
Abstract
Drought resistance of rice is a complex trait and is mainly determined by mechanisms of drought avoidance and drought tolerance. The present study was conducted to characterize the genetic basis of drought resistance at reproductive stage in field by analyzing the QTLs for drought response index (DRI, normalized by potential yield and flowering time), relative yield, relative spikelet fertility, and four traits of plant water status and their relationships with root traits using a recombinant inbred population derived from a cross between an indica rice and upland rice. A total of 39 QTLs for these traits were detected with individual QTL explained 5.1–32.1% of phenotypic variation. Only two QTLs for plant water status were commonly detected in two environments, suggesting different mechanisms might exist in two types of soil conditions. DRI has no correlation with potential yield and flowering time under control, suggesting that it can be used as a good drought resistance index in field conditions. The co-location of QTLs for canopy temperature and delaying in flowering time suggested a usefulness of these two traits as indexes in drought resistance screening. Correlation and QTL congruence between root traits and putative drought tolerance traits revealed that drought avoidance (via thick and deep root traits) was the main genetic basis of drought resistance in sandy soil condition, while drought tolerance may play more role in the genetic basis of drought resistance in paddy soil condition. Therefore, both drought mechanisms and soil textures must be considered in the improvement of drought resistance at reproductive stage in rice.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Drought frequently caused loss of yield in rice, one of the staple food crops in Asian countries. Grain yield can be drastically reduced if drought stress occurs during flowering, so developing drought resistant cultivars especially with good performance under late season drought stress is one of the major objectives in rice breeding programs (Boonjung and Fukai 1996; Pantuwan et al. 2002b). However, the progress in breeding for drought resistance is rather slow in rice due to the complexity of the trait and poor understanding of the genetic basis and mechanism of drought resistance in real field conditions, especially at reproductive stage.
Drought tolerance (DT) and drought avoidance (DA) are two major mechanisms for drought resistance of rice at late season. DA can be achieved via enhanced water uptake and reduced water loss while DT is achieved mainly via osmotic adjustment (OA) and antioxidant capacity (Nguyen et al. 1997; Zhang et al. 2001). Putative DA and DT related traits including root density at depth and osmotic adjustment have been well studied and reviewed in rice (Fukai and Cooper 1995; Nguyen et al. 1997). Leaf water potential, leaf rolling, leaf drying, canopy temperature and delay in flowering time can reflect the internal plant water status under water stress, and these traits can be considered as integrative traits to identify drought resistant genotypes (Garrity and O’Toole 1995; Pantuwan et al. 2002a; Jongdee et al. 2002). Although QTL mapping for these traits has been carried out (Champoux et al. 1995; Ali et al. 2000; Zheng et al. 2000; Zhang et al. 2001; Kamoshita et al. 2002; Price et al. 2002; Robin et al. 2003), limited work has been conducted on evaluating their contribution to yield stability in rice. Although some root traits were found associated with plant water status or yield under stress in previous reports (Champoux et al. 1995; Venuprasad et al. 2002; Babu et al. 2003; Lanceras et al. 2004), QTLs for root traits were also found not related to these for leaf rolling and leaf drying score under drought stress (Price et al. 2002). These studies suggest that the genetic basis of drought resistance in rice is extremely complex.
Phenotypic variation resulted from segregation of flowering time in a population often made the drought phenotyping inaccurate. Although staggering sowing date proved to be useful (Lanceras et al. 2004), heading date of a large population still can not be easily synchronized because of genotype×environment interaction (Price and Courtois 1999). To remove the effect from the variation of phenology and yield potential, Bidinger et al. (1987) developed a method to assess drought resistance in pearl millet by using drought response index (DRI), and DRI was also used in the evaluation of drought resistance in rice (Garrity and O’Toole 1994; Pantuwan et al. 2002a).
In this study, drought resistance testing was conducted for a RIL population in field with different types of soil on the basis of synchronization flowering time. QTL mapping was performed for drought resistance indexes (including DRI, relative yield and relative spikelet fertility) and plant water status traits in different types of soil conditions and phenotypic correlations between these drought resistance and root traits were analyzed. The goal of this work is to gain insight into the genetic basis of drought resistance of rice at reproductive stage in different field conditions.
Materials and methods
The experimental population, planting and drought stress
A population consisting of 180 recombinant inbred lines (RILs at F8/F10generation) was developed from a cross between a paddy rice Zhenshan 97 (Oryza sativa L. ssp. indica), and an upland rice IRAT109 (O. sativa L. ssp. japonica). Zhenshan 97 is a maintainer line for a number of elite hybrids widely cultivated in China, and IRAT109 is introduced from Cote d’Ivoire.
The experiments were conducted in two field conditions: natural paddy soil (with pH of 5.8, structure of microaggregate and texture of silty loam) and artificially refined sandy soil (with pH of 6.2, structure of microaggregate and texture of loamy sand). Sowing time was staggered for the lines with interval of 8 days or so to allow a synchronized flowering for all materials based on the data of heading date of this population collected in previous year. All lines were tested under control (with normal irrigation) and drought stress (at reproductive stage) conditions respectively.
Field experiments were carried out following the randomized complete block design with three replications for both control and drought stress conditions. Twenty seedlings (approximately 25 days old) for each line were transplanted into a two-row plot, with the distance of 16.5 cm between the plants within a plot, and 33 cm between adjacent plots. To drain the water rapidly, deep ditch (2 m in depth) was constructed to surround the field under a moveable rain-off shelter. The distance from ditch was same for the lines within a replication thus reducing the variation from the speed of water drainage at different location in the field.
For stress treatment, two consecutive drying cycles were imposed in order to prevent the plants from dying completely and make most of the lines experience drought stress at reproductive stage when plants started panicle initiation. Stress was realized by stopping irrigation, draining out of water (water table kept at 1 m below soil surface) and keeping off rainfall using the shelter. Drought stress was removed by irrigating the field to full water capacity when monitored soil moisture (0–20 cm layer) below half of soil water capacity (16.4% by volume in paddy soil field and 13.9% by volume in sandy soil field). The second round of drought stress was applied to same degree. After drought stress, normal irrigation was followed throughout the late stages of rice.
The soil water content was monitored using a TRIME-T3 sensor device (TRIME-T3, Ettlingen, Germany) by the method of time domain reflectometry (TDR).
Traits and measurements
Drought resistant indexes, including drought respond index (DRI), relative yield (RY), relative spikelet fertility (RSF), and plant water status related traits, including canopy temperature (CT), leaf rolling score (LRS), leaf drying score (LDS), heading date and delay in flowering time by drought (DFT) were recorded.
Leaf rolling score was visually scored with scale from “0” to “4” at noon before re-irrigation when the difference among the lines became most obvious following the method by Turner (1997). A score of “0” indicates no symptom of rolling, and the score “4” indicates complete leaf rolling. Canopy temperature was measured at noon just before re-irrigation using a hand-hold infrared thermometer (Raytek Corporation, CA, USA) as described by Garrity and O’Toole (1995) and Pantuwan et al. (2002b). LDS was visually scored with a scale from “0” to “4” just before re-irrigation. A score of “0” indicates no symptom of drying, and the score “4” indicates a susceptible type (about half of the leaf areas dried). DFT was calculated as the differences of heading date under drought stress and control conditions. Heading date was determined visually when about 50% of the tillers in each plot were headed.
Ten consecutive plants in the middle of each row were measured for yield and spikelet fertility under both control and stress conditions. Spikelet fertility (%) was defined as the rate of the number of grains to the total number of spikelets of a plant. The relative performance of RY and RSF was simply measured as the ratios of the measurements taken under drought stress and control conditions. DRI was calculated as described by Pantuwan et al. (2002b) using the format as follows:
where Yacti is the actual grain yield (grain yield under drought stress) for each line, Yest is the estimated grain yield for each line, and SE is the standard error of the Yest of all lines. Estimated grain yield (Yest) was derived from the calculation using multiple linear regression analysis since yield and flowering time under control was strongly correlated with yield under drought stress. Yp i and FT i is potential yield and flowering time respectively under control condition for ith genotype, and a,b, c are the regression coefficients.
Root traits were investigated for maturated plants of each line growing in PVC pipes (1 m in depth and 20 cm in diameter), one plant per pipe, under both well water and late season drought stress conditions in the year 2003. The pipes were laid out in six blocks following a randomized complete block design. Drought stress was applied to three of the blocks while the other three blocks were used as control. To measure root traits, the plastic bag containing the soil and roots was pulled out from the PVC tube and laid out on a stand furnished with screen surface. The lowest visible root in the soil after removing the plastic bag was scored as the maximum root depth (MRD, cm). The body of soil and roots was cut into two parts at 30 cm from the basal node of the plant and the soil was washed away carefully to collect roots. The volumes (ml) of roots from the two parts were measured in a cylinder using the water replacing method. The root mass below 30 cm was considered to be deep root, from which a number of measurements were derived, including total root volume (RV) and deep root rate for volume (DRV, %). Root growth rate in depth (RGD, cm/day) and root growth rate in volume (RGV, ml/day) were obtained as dividing maximum root depth and total root volume, respectively, by root growth period (number of days from sowing to heading of the plant). Drought induced root growth was evaluated by two traits: drought induced root growth in depth (DIRD, cm) and drought induced deep root rate in volume (DIDRV, ml) that were calculated as the differences of maximum root depth and deep root rate in volume between the measurements obtained from drought stress and control conditions. Root thickness (RTH, cm) was measured at 2 cm below the stem bases. Biomass (BM, g) of each plant was weighted after the plants dried by air.
DNA markers, map construction and QTL analysis
A total of 245 nuclear simple sequence repeat (SSR) markers were used for constructing the linkage map. The SSR primers and marker assays essentially followed Temnykh et al. (2000, 2001) and McCouch et al (2002). The program of Mapmaker/EXP 3.0 (Lincoln et al. 1992) was used to construct the genetic linkage map (LOD score was 3.0, map function was Kosambi). The marker order was well matched with the map of McCouch et al (2002). The means of the traits were used to identify QTLs by the method of complexity interval mapping (CIM) using the software of QTLCartographer 2.0 (Zeng 1994). The permutation method was used to obtain the thresholds (1.97–2.42 for these traits in two soil conditions) of the experiment based on 1,000 runs of randomly shuffling the trait values (P=0.05), thus a threshold of LOD score 2.4 was used in this study.
Results
Flowering time synchronizing by staggering sowing dates
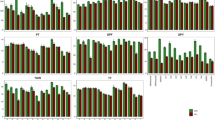
The span of flowering time among this RIL population was more than 55 day, and it was shortened to 40 day or so by staggering sowing dates both in paddy and sandy soil conditions. Since the lines flowering before drought stress were unaffected by drought stress and the line flowering extremely late were affected by low temperature, these lines (appropriately 15% under both soil conditions) were removed from analyses. Yield under drought stress was positively and significantly correlated with potential yield (0.59 in paddy soil field and 0.42 in sandy soil field), and strong correlation was also detected between yield under drought stress and flowering time under control conditions (Fig. 1), thus DRI was calculated and used as drought resistance index in this study.

Scatter plots of DRI, relative yield and relative spikelet fertility against yield and flowering time under control conditions.
Phenotypic variation of the parents and RILs
The phenotypic differences between parents as well as the variation in the RIL population are summarized in Table 1. Transgressive segregations were observed in the RIL population for all the traits investigated. Both skewness and kurtosis for all traits except for DFT (only for kurtosis) were less than 1.0 (Table 1), suggesting that the segregation of all the traits in the RIL population fits a normal distribution. ANOVA of the data collected in two field conditions indicated that variation due to genotype differences was highly significant for all the traits, although the relative proportions of variance explained varied from one trait to another (Table 2). Variation between replications was also significant under drought stress in paddy field. However, it was not significant for relative spikelet fertility, leaf rolling and leaf drying score in sandy field. These indicated that the development of drought stress was more uniform in the refined sandy soil.
IRAT109 showed more drought resistance than Zhenshan 97 in both soil conditions by having higher values of DRI, relative yield and relative spikelet fertility. IRAT109 also showed more drought resistance than Zhenshan 97 in both soil conditions as indicated by having low canopy temperature and fewer days for delaying in flowering time under drought stress. Reverse performance was observed between the parents for the other two plant water status traits (Table 1). The leaf drying score and leaf rolling score of IRAT109 were higher than that of Zhenshan 97 in both soil conditions.
When the data collected from the two kinds of soil conditions were compared, DRI, relative yield and relative spikelet fertility in sandy soil condition were higher than that in paddy soil condition for both parents and the means of RILs (Table 1), indicating that the stress developed more severe in the paddy soil condition. This may be explained by the fact that the roots can easily penetrate deeper in sandy soil condition. Same trend of difference between the two soil conditions was also observed for a few plant water status related traits including delaying in flowering time, leaf rolling score and leaf drying score. However, canopy temperature was higher in sandy soil condition.
Correlations analysis of the traits
Figure 1 illustrates the relationships of DRI, relative yield and relative fertility with yield, flowering time and spikelet fertility under control conditions. The data suggested that DRI was not correlated with both potential yield and flowering time under control conditions. Similarly, there was little correlation between relative yield and relative spikelet fertility with yield or spikelet fertility under control conditions. These results suggested that the effect of yield potential and spikelet fertility under control conditions on drought resistance evaluation was eliminated by using relative parameters. However, relative yield and relative spikelet fertility was marginally correlated with flowering time under control, which was due to the residual variation of flowering time that still existed in this population even though seed sowing was staggered to synchronize the flowering time.
The three drought resistance indexes were highly intercorrelated in two soil conditions (Table 3), suggesting that drought resistance at reproductive stage in field was highly related to relative yield or relative spikelet fertility on the basis of synchronization of flowering time.
Three plant water status related traits, including canopy temperature, leaf drying score and leaf rolling score were also highly intercorrelated in two field conditions, while delaying in flowering time was significantly correlated with canopy temperature only in the sandy field (Table 3). Negative and significant correlations were detected between DRI with delaying in flowering time, canopy temperature and leaf rolling score at least in one field condition. Interestingly, the correlation between relative spikelet fertility and delaying in flowering time was significant but in the opposite way in two field conditions.
Correlations between drought resistance indexes (plant water status traits as well) and normalized root traits (the value of root traits divided by biomass under control condition) are given in Table 4. In general DRI, relative yield and relative spikelet fertility had no correlation (only a few marginally negatively correlation) with root traits under paddy soil condition. In sandy soil condition, however, significant and positive correlations were detected between drought resistance indexes and root thickness, deep root traits (including maximum root depth, root growth rate in depth and drought induced root growth in depth). Drought resistance index was negatively correlated with root size (including the traits of root volume, root growth rate in volume) both under control and stress conditions. These results suggested soil conditions have great affection on drought resistance in field.
In general, thick and deep root traits were negatively correlated with delaying in flowering time in two field conditions. However, plant or root size related traits were negatively and significantly correlated with canopy temperature, leaf drying score and leaf rolling score in paddy soil condition (Table 4).
The linkage map
A linkage map was constructed based on the data of the 245 SSR markers assayed on the 180 RIL lines by Mapmaker analysis (Fig. 2). The map covered a total length of 1,530 cM with an average interval of 6.2 cM between adjacent markers.

Molecular genetic linkage map of rice based on the RIL population from a cross between Zhenshan 97 with IRAT109. Distances are given in Kosambi centiMorgans. The QTLs detected in this study were on the left of the chromosomes, and the associating QTLs for root traits (Yue 2005) and OA detected by Robin et al. (2003) were marked on the right. The QTLs in bold were detected in two soil conditions. The italic QTLs indicate that the alleles for increasing trait values were from Zhenshan 97
QTL detection
Quantitative trait loci detected for drought resistant indexes are listed in Table 5 (also Fig. 2). A total of 16 QTLs were resolved for DRI, relative yield, relative spikelet fertility, but none of them were detected in both field conditions, and individual QTL explained 4.9–32.1% of phenotypic variation. In sandy soil condition, more QTLs were identified and explained more phenotypic variation, especially for the index of DRI.
A total of 23 QTLs were resolved for the four plant water status related traits including two detected in both field conditions, 21 of them only detected in one field condition, and individual QTL explained 5.1–23.5% of phenotypic variation (Table 5). In sandy soil condition, more QTLs were identified for these traits and explained more phenotypic variation, especially for the index of canopy temperature.
Congruence of QTL
There were two regions congregated more than three QTLs and seven regions contained two QTLs. For examples, the region RM237-RM403 on chromosome 1 has QTLs for DRI, relative yield and delaying in flowering time simultaneously, and the region RM125-MRG4499 on chromosome 7 is clustered with QTLs for relative yield, relative spikelet fertility, delaying in flowering time and leaf drying score (Fig. 2). Among these QTL-clustered regions, seven regions were associated with both drought resistant indexes and plant water status related traits, such as the region RM279-RM555 on chromosome 2, RM176-RM494 on chromosome 6 and the region RM339-RM342 on chromosome 8 controlled DRI and canopy temperature simultaneously. Moreover, there were two regions controlling DRI and delaying in flowering time simultaneously. However, QTL for leaf rolling score had no overlap with QTL for drought resistant indexes, and only one region was clustered QTLs for drought resistant indexes and leaf drying score.
Discussion
In this study, QTL mapping for three drought resistant indexes and four plant water status related traits in field conditions was conducted under two types of soil conditions using a RIL population. For the first time, we mapped drought response index (DRI) and studied the role of drought tolerance and drought avoidance in the drought resistance at reproductive stage in rice. Our results suggested that using DRI can eliminate the effect of segregation of yield potential and flowering time that can affect the phenotyping of drought resistance under field conditions, and the genetic basis of drought resistance was different in different types of soil conditions.
Drought resistance indexes at reproductive stage in field
Drought resistance at reproductive stage in rice can be assessed by drought susceptibility index (corresponding to relative yield) by reducing the interference from the variation of potential yield (Fischer and Maurer 1978), and the variation in potential yield and flowering time can be reduced by using DRI (Bidinger et al. 1987; Garrity and O’Toole 1994; Pantuwan et al. 2002a). Spikelet fertility can be greatly affected when stress occurs near flowering (Laffite et al. 2003), so relative spikelet fertility can also be used to assess drought resistance. In this study, DRI had no correlation with potential yield and flowering time, while other yield related indexes (relative yield and relative spikelet fertility) had correlation with flowering time. These indicated that the effects from the variation of potential yield and flowering time were eliminated using DRI as parameter, and DRI can be also used as indirect index for drought resistance at reproductive stage. Although strong positive correlations were detected between DRI, relative yield and relative spikelet fertility, congruence of QTL for DRI and relative yield was found only in one chromosome region in the experiment conducted in paddy field. This suggested that the genetic basis of DRI and relative parameters might be different. On the other hand, canopy temperature and delaying in flowering time was negatively and significantly correlated to DRI, and common QTLs were detected for DRI and canopy temperature and delaying in flowering time in five regions. This indicated that canopy temperature and delaying in flowering time can also be used as good indirect indexes for evaluation of drought resistance in rice, and this view was also suggested by Garrity and O’Toole (1995) and Pantuwan et al. (2002a). However, the correlation between leaf drying score and DRI was not strong, and there was no QTL for leaf rolling score and leaf drying score overlapped with the QTLs for DRI, suggesting that the visual leaf phenotypes may not be used as indexes for evaluation of drought resistance at reproductive stage under field conditions.
The mechanisms of drought resistance at reproductive stage in field
Drought tolerance (DT) and drought avoidance (DA) are two major mechanisms for drought resistance at late season in rice. DA can be achieved via enhanced water uptake and reduced water loss and DT via osmotic adjustment (OA) and antioxidant capacity (Nguyen et al. 1997; Zhang et al. 2001). Comparison with the QTLs for root traits detected in the same population, 37.5% of QTLs (five regions) for drought resistant indexes and 47.8% of QTLs (nine regions) for plant water status related traits in this study were collocated with the QTLs for root traits (Yue 2005, Fig. 2). These indicated that drought avoidance might play an important role in drought resistance at reproductive stage in rice. However, no common QTL for drought resistant indexes was detected in two soil conditions. This implied that different mechanisms might be involved in the establishment of drought resistance under different types of soil conditions.
Correlation analysis revealed that DRI was positively and significantly correlated with thick and deep root traits, and negatively correlated to plant size (biomass under control) and root size in sandy field. However, there was no root trait correlated with DRI in paddy field. The genetic basis of these relationships can be explained by the co-location of the QTLs for these traits. The two QTLs for DRI detected in paddy soil condition had no overlapping with QTLs for root traits, while three of the five DRI QTLs identified in sandy field were associated with QTLs for root traits (Fig. 2). These results suggested that DA may be the main drought resistance mechanism via thick, deep root traits and little plant or root size in sandy field.
Near the genomic region RM279-RM555 on chromosome 2 in which QTL for osmotic adjustment (OA) was ever reported (Robin et al., 2003), QTLs for DRI and canopy temperature were detected in paddy field in this study. These indicated that the physiological traits (such as OA) may play more important role in paddy field in which root traits can hardly play any role in drought resistance because of the hard layer of plough pan can largely prevent roots from downward penetrating in paddy field. This data also supported the suggestion that physiological traits will play more roles in conferring drought resistance when drought stress become severe (Blum 1993; Fukai and Cooper 1995; Pantuwan et al. 2002b; Lafitte et al. 2003).
The relationships between soil types and drought resistance
Root diameter, rooting depth, and root/shoot dry weight ratio were thought to be related to drought avoidance in rice based on the positive correlations between these characters and visual scores of plant vigor in upland field drought screening trials (O’Toole and Soemartono 1981; Yoshida and Hasegawa 1982). Root growth in depth depends on the ability of root penetration into the hard pans in lowland field conditions. Although genotypic variation of root penetration ability existed and QTLs had also been detected for this trait (Yu et al. 1995; Ray et al. 1996), there was no evidence proving that this trait is likely to be useful in lowland environment. Since small plants normally consume less water than large plants under drought stress, the data of root traits should be normalized by plant size for assessment of the role of root traits in drought resistance. In this study, the correlations between DRI and normalized thick and deep root traits were significant in sandy field but not in paddy field, clearly suggesting that the contribution of root traits to drought avoidance is associated with soil types. In this study, the soil structure and pH was similar in two field conditions, while their soil texture was very different. So soil texture is mostly crucial to root penetration in rice field. For the roles of root traits in drought avoidance, the results were also inconsistent in previous studies (Champoux et al. 1995; Price et al. 2002; Babu et al. 2003), which may be caused by different root traits investigated or different types of soil conditions in different reports.
Drought stress developed before flowering often has an effect in delaying flowering time and the delaying in flowering time was negatively associated with grain yield, fertile panicle percentage and filled grain percentage in rainfed lowlands (Pantuwan et al. 2002a). In this study, delaying in flowering time was positively correlated with relative spikelet fertility in paddy field, but the correlation was negative in the sandy field, suggesting the effect of delayed flowering time by drought stress on yield stability depends on soil types as well.
Large genotype-by-environment interaction observed for grain yield of rainfed lowland rice in northeast Thailand encountered different types of drought across years (Cooper et al. 1999). Lack of consistency of DRI across experiments has also been demonstrated by Pantuwan et al. (2002a). These suggested that different mechanisms may exist in drought resistance under different types of environments. In our study, we conclude that the mechanism for drought resistance at reproductive stage in rice is indeed associated with soil types. Taken together, the drought mechanisms, drought types, soil textures and the important role of drought avoidance should be integratively considered in the improvement of drought resistance at reproductive stage in rice.
References
Ali ML, Pathan MS, Zhang J, Bai G, Sarkarung S, Nguyen HT (2000) Mapping QTLs for root traits in a recombinant inbred population from two indica ecotypes in rice. Theor Appl Genet 101:756–766
Babu RC, Nguyen BD, Chamarerk V, Shanmugasundaram P, Chezhian P, Jeyaprakash P, Ganesh SK, Palchamy A, Sadasivam S, Sarkarung S, Wade LJ, Nguyen HT (2003) Genetic analysis of drought resistance in rice by molecular markers: association between secondary traits and field performance. Crop Sci 43:1457–1469
Bidinger FR, Mahalakshmi V, Rao GDP (1987) Assessment of drought resistance in pearl millet [ Pennisetum americanum (L.) Leeke]. II Estimation of genotypes response to stress. Aus J Agric Res 38: 49–59
Blum A (1993) Selection for sustained production in water deficit environments. In: Buxton DR, Shibles R, Forsberg RA, Blad BL, Asay KH, Paulsen GM, Wilson RF (eds) International crop science, I. Crop Science Society of America, USA, pp 343–347
Boojung H, Fukai S (1996) Effects of soil water deficit at different growth stage on rice growth and yield under upland conditions. 2. Phenology, biomass production and yield. Field Crop Res 48:47–55
Champoux MC, Wang G, Sarkaruag S, Mackill DJ, O’Toole JC, Huang N, McCouch SR (1995) Locating genes associated with root morphology and drought avoidance in rice via linkage to molecular markers. Theor Appl Genet 90:969–981
Cooper M, Rajatasereekul S, Immark S, Fukai S, Basnayake J (1999) Rainfed lowland rice breeding strategies for Northeast Thailand. I. Genotypic variation and genotype×environment interactions for grain yield. Field Crops Res 64:131–151
Fischer RA, Maurer R (1978) Drought resistance in spring wheat cultivars. I. Grain yield responses. Aust J Agric Res 29: 897–916
Fukai S, Cooper M (1995) Development of drought-resistant cultivars using physio-morphological traits in rice. Field Crops Res 40:67–86
Fukai S, Pantuwan G, Jongdee B, Cooper M (1999) Screening for drought resistance in rainfed lowland rice. Field Crops Res 64:61–74
Garrity DP, O’Toole JC (1994) Screening rice for drought resistance at the reproductive phase. Field Crop Res 39:99–110
Garrity DP, O’Toole JC (1995) Selection for reproductive stage drought avoidance in rice, using infrared thermometry. Agron J 87:773–779
Jongdee B, Fukai S, Cooper M (2002) Leaf water potential and osmotic adjustment as physiological traits to improve drought tolerance in rice. Field Crop Res 76:153–163
Kamoshita A, Wade LJ, Ali ML, Pathan MS, Zhang J, Sarkarung S, Nguyen HT (2002) Mapping QTLs for root morphology of a rice population adapted to rainfed lowland conditions. Theor Appl Genet 104:880–893
Lafitte R, Blum A, Atlin G (2003) Using secondary traits to help identify drought-tolerant genotypes. In: Fischer KS, Lafitte R, Fukai S, Atlin G, Hardy B (eds) Breeding rice for drought-prone environments. IRRI, Los Banos, Philippines, pp 37–48
Lanceras JC, Pantuwan G, Jongdee B, Toojinda T (2004) Quantitative trait loci associated with drought tolerance at reproductive stage in rice. Plant Physiol 135:384–399
Lincoln S, Daly M, Lander E (1992) Constructing genetics maps with MAPMAKER/EXP 3.0. Whitehead Institute Technical Report, Whitehead Institute, Cambridge, Massachusctts, USA
McCouch SR, Teytelman L, Xu YB, Lobos KB, Clare K, Walton M, Fu B, Maghirang R, Li Z, Xing Y, Zhang Q, Kono I, Yano M, Fjellstrom R, DeClerck G, Schneider D, Cartinhour S, Ware D, Stein L (2002) Development and mapping of 2240 new SSR markers for rice (Oryza sativa L.). DNA Reserch 9:199–207
Nguyen HT, Babu RC, Blum A (1997) Breeding for drought resistance in rice: physiology and molecular genetics considerations. Crop Sci 37:1426–1434
O’Toole JC, Soemartono (1981) Evaluation of a simple technique for characterizing rice root systems in relation to drought resistance. Euphytica 30:283–290
Pantuwan G, Fukai S, Cooper M, Rajatasereekul S, O’Tool JC (2002a) Yield response of rice (Oryza Sativa L.) genotypes to drought under rainfed lowlands 2. Selection of drought resistant genotype. Field Crops Res 73:169–180
Pantuwan G, Fukai S, Cooper M, Rajatasereekul S, O’Tool JC (2002b) Yield response of rice (Oryza Sativa L.) genotypes to different types of drought under rainfed lowlands. 3. Plant factors contributing to drought resistance. Field Crops Res 73:181–200
Price A, Courtois B (1999) Mapping QTLs associated with drought resistance in rice: Progress, problems and prospects. Plant Growth Regulation 29:123–133
Price AH, Cairms JE, Horton P, Jones HG, Griffiths H (2002) Linking drought-resistance mechanisms to drought avoidance in upland rice using a QTL approach: progress and new opportunities to integrate stomatal and messophyll responses. J Exp Bot 53:989–1044
Ray JD, Yu L, McCouch SR, Champoux MC, Wang G, Nguyen HT (1996) Mapping quantitative trait loci associated with root penetration ability in rice (Oryza sativa L.). Theor Appl Genet 92:627–636
Robin S, Pathan MS, Courtois B, Lafitte R, Carandang S, Lanceras S, Amante M, Nguyen HT, Li Z (2003) Mapping osmotic adjustment in an advanced back-cross inbred population of rice. Theor Appl Genet 107:1288–1296
Temnykh S, Park WD, Ayres N, Cartihour S, Hauck N, Lipovich L, Cho YG, Ishii T, McCouch SR (2000) Mapping and genome organization of microsatellite sequences in rice (Oryza sativa L.). Theor Appl Genet 100:697–712
Temnykh S, Declerck G, Luashova A, Lipovich L, Cartinhour S, McCouch S (2001) Computational and experimental analysis of microsatellites in rice (Oryza sativa L.): frequency, length variation, transposon associations, and genetic marker potential. Genome Res 11:1441–1452
Turner NC (1997) Further progress in crop water relation. Adv Agron 58:293–339
Venuprasad R, Shashidhar HE, Hittalmani S, Hemamalini GS (2002) Tagging quantitative trait loci associated with grain yield and root morphological traits in rice (Oryza sativa L.) under contrasting moisture regimes. Euphytica 128:293–300
Yoshida S, Hasegawa S (1982) The rice root system: its development and function. In: Drought resistance in crops with emphasis on rice. IRRI, Los Banos, Philippines, pp 97–114
Yu L, Ray JD, O’Toole JC, Nguyen HT (1995) Using of wax-petrolatum layers to simulate compacted soil for screening rice (Oryza sativa L.) root penetration ability. Crop Sci 35:684–687
Yue B (2005) Study on the genetic basis of late season drought resistance in rice. Ph D thesis, Hunzhong Agricultural University, Wuhan, China
Zeng ZB (1994) Precision mapping of quantitative trait loci. Genetics 136:1457–1468
Zhang J, Zheng HG, Aarti A, Pantuwan G, Nguyen TT, Tripathy JN, Sarial AK, Robin S, Babu RC, Nguyen BD, Sarkarung S, Blum A, Nguyen HT (2001) Locating genomic regions associated with components of drought resistance in rice: comparative mapping within and across species. Theor Appl Genet 103:19–29
Zheng HG, Babu RC, Pathan MS, Ali L, Huang N, Courtois B, Nguyen HT (2000) Quantitative trait loci for root-penetration ability and root thickness in rice: comparison of genetic backgrounds. Genome 43:53–61
Acknowledgements
This research was supported by grants from the National Program on the Development of Basic Research, the National Special Key Project on Functional Genomics and Biochips, the National Natural Science Foundation of China, and the Rockefeller Foundation.
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by C. Möllers
Rights and permissions
About this article
Cite this article
Yue, B., Xiong, L., Xue, W. et al. Genetic analysis for drought resistance of rice at reproductive stage in field with different types of soil. Theor Appl Genet 111, 1127–1136 (2005). https://doi.org/10.1007/s00122-005-0040-1
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00122-005-0040-1