and perennial grass control. Herbicide treatment for the 2006 planting year also had a pre- and a post-application. The pre-application was done on 01 June 2006 using Valor at 140 ml/ha. Dual Magnum™ (s-metolachlor + benoxacor-SYNGENTA-EPA Reg. No. 100-816) was also sprayed at 1.68 l/ha (1.5 pt. per acre) for pre-grass, annual broadleaf and grass weeds. Touchdown™ (glyphosate diamonium salt-SYNGENTA-EPA Reg. No. 100-1121) was also sprayed at 1.68 l/ha for non-selective burn down before planting. Post-emergence application was done on 17 July 2006 using Stellar at 490 ml/ha and Select at 560 ml/ha.
Seed weight and yield measurements
Seeds were harvested and de-podded using a two row combine. The seeds were then sieved several times to get rid of broken seeds, seed coat debris, pods and weed seeds. Seed weight was measured for 100 seeds and the whole plot by weight.
Insect feeding in fields
The plants were scored for PS, PI, pest number (PN) and pest index (PX). PX was PS × PI. PS was calculated as the scale of damage on the 100 E × F lines (scale 0–9). The lines were damaged from 0, or very little damage, to 9 that were more than 90% defoliated. The PI was calculated as the number of individual plants within a given line that were affected by JBs and the PN was calculated based on the number of JBs that were present per plant during the feeding period. The PS field data were used to select the 30 RIL lines as high (n = 10), mid (n = 10) and low (n = 10) genotype groups. In addition, 20 NIL lines as worst (n = 10) and best (n = 10) were selected for the forced feeding study.
No choice feeding tests for JBs
In 2005 and 2006 two leaflets from each of 30 RILs were taken and placed in a Petri dish with a moistened filter paper disc. Two JBs were weighed and introduced into each of the Petri dishes. The Petri dishes were sealed and left on the bench under fluorescent light at 22°C for a week. The final JB weights were taken as measure of antibiosis (ANTIB) and number of alive and dead JBs noted as antixenosis (ANTIX). This data were used for analyzing feeding preference through statistical analysis systems, analytical methods (SAS 2006). The forced feeding assay data were pooled for the years 2005 and 2006 as a t test showed no significant difference between the 2 years (Snedecor and Cochran 1980).
Data analysis
Following Kassem et al. (2006), the programs used were Mapmaker EXP 3.0, Mapmaker QTL 1.1 (Lander et al. 1987) and QTL Cartographer 1.16 for trait distribution associations. SAS and Microsoft Excel programs were used for heritability estimates and correlations. Pearson’s correlation coefficient analysis was performed on the mean trait data (Snedecor and Cochran 1980) in order to find correlations among traits.
Results
Trait distributions
All traits measured were highly heritable and ranged from 82 to 95% in the broad sense (Table 1). Mean PI and mean ANTIB each showed a nearly normal and continuous distribution. Mean PS and ANTIX were not normal and either biphasic or discontinuous indicating the effect of major loci. The mean value of PS was 3.01 and the standard deviation was 2.15 (Fig. 1). Distributions show positive skewness (0.95) and negative kurtosis (−0.54) resulting in a flattened-peaked distribution (Fig. 1a). Essex was 3.51 ± 0.05 units and Forrest was 2.62 ± 0.05 so there were both positive and negative transgressive segregants (Fig. 2). Mean PI had a mean value of 19.82 and the standard deviation was 4.89. The distribution was positively skewed (1.25) and kurtotic (2.79; Fig. 1b). Essex was 25.5 ± 0.9% and Forrest was 16.2 ± 0.8% so there were both positive and negative transgressive segregants. The mean value of ANTIB was 0.017 g and the standard deviation was 0.005 g. The distribution showed positive skewness (1.69) and kurtosis (2.16) resulting in a peaked distribution (Fig. 1c). Essex was 0.025 ± 0.002 g and Forrest was 0.011 ± 0.002 g so there were both positive and negative transgressive segregants. The mean ANTIX was 1.3 with standard deviation of 0.4. The distribution had just three major categories, 1, 1.5 or 2 dead JBs (Fig. 1d). Essex had 1.07 ± 0.12 and Forrest had 1.81 ± 0.12 beetle deaths so there were both positive and negative transgressive segregants. There were no significant correlations among the mean traits suggesting they were separate measures of the effects of soybean herbivory. Traits did not correlate significantly with seed composition or yield, including seed phytoestrogen, seed protein and seed oil content.

Trait distribution among 100 lines for the JB related mean traits measured in the 100 RIL population of Essex × Forrest. The individual traits were: pest severity (PS); pest incidence (PI); antibiosis (ANTIB) was the weight gain of JBs during no choice feeding; and antixenosis (ANTIX) was the lethality of JBs per RIL. Means (MN) of traits are shown. Values on the y axis are midpoint range values. The Essex cultivar parent means were 1.07 ± 0.12 beetle deaths (MNANTIX), 0.025 ± 0.002 g weight gain per beetle (MNANTIB), 25.5 ± 0.9% MNPI and 3.51 ± 0.05 units MNPS. The Forrest cultivar parent means were 1.81 ± 0.12 beetle deaths (MNANTIX), 0.011 ± 0.002 g (MNANTIB), 16.2 ± 0.8% MNPI, 2.62 ± 0.05 units of MNPS

Japanese beetles on soybean leaflets. a Simultaneous mating and feeding. b Three leaflets with 10% PS rating caused by two JBs in 1 h. c The white grubs that are the larval stages of JBs (taken from Fig. 14. White grubs of (L-R) Japanese beetle, European chafer, May–June beetle. Photo courtesy of David Cappaert, http://www.ext.colostate.edu/pubs/insect/05601.htm). d Leaves with 80% defoliation showing the web of veins left

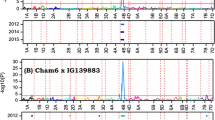
Locations of the QTL found in the EF94 population on linkage groups A2, D2 and F for resistance to JB, in relation to known QTL for resistance to SCN and RKN. JB (grey solid arrow), Hg type 1.2.5- (grey stippled arrows), Hg type 0 (black stippled arrows), Hg type 1.3 (black solid arrow), RKN (white arrow). The size of the arrow reflects the interval significantly associated by QTL Cartographer or Mapmaker at LOD > 2.5 or ANOVA at P < 0.0005
QTL analysis
The 218 BARC-Satt marker data set and linkage groups developed by Kassem et al. (2006) were used to scan for QTL along with AFLP, SCAR and RAPD markers developed earlier (Chang et al. 1997).
PS of JBs
Five QTL were detected for the mean PS field trait (16% < R 2 < 27%; Table 2; Fig. 3). The loci were within the intervals Satt632–A2D8 on linkage group (LG) A2 (chromosome 8); Satt583–Satt415 on LG B1 (11); Satt009–Satt530 on LG N (3); and close to two markers not closely linked to others OB02_140 (LG E) and OZ15_150 (LG C2; Table 3). Beneficial alleles were from Forrest on A2, N and E but from Essex on B1 and C2. Allelic effects ranged from 1.1 to 1.7 units difference in PS.
PI of JBs
Two QTL were detected for the mean PI field trait (16% < R 2 < 18%) close to Satt385 on LG A1 and Satt440 on LG I. Beneficial alleles were from Forrest. Allelic effects ranged from 3.0 to 3.1% reduction in PI. One locus was detected where the beneficial allele was different in the two locations (Table 3). Marker SIUC_B08D14 on LG B1 (11) had Forrest as a beneficial allele in 2005 and Essex in 2006. Therefore, this locus was not significantly associated with the mean trait.
Antixenosis to JBs
The no choice feeding studies detected two QTL that were significant for antixenosis (22% < R 2 < 24%) between Satt632–A2D8 on LG A2 (8) and Sat_039- Satt160 on LG F (13; Table 2; Fig. 3). Beneficial alleles causing more JB deaths were from Forrest. Allelic effects ranged from 0.22 to 0.25 increase in JB mortality.
Antibiosis to JBs
A major locus effect (R 2 = 54%) was detected for antibiosis on LG D2 (17) between Satt464–Satt488 (Table 2; Fig. 3). The beneficial allele causing lower JB weight gains were from Forrest. The allelic effect was a 0.011 g difference in JB weight over the forced feeding period.
Discussion
Analysis in the Rosaceae of QTL for JB resistance suggested that there was a significant environmental dependence but the genetic component was underlain by specific biochemical pathways that produce antixenotic products (Patton et al. 1997b). However, in EF94 soybean JB resistance was a highly heritable genetic trait by all measures (Table 1) suggesting a multi-factored genetic interaction.
PS was a measure of the tendency to swarm on particular genotypes. Several QTL of small effect were detected. Such QTL might underlie the amount or quality of odorants produced during herbivory that serve to attract other JBs to a feeding swarm (Loughrin et al. 1996). Known attractants include simple sugars and volatiles with a fruity or flowery smell such as terpenoids, aliphatics, and aromatics. The five QTL each encompass an interval with genes encoding enzymes in these broad synthetic pathways. Biochemical mechanisms triggered by herbivory and signals initiated during the wounding responses (Patton et al. 1997a, b; de Bruxelles and Roberts 2001; Underwood and Rausher 2002) might underlie the QTL detected in soybean if taste rather than toxicity or nutritive values are altered.
PI was a measure of a tendency toward cultivar choice within the EF94 population. Only two QTL were detected and they were of large effect. One QTL that was detected in single years was not found in the mean of those years because the beneficial allele was opposite across the 2 years, suggesting a G × E interaction at that locus. JBs are not known to choose among host plants on the basis of genotype, being unable to detect or avoid toxins (Ladd 1987, 1989; Loughrin et al. 1996). However, it is possible that since soybeans and JBs have co-evolved in Japan, JBs are able to detect certain chemicals in soybean that serve as attractants or repellents.
Antibiosis showed the JBs grew larger when a single locus on LG D2 was inherited from the Essex parent. Consistent with this the Essex parent was more nutritive for JBs than Forrest. This locus appeared to be a major gene effect and might be underlain by a locus altering leaf composition, of nutrients, anti-nutrients or toxins. QTL in this region include those for resistance to SCN (Schuster et al. 2001; Kazi et al. 2009), SDS (Kazi et al. 2008) Sclerotinia (Arahana et al. 2001) and corn ear worm (Chase et al. 2001). Therefore, genes in this region involved in broad pest resistance might underlie the antinutritive effects recorded for JBs. Alternately, a single locus might underlie resistance to the three insect-like pests JBs, CEW and SCN Hg types 1.2.5- and 1.3-. This locus might be ascribed to a gene symbol Rpj1 with permission from the Soybean Genetics Committee.
Antixenosis showed the JBs had greater mortality rates when two loci on LG 2 and F were inherited from the Forrest parent. Consistent with this the Essex parent caused less death for JBs than Forrest. These loci appeared to be major gene effects and might be underlain by a loci altering leaf toxin content. Loci in the A2 region included Rhg4 for resistance to SCN (Webb et al. 1995; Meksem et al. 2001; Kazi et al. 2009) and a QTL for leaf area (Ashley et al. 1998). The F region encompassed QTL for resistance to root knot nematodes (Boerma et al. 1997). Therefore, genes in these regions involved in nematode resistance might underlie the lethality effects recorded for JBs. These locus might be ascribed to gene symbols Rpj2 and Rpj3 with permission from the Soybean Genetics Committee.
The QTL reported previously for antibiosis and antixenosis to Mexican bean beetle located on LGs M from P.I. 171451 (Narvel et al. 2001) were not detected in JB resistance assays. There was a QTL for PS detected in 2005 on LG M by marker SIUC_B20C11 which might reflect the activity of the same locus. There were very few polymorphic markers on LG M in the EF94 population (Kassem et al. 2006) so another possibility is that the M locus might be fixed. The loci for soybean resistance to herbivory by other insects on LG D1b, G and H were not detected although many markers were polymorphic on those LGs.
Pleiotropy or linkage among QTL may have been detected between Rhg4 Rhg3, Mj and the JB resistance QTL found on LG A2, D2 and F, respectively. Loci underlying resistance to soybean cyst nematode (Rhg genes) are either linked or pleiotropic to genes underlying resistance to sudden death syndrome; at rhg1/Rfs2 (Triwitayakorn et al. 2005); and at rhg3/rfs10 (Kazi et al. 2008, 2009). Proven examples of pleiotropy are rare since they require the isolation in transgenic plants of at least one gene and measurement of two traits. However, there are examples from insect resistances in tomato. The tomato Mi-1 gene confers resistance against root knot nematodes (Meloidogyne spp.), whitefly (Bemisia tabaci; Nombela et al. 2003) and a biotype of the potato aphid (Macrosiphum euphorbiae; Martinez de Ilarduya et al. 2003). The phenyl propanoid pathway leading to maysin has been associated with resistance to multiple insect pests in maize (Abel et al. 2000) that are controlled by alleles at the P1gene (a transcription factor) that was epistatic over the a1 (dihydroflavanol reductase; E.C. 1.1.1.219) and whp (white pollen 1; unknown protein) allelic variations (Szalma et al. 2005). Arabidopsis induces common pathways to insect pests (Mewis et al. 2005). However, linkage is more likely across the majority of clustered QTL. Also, possible is that the extensive epistasis that has been detected in soybean (Chase et al. 2001) due to the homeologous genomes (Shultz et al. 2007) may cause the loci to appear multi-functional.
QTL maps suggested the region for resistance to SCN and Japanese beetle on LG A2 overlapped. Therefore, genes underlying the two QTL might be linked or pleiotropic. Recombination is suppressed in this region due to the I gene associated deletions and the subsequent introgression of Rhg4 into the region encompassed by SIUC–A2D8 and BLT65. The SCN susceptible cultivars ‘Williams 82’ and ‘A3244’ encoded a cluster of nine genes within this region (Lightfoot and Meksem 2001; Kazi et al. 2009; Campbell et al. 2009). The genes linked to the RLK, primary candidate for SCN resistance Rhg4, may be involved in resistance to JBs. The linked set of genes at the Rhg4 locus included an EST (gi 5677126) expressed in leaves, a brassinosteroid-regulated protein (gi 347458) abundant in etiolated hypocotyls of very young seeds, a paralog of EST (gi 12493776) from G. soja abundant in etiolated seedling, a predicted gene of no known expression, a duplicate pair of paralogs of an EST (gi 15662259) that was similar to a UDP-glucose:anthocyanin 5-O-glucosyltransferase, a paralog of an EST (gi15662259), the presumed Rhg4 receptor like kinase abundant in roots, an EST (gi 21256330) similar to a serine protease abundant in roots, and the gene (gi 2970554) encoding aspartokinase-homoserine dehydrogenase (Gebhardt et al. 1998). Each gene may be a candidate to underlie the resistance to JBs. The dual activity of many insect resistance genes would mean that the Rhg4 nematode resistance gene was a strong candidate. However, since nematodes parasitize plant cells by cytoplasm to cytoplasm, or membrane to membrane, contact through a stylet whereas insect pests ingest crushed cells, a common mechanism would infer the involvement of the induction of a common metabolite or small molecule.
The RLK protein at Rhg4 may be implicated in sensing the nematode and setting up a response that includes the induction of the phenylpropanoid and glucosinolate pathways (Mahalingham and Skorupska 1996; Mewis et al. 2005). This response could confer broad insect resistance like the Mi gene (Nombela et al. 2003; Martinez de Ilarduya et al. 2003).The duplicate pair of paralogs of an EST (gi 15662259) that were similar to a UDP-glucose:anthocyanin 5-O-glucosyltransferase could be involved in the production of chemicals toxic to insect pests such as maysin in maize (Abel et al. 2000; Szalma et al. 2005). Aspartokinase-homoserine dehydrogenase is a bifunctional enzyme found in plant chloroplasts that catalyses the first and third steps of methionine and threonine biosynthesis. Both amino acids can be used to produce defense compounds through special nitrogen metabolism (Gebhardt et al. 1998). Equally intriguing, the EST (gi 5677126) expressed in leaves and the defense-related brassinosteroid-regulated protein (gi 347458) abundant in etiolated hypocotyls of very young seeds are expressed in the organ that the JBs feed upon. Further tests of function will use fine mapping in NILs followed by candidate gene TILLING and transformation (Lightfoot 2008).
The development of host plant resistance in a wide range of plant species (Patton et al. 1997a) may cause JBs to increase their feeding on soybean leaves. Global warming and increased CO2 concentrations would increase the nutrition value of soybean leaves to JB populations (Casteel et al. 2008). Better understanding of the pleiotropic effects of genes, QTL interactions (Walker et al. 2004) and their effects on related traits like seed composition to JB herbivory will help develop new varieties with resistance to JB without introgression from PIs (Hammond and Cooper 1989). Pyramiding or stacking of the QTL discovered here can lead to the development of better varieties of soybean cultivars.
References
Abel CA, Wilson RL, Wiseman BR, White WH, Davis FM (2000) Conventional resistance of experimental maize lines to corn earworm (Lepidoptera: Noctuidae), fall armyworm (Lepidoptera: Noctuidae), southwestern corn borer (Lepidoptera: Crambidae), and sugarcane borer (Lepidoptera: Crambidae). J Econ Entomol 93:982–988
Afzal AJ, Natarajan A, Saini N, Iqbal MJ, Geisler MA, El Shemy H, Mungur R, Willmitzer L, Lightfoot DA (2009) The nematode resistance allele at the rhg1 locus alters the proteome and metabolome of soybean roots. Plant Physiol 151:1264–1280
Arahana VS, Eskridge KM, Graef GL, Specht JE, Steadman JR (2001) Identification of QTL for Resistance to Sclerotinia sclerotiorum in Soybean. Crop Sci 41:180–188
Ashley DA, Boerma HR, Carter TE Jr, Mian MAR, Wells R (1998) An additional QTL for water use efficiency in soybean. Crop Sci 38:390–393
Baur ME, Boethel D (2004) Host plant resistance in IPM and the advances and restraints on Bt engineered soybean/crops. In: Proceedings of the VII world soybean research conference; IV international soybean processing and utilization conference; III Congresso Mundial de Soja (Brazilian soybean congress)
Boerma HR, Hussey RS, Luzzi BM, Parrott WA, Tamulonis JP (1997) RFLP mapping of resistance to southern root-knot nematode in soybean. Crop Sci 37:1903–1909
Bohlen PJ, Barrett GW (1990) Dispersal of the Japanese beetle (Coleoptera: Scarabaeidae) in strip-cropped soybean agroecosystems. Environ Entomol 19:955–960
Bradshaw JD, Rice ME, Hill JH (2008) Evaluation of management strategies for bean leaf beetles (Coleoptera: Chrysomelidae) and bean pod mottle virus (Comoviridae) in soybean. J Econ Entomol 101:1211–1227
Campbell N, Warner AL, Lightfoot DA, Matthews BF, Knap HT (2009) Duplication of a chromosomal region from linkage group A2 involved in cyst nematode resistance in soybean. Mol Gen Genet (in press)
Casteel CL, O’Neill BF, Zavala JA, Bilgin DD, Berenbaum MR, Delucia EH (2008) Transcriptional profiling reveals elevated CO2 and elevated O3 alter resistance of soybean (Glycine max) to Japanese beetles (Popillia japonica). Plant Cell Environ 31:419–434
Chang SJC, Doubler TW, Kilo V, Suttner RJ, Klein III JH, Schmidt ME, Gibson PT, Lightfoot DA (1997) Association of field resistance to soybean sudden death syndrome (SDS) and cyst nematode (SCN). Crop Sci 37:965–971
Chase K, Jarvik T, Lark KG, Mansur LM, Orf JH, Terry LI (2001) Soybean quantitative trait loci for resistance to insects. Crop Sci 40:375–382
Chiari L, Piovesan ND, Naoe LK, Jose IC, Viana JMS, Moreira MA, De Barros EG (2004) Genetic parameters relating isoflavone and protein content in soybean seeds. Euphytica 138:55–60
de Bruxelles GL, Roberts MR (2001) Signals regulating multiple responses to wounding and herbivores. Crit Rev Plant Sci 20:487–521
DeLucia EH, Casteel CL, Nabity PD, O’Neill BF (2008) Insects take a bigger bite out of plants in a warmer, higher carbon dioxide world. Proc Natl Acad Sci USA 105:1781–1782
Gebhardt JS, Wadsworth GJ, Matthews BF (1998) Characterization of a soybean aspartate aminotransferase cDNA encoding isozymes that are differentially targeted to two subcellular compartments. Plant Mol Biol 37:99–108
Gould GE (1963) Japanese beetle damage to soybeans and corn. J Econ Entomol 56:776–781
Hammond RB, Cooper RL (1989) Development and antibiosis of released soybean germplasm lines resistant to Mexican bean beetle (Coleoptera: Coccinellidae). J Econ Entomol 82:259–263
Hammond RB, Bierman P, Levine E, Cooper RL (2001) Field resistance of two soybean germplasm lines, HC95–24 MB, against bean leaf beetle (Coleoptera: Chrysomelidae), western corn rootworm (Coleoptera: Chrysomelidae), and Japanese beetles (Coleoptera: Scarabaidae). J Econ Entomol 94:1594–1601
Iqbal MJ, Yaegashi S, Ahsan R, Shopinski KL, Lightfoot DA (2005) Root response to Fusarium solani f. sp. glycines: temporal accumulation of transcripts in partially resistant and susceptible soybean. Theor Appl Genet 110:1429–1438
Kassem A, Meksem K, Njiti V, Iqbal MJ, Banz WJ, Winters TA, Wood AJ, Lightfoot DA (2004) Definition of soybean genomic regions that control seed phytoestrogen amounts. J Biotech Biomed 2:52–60
Kassem MA, Shultz J, Meksem K, Cho Y, Wood AJ, Iqbal MJ, Lightfoot DA (2006) An updated ‘Essex’ by ‘Forrest’ linkage map and first composite interval map of QTL underlying six soybean traits. Theor Appl Genet 113:1015–1026
Kazi S, Shultz J, Bashir R, Afzal J, Njiti V, Lightfoot DA (2008) Separate loci underlie resistance to soybean sudden death syndrome in ‘Hartwig’ by ‘Flyer’. Theor Appl Genet 116:967–977
Kazi S, Shultz J, Afzal J, Hashmi R, Jasim M, Bond J, Arelli P, Lightfoot DA (2009) Iso-lines and inbred-lines confirmed loci that underlie resistance from cultivar ‘Hartwig’ to three soybean cyst nematode populations Theor Appl Genet 117 (in press)
Killen TC, Lambert L (1986) Evidence for different genes controlling insect resistance in three soybean genotypes. Crop Sci 26:869–871
Komatsu K, Okuda S, Takahashi M, Matsunaga R, Nakazawa Y (2005) QTL mapping of antibiosis resistance to common cutworm (Spodoptera litura Fabricius) in soybean. Crop Sci 45:2044–2048
Ladd TL Jr (1987) Japanese beetle (Coleoptera:Scarabaeidae): influence of favored food plants on feeding response. J Econ Entomol 80:1014–1017
Ladd TL Jr (1989) Japanese beetle (Coleoptera: Scarabaeidae): feeding by adults on minor host and non-host plants. J Econ Entomol 82:1616–1619
Lander ES, Green P, Abrahamson J, Barlow A, Daly MJ, Lincoln SE, Newburg L (1987) MAPMAKER: an interactive computer package for constructing primary genetic linkage map of experimental and natural populations. Genomics 1:174–181
Lightfoot DA (2008) Soybean genomics: developments through the use of cultivar Forrest. Int J Plant Genome 2008:1–22. doi:10.1155/2008/793158
Lightfoot DA, Meksem K (2001) Novel polynucleotides and polypeptides relating to loci underlying Resistance to Soybean Cyst Nematode and methods of use thereof. Patent pending. # 2002 0144310). Filing Date 01-29-2001
Lightfoot DA, Njiti VN, Gibson PT, Kassem MA, Iqbal JM, Meksem K (2005) Registration of Essex × Forrest recombinant inbred line (RIL) mapping population. Crop Sci 45:1678–1681
Loughrin JH, Potter DA, Hamilton-Kemp TR, Byers ME (1996) Volatile compounds from crabapple cultivars (Malus spp.) differing in susceptibility to the Japanese beetle (Popillia japonica Newman). J Chem Ecol 22:1295–1305
Mahalingham R, Skorupska HT (1996) Cytological expression of early response to infection by Heterodera glycines Ichinohe in resistant PI 437654 soybean. Genome 39:986–998
Martinez de Ilarduya O, Xie Q, Kaloshian I (2003) Aphid-induced defense responses in Mi-1-mediated compatible and incompatible tomato interactions. Mol Plant Microb Interact 16:699–708
Meksem K, Pantazopoulos P, Njiti VN, Hyten DL, Arelli PR, Lightfoot DA (2001) ‘Forrest’ resistance to the soybean cyst nematode is bigenic: saturation mapping of the rhg1 and Rhg4 loci. Theor Appl Genet 103:710–717
Mewis I, Appel HM, Hom A, Raina R, Schultz JC (2005) Major signaling pathways modulate Arabidopsis glucosinolate accumulation and response to both phloem feeding and chewing insects. Plant Physiol 138:1149–1162
Nakazawa Y (2005) QTL mapping of antibiosis resistance to common cutworm (Spodoptera litura Fabricius) in soybean. Crop Sci 45:2044–2048
Narvel JM, Walker DR, Rector BG, All JN, Parrott WA, Boerma HR (2001) A retrospective DNA marker assessment of the development of insect resistant soybean. Crop Sci 41:1931–1939
Nombela G, Williamson VM, Muniz M (2003) The root-knot nematode resistance gene Mi-1.2 of tomato is responsible for resistance against the whitefly Bemisia tabaci. Mol Plant Microbe Interact 16:645–649
Patton CA, Ranney TG, Burton JD, Walgenbach JF (1997a) Natural pest resistance of Prunus taxa to feeding by adult Japanese beetles: role of endogenous allelochemicals in host plant resistance. J Am Soc Hortic Sci 122:668–672
Patton CA, Ranney TG, Burton JD, Walgenbach JF (1997b) Feeding responses of Japanese Beetle to naturally occurring metabolites found in rosaceous plants. J Environ Hort 15:222–227
Potter DA, Held DW (2002) Biology and management of the Japanese Beetle. Ann Rev Entomol 47:175–205
Rector BG, All JN, Parrott WA, Boerma HR (2000) Quantitative trait loci for antibiosis resistance to corn earworm in soybean. Crop Sci 40:233–238
SAS Inc., Cary, NC (2006)
Schuster I, Abdelnoor RV, Marin SRR, Carvalho VP, Kiihl AS, Silva JFV, Sedyama CS, Barros EG, Moreira MA (2001) Identification of a new major QTL associated with resistance to the soybean cyst nematode (Heterodera glycines). Theor Appl Genet 102:91–96
Shultz JL, Kazi S, Afzal JA, Bashir R, Lightfoot DA (2007) The development of BAC‐end sequence‐based microsatellite markers and placement in the physical and genetic maps of soybean. Theor Appl Genet 114:1081–1090
Smith AW, Hammond RB, Stinner BR (1988) Influence of rye-cover crop management on soybean foliage arthropods. Environ Entomol 17:109–114
Snedecor GW, Cochran WG (1980) “Statistical Methods”©. The Iowa State University Press, Ames. ISBN 0-8138-1560-6
Szalma SJ, Buckler ES IV, Snook ME, McMullen MD (2005) Association analysis of candidate genes for maysin and chlorogenic acid accumulation in maize silks. Theor Appl Genet 110(7):1324–1333
Tigreros N, Switzer PV (2009) Observations of multiple mating under field conditions for Japanese Beetles (Popillia japonica Newman) (Coleoptera: Scarabaeidae). J Kansas Entomol Soc 82:151–159
Triwitayakorn K, Njiti VN, Iqbal MJ, Yaegashi S, Town C, Lightfoot DA (2005) Genomic analysis of a region encompassing QRfs1 and QRfs2: genes that underlie soybean resistance to sudden death syndrome. Genome/Génome 48:125–138
Underwood N, Rausher M (2002) Comparing the consequences of induced and constitutive plant resistance for herbivore population dynamics. Am Nature 160:20–30
Walker DR, All JN, McPherson RM, Boerma HR, Parrott WA (2000) Field evaluation of soybean engineered with a synthetic cry1Ac transgene for resistance to corn earworm, soybean looper, velvetbean caterpillar (Lepidoptera: Noctuidae), and lesser cornstalk borer (Lepidoptera: Pyralidae). J Econ Entomol 93:613–622
Walker DR, Narvel JM, Boerma HR, All JN, Parrott WA (2004) A QTL that enhances and broadens Bt insect resistance in soybean. Theor Appl Genet 109:1051–1057
Webb DM, Baltazar BM, Arelli PR, Schupp J, Keim P, Clayton K, Ferreira AR, Owens T, Beavis WD (1995) QTL affecting soybean cyst-nematode resistance. Theor Appl Genet 91:574–581
Yesudas CR (2007) Seed and leaf trait correlations and identification of underlying loci in soybean. MS thesis, PLB, SIUC, Carbondale IL, 161pp
Zavala JA, Casteel CL, Nabity PD, Berenbaum MR, DeLucia EH (2009) Role of cysteine proteinase inhibitors in preference of Japanese beetles (Popillia japonica) for soybean (Glycine max) leaves of different ages and grown under elevated CO2. Oecologia 161:35–41
Acknowledgments
This research was funded by grants from the College of Agriculture and Office of the Vice Chancellor for Research. The authors thank Drs. P. Gibson, O. Myers Jr. and M. Schmidt for assistance with germplasm development and maintenance from 1991 to 2000. We thank the “Soybean Genome Project”, at DoE Joint Genome Institute, for the timely release of the WGS reads, scaffolds and genome sequence.
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by C. Gebhardt.
Rights and permissions
About this article
Cite this article
Yesudas, C.R., Sharma, H. & Lightfoot, D.A. Identification of QTL in soybean underlying resistance to herbivory by Japanese beetles (Popillia japonica, Newman). Theor Appl Genet 121, 353–362 (2010). https://doi.org/10.1007/s00122-010-1314-9
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00122-010-1314-9