
Abstract
Southern corn rust (SCR), Puccinia polysora Underw, is a destructive disease in maize (Zea mays L.). Inbred line Qi319 is highly resistant to SCR. Results from the inoculation test and genetic analysis of SCR in five F2 populations and five BC1F1 populations derived from resistant parent Qi319 clearly indicate that the resistance to SCR in Qi319 is controlled by a single dominant resistant gene, which was named RppQ. Simple sequence repeat (SSR) analysis was carried out in an F2 population derived from the cross “Qi319×340”. Twenty SSR primer pairs evenly distributed on chromosome10 were screened at first. Out of them, two primer pairs, phi118 and phi 041, showed linkage with SCR resistance. Based on this result, eight new SSR primer pairs surrounding the region of primers phi118 and phi 041 were selected and further tested regarding their linkage relation with RppQ. Results indicated that SSR markers umc1,318 and umc 2,018 were linked to RppQ with a genetic distance of 4.76 and 14.59 cM, respectively. On the other side of RppQ, beyond SSR markers phi 041 and phi118, another SSR marker umc1,293 was linked to RppQ with a genetic distance of 3.78 cM. Because the five linkage SSR markers (phi118, phi 041, umc1,318, umc 2,018 and umc1,293) are all located on chromosome 10, the RppQ gene should also be located on chromosome 10. In order to fine map the RppQ gene, AFLP (amplified fragment length polymorphism) analysis was carried out. A total 54 AFLP primer combinations were analyzed; one AFLP marker, AF1, from the amplification products of primer combination E-AGC/M-CAA, showed linkage with the RppQ gene in a genetic distance of 3.34 cM. Finally the RppQ gene was mapped on the short arm of chromosome 10 between SSR markers phi 041 and AFLP marker AF1 with a genetic distance of 2.45 and 3.34 cM respectively.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Southern corn rust (SCR) distributes broadly in Americas, Africa, Asia and Australia. It is caused by Puccinia polysora Underw, and has enormous destructive potential in maize (Zea mays L.) (Melching 1975). Severe epiphytotics were observed in Africa since 1949 and its resulted yield loss was estimated up to 50% (Rhind et al. 1952). Futrell (1975) reported that southern rust reached epiphytotic levels in US in 1972–1974. Rodriguez-Ardon et al. (1980) reported that southern rust caused yield loss of up to 45% on susceptible maize hybrids. In China, Duan and He (1984) reported that southern corn rust was first found in Ledong, Hainan Province, in 1972. It occurred mainly in the southern part of China. In 1998, SCR broke out and for the first time reached epiphytotic levels in the northern part of China, and caused serious yield loss of up to 42% to 53% (Liu and Wang 1999; Wang 1999). In recent years, SCR has become a main maize disease in China.
The epiphytotics of SCR were attributed, in part, to the susceptibility of inbred lines and hybrids of southern rust. Rodriguez-Ardon et al. (1980) suggested that most of the commercial American hybrids were susceptible to southern rust. In China, Ye (2000) made a field survey on maize inbreds and hybrids, and obtained similar results. Therefore, it is desirable to exploit resources of resistance against southern rust to improve the disease resistance of parents used in maize breeding. Using the resistant line as a parent in crossing, is the preferred method to control SCR infestations.
Ullstrup (1965) identified a single dominant-gene, Rpp9, from a South African cultivar that conferred resistance to race pp. 9. Futrell et al. (1975) identified another single dominant gene resistance to pp 9 in a line designated B1138TRpp, but the relationship of this gene to Rpp9 was not studied. Scott et al. (1984) studied the inheritance of resistance found in several lines by using four corresponding segregation populations. One-gene or two-gene models with varying degrees of dominance were used to explain the resistance found in different populations. Tests of allelism with Rpp9 indicated that each line carried a resistance gene either at or closely linked to the Rpp9 locus. Several studies (Bailey et al. 1987; Zummo 1988; Holland et al. 1998) have indicated that partial resistance to southern rust exists in some maize genotypes.
Ye (2000) tested the resistance of 200 inbred lines to SCR on a field survey in Hainan Island; the results indicated that only Qi319 was completely resistant to SCR. Chen et al. (2003) identified the resistance to SCR of ten inbred lines, which have been most widely employed of corn breeding programs in China; the results showed that only Qi319 was highly resistant to SCR both in the field and in greenhouse conditions. So, Qi319 is an elite germplasm in Chinese maize resistant breeding. The objectives of this study were to map the resistance gene by molecular markers (SSRs, AFLPs) and to develop a linkage map of the resistance gene-encompassing region. It will provide a valuable tool for marker-assisted selection (MAS) in maize breeding.
Materials and methods
Plant materials
SCR resistant maize inbred line Qi319 was crossed with five susceptible lines (Luyuan 92, 478, 340, 9801 and Huangzao 4); then five F2 and five BC1F1 populations were developed from the progeny of these crosses.
Pathogen, inoculation and symptom scoring
Inoculation experiments were performed on the parents, F1, F2 and BC1F1 populations in the Experimental Farm-station of Shandong Agricultural University, respectively, in 2001. The SCR pathogen, P. polysora Underw, was obtained from susceptible corn in Lingshui County, Hainan Province. We purified the pathogen by using the detached leaf method (Hooker and Yarwood 1966) and obtained single uredium isolates; then the uredium was cultured by serial transfers and maintained on susceptible corn seedlings in the greenhouse, where temperatures ranged from 18°C to 34°C. The inoculum used in this experiment was a solution of P. polysora uredospores mixed with distilled water containing two drops of Tween-20/0.5 l of water. The parents, F1, F2 and BC1F1 populations were inoculated twice with the inoculum at the 4–5 leaf stage using the brush technique, and then using the airbrush technique 1 week later. The temperatures ranged from 17°C to 34°C in the field. Symptoms were scored at weekly intervals and the first time was taken at 2 weeks after the second inoculation. Final scoring was conducted 30 days after the initial inoculation when SCR symptoms were fully developed.
Statistics and genetic analysis
In the tests, F1 and the parents were taken as controls. The numbers of resistant plants and susceptible plants were counted and calculated in the F2 and BC1F1 populations, respectively. Then, χ2 tests were performed to determine whether the goodness of fit is a ratio of 3:1 in F2 populations, and a ratio of 1:1 in BC1F1 populations.
DNA preparation
Leaves from susceptible plants of the F2 populations were harvested after 2 weeks of the second SCR inoculation. Leaves from resistant plants were harvested individually after 30 days of the initial inoculation, when SCR symptoms were fully developed. Harvested leaves were freeze-dried and well-ground to a powder in liquid nitrogen. DNA extraction was performed according to the standard CTAB (Cetyl trimethyl ammonium Bromide) method (Hoisington et al. 1994), with one additional purification step using chloroform isoamyl-alcohol to obtain high-quality DNA.
SSR analysis
Ullstrup (1965) performed traditional genetic analysis to the resistant gene Rpp9 and showed that Rpp9 was closely linked to the gene Rp, which resulted in resistance to P. sorghi on chromosome 10. We selected 20 SSR primers on chromosome 10 at first, to screen the polymorphic SSR markers by using BSA (bulked segregant analysis). The polymorphic markers on BSA were amplified on the population to verify linkage to the resistant gene. Then, around the polymorphic SSR markers, eight new SSR markers were chosen to further screen more linkage markers to the resistant gene.
SSR amplification was performed as described by Chin et al. (1996), Lubberstedt et al. (1988) and Akagi et al (1996) with minor modification. Each SSR reaction mixture contained 1×PCR buffer, 1.5–2.0 mM of MgCl2, 0.1 mM of each dNTP, 1 U of Taq-polymerase, 150 ng each of the left and right primers, and 50 ng of template DNA in a total volume of 20 μl. The PCR reaction was accomplished in an MJ PTC-100 thermocycler, at standard amplifications of 94°C for 2 min; followed by 30 cycles of 94°C for 1 min, 58°C for 2 min and 72°C for 2 min, then with a final extension step at 72°C for 5 min before cooling to 4°C. The SSR products were separated by electrophoresis in 3.5% agarose gels in preliminary screening. For further SSR screening, each SSR reaction mixture also contained [α-32P]dCTP 0.8 µCi (purchased from Amersham Technology Inc.). The amplification products were separated on the 6% PAGE (polyacrylamide gel electrophoresis) sequencing gel at 100 W for 2.0 h after pre-electrophoreses for 30 min. The gel was then removed from the apparatus, dried with the BRL Model 583 gel dryer and exposed to X-ray film at −70°C for 2–3 days until the film was adequately exposed.
The sequence of SSR primers were obtained from the maize database (http://www.agron.missouri.edu) and synthesized by Bioasia Biotech Lt. Co., Shanghai.
AFLP analysis
AFLP analysis was performed by using the AFLP Analysis System I Kit (GibcoBRL, Life Technologies), with 33P-labelled oligonucleotides according to the suppler’s instructions and as described by Vos and Hogers (1995). All of the 54 primer combinations recommended for maize in the AFLP Analysis System I were screened. The obtained putative linkage markers were confirmed by co-segregation analysis with a large number of F2 individuals. Amplification products were separated on a 6% PAGE sequencing gel, at 100 W for 2.5 h after pre-electrophoresis for 30 min. The gel was then removed, dried and exposed according to the procedure described above.
Primary mapping
For bulked segregant analysis (BSA), an equal amount of DNAs from each of the ten high resistant F2 plants were pooled to form the resistant bulk (BR); while an equal amount of DNAs from each of the ten highly susceptible F2 plants were pooled to form the susceptible bulk (BS). DNA samples of the two parents and the two bulks were subjected to BSA analysis together. Once a potential linkage marker was identified by BSA screening, co-segregation analysis would be carried out in the F2 population, derived from the cross “Qi319×340”, to confirm and determine the genetic linkage between the SCR resistant gene and the marker.
Constructing a local linkage map
Based on the SSR and AFLP data, combined with the phenotypic data, linkage analysis was performed with MAPMAKER Version 3.0 (Lander et al. 1987). Recombination fractions were converted into genetic map distance (cM) using the Kosambi mapping function (Kosambi 1944).
Results
Phenotype observation and genetic analysis of SCR resistance
Inoculation experiments were performed on the parents, F1, F2 and BC1F1 populations in the experimental Farm Station of Shandong Agricultural University, respectively, in 2001. The symptoms began to develop on the leaf surface of susceptible plants about ten days after initial inoculation. After 1 month, almost all leaves of susceptible plants displayed densely SCR pustules on the leaf surface. F1 plants were all resistant, while the F2 and BC1F1 populations segregated significantly. There were two phenotypic responses to SCR in the segregation populations, phenotypic resistant plants with no SCR pustule and with the SCR pustule of the phenotype susceptible plant. The final scoring for resistance and susceptibility was performed after 30 days of the initial inoculation; scoring results were summarized in Table 1. In the five tested F2 populations, the ratios of resistant individuals to susceptible individuals were all in 3:1. While in the five tested BC1F1 populations, the ratios of resistant individuals to susceptible individuals were all in 1:1 (Table 1). Chi-square tests were performed to determine the goodness of fit of a 3:1 ratio or a 1:1 ratio. Results indicated that in any crosses the ratio of resistant plants to susceptible plants was fitted to the 3:1 ratio in the F2 populations and the 1:1 ratio in the BC1F1 populations (Table 1). These results clearly pointed out that a single dominant gene controlled the resistance to the pathogen P. polysora Underw, and the resistance was from the resistant parent Qi319, which was referred to as RppQ temporarily.
Screening of SSR markers linked to RppQ and primary mapping of the RppQ gene
Out of the F2 population of the cross “Qi319×340”, 120 selected individuals (60 resistant plants and 60 susceptible plants) were used for screening molecular markers putatively linked to the SCR resistance gene. In order to find molecular markers linked with RppQ, and to quickly map the RppQ gene to a specific chromosome, SSR-BSA analysis was carried out. Twenty SSR primer-pairs (phi118, phi041, phi052, umc2114, umc2034, umc2069, umc2016, umc2017, umc2067, umc2043, umc2021, umc1047, umc1648, umc1280, umc1402, umc1507, umc1678, umc1827, umc1898 and umc1569), distributed roughly throughout the ten chromosomes, were screened. Out of them, two primers on chromosome 10, phi118 and phi041, showed linkage with the SCR resistance gene RppQ; the amplification pattern of phi118 was shown in Fig. 1. Because the two linkage markers, phi118 and phi041, are already located on chromosome 10, the RppQ gene should also be located on chromosome 10.

The amplification primer of SSR marker phi118 on 30 susceptible F2 plants from the population of “Qi319×340”. The samples in each lane are: P R resistant parent Qi319; P S susceptible parent 340; B R resistant bulk; B S susceptible bulk; 1–30 F2 susceptible individuals. This is a 3.5% agarose gel stained by ethidium bromide. The star indicates the recombinant
Based on the above results, eight new SSR primer-pairs (umc1293, umc2018, phi117, umc2053, umc1291, umc1318, umc1380 and umc1152) surrounding the region of primers phi118 and phi041 on chromosome 10 were selected, and were further tested regarding their linkage with RppQ. Results revealed that three out of eight SSR primers (umc2018, umc1293 and umc1318) were linked to RppQ. The amplification pattern of umc1293, one of the three, was shown in Fig. 2.

The amplification primer of SSR marker umc1293 on 30 resistant F2 plants from the population of “Qi319×340”. The samples in each lane are: P R resistant parent Qi319; P S susceptible parent 340; B R resistant bulk; B S susceptible bulk; 1–30 F2 susceptible individuals. This is a 6% PAGE gel. Stars indicate the recombinant
Screening AFLP markers linked to RppQ
AFLP-BSA analysis showed that there were about 50–80 bands in each 3+3 primer combination when labeled with [γ-33p] ATP. In the bulked segregant analysis, 54 primer combinations were screened and only one primer combination, E-AGC/M-CAA, amplified polymorphic bands linked with RppQ, which was dominated as AF1 (Fig. 3).

AFLP pattern amplified with primer combination E-AGC/M-CAA on the F2 population. The star indicates the recombinant. The samples in each lane are: P R resistant parent Qi319; P S susceptible parent 340; B R resistant bulk; B S susceptible bulk; 1–15 F2 resistant individuals; 16–30 F2 susceptible individuals. The arrow indicates the specific band AF1
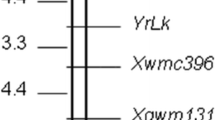
Constructing a local linkage map of the RppQ gene encompassing the region on chromosome 10
Based on above linkage analysis, a total of six markers showed a linkage relation with the RppQ gene. The phenotypic data and the SSR/AFLP data were combined for linkage analysis using the MAPMAKER Vision 3.0 program. Finally a local linkage map of the RppQ gene-encompassing region on chromosome 10 was constructed (Fig. 4). RppQ is located in an interval of 5.79 cM flanked by SSR marker phi041 and AFLP marker AF1 on the short arm of Chr.10, with a genetic distance of 2.45 cM and 3.34 cM respectively (Fig. 4). On the AF1 side, a SSR marker umc1318 is 4.76 cM apart from RppQ. On the phi 041 side, two additional SSR markers, phi118 and umc1293, were linked to RppQ with a genetic distance of 3.33 and 3.78 cM, respectively (Fig. 4).

A partial linkage map of the region surrounding RppQ on the short arm of Chr. 10
Discussion
The exploitation of the new resistant inbred line Qi319 will facilitate maize breeding for SCR resistance
Southern corn rust caused by P. polysora Underw is a broadly distributed maize disease in the whole world. It was not so severe compared with the major disease in the past. However, it has been developing very quickly and widely. Now it becomes one of the major maize diseases in some areas or countries, such as China. It affects maize production both in yield loss and reduced quality. The major reason leading to the rapid spreading of SCR is due to the lack of resistant resources. Most of the inbred lines used in corn breeding, and the commercial varieties used in production, are susceptible to SCR (Rodriguez-Ardon et al. 1980; Ye 2000). Adopting good resistant material for maize SCR breeding will be an effective method to control the quick spreading of SCR. The exploitation of the new resistant inbred line Qi319 will make its contribution to maize SCR resistance breeding due to its high resistance and the simple genetic model for SCR.
Identification and genetic mapping of RppQ to SCR and its marker-assisted selection
SCR is an enormous destructive disease on susceptible maize. So it is desirable to exploit additional sources of resistance against southern rust to improve the disease resistance of present maize hybrids. Moreover, SCR does not occur every year, so general resistance genes could easily be lost in the absence of selection pressure in the maize conventional breeding program. Marker-assisted selection (MAS) promises to be superior to conventional phenotypic selection, if the trait is severely affected by environmental conditions or is difficult to evaluate (Visscher et al. 1996). Localization of genes controlling disease resistance via DNA markers could allow introgression of these genes into elite materials, even in areas where the disease is not common (Hillel et al. 1990; Hospital et al. 1992). In this paper, we identified a dominant resistance gene to SCR in inbred Qi319, which was completely resistant to SCR. SSR and AFLP techniques were carried out to screen markers linked with SCR resistance; as a result five SSR markers and one AFLP marker, which displayed linkage to the RppQ gene, were obtained. The closest linkage markers are AFLP marker AF1 and SSR marker phi041, with the genetic distance away from the RppQ of 3.34 cM and 2.45 cM, respectively. Although they are not close enough to the RppQ gene, they can be used for MAS. Research work is currently in progress to perform fine mapping the RppQ gene with a larger F2 segregation population.
The results reported in this paper clearly demonstrated that the BSA combination with the SSR and AFLP techniques are very powerful, reliable and rapid for molecular marker screening in maize.
Linkage or allelic relationships between RppQ and Rpp9
Genes conferring resistance to different pathogens are often clustered in the same chromosome region of the maize genome (McMullen and Simcox 1995). The short arm of chromosome 10 harbors the resistance genes Rp1 and Rp5, both against common rust (Hulbert and Bennetzen 1991) and Rpp9 (Ullstrup 1965; Holland et al. 1998). Moreover, Rp1, Rp5, Rpp9 are closely linked to each other or allelic. In the present study, a single dominant gene, RppQ, was identified. Linkage analysis demonstrated that RppQ was also located on the short arm of Chr.10, the same region of Rp1. So it was suggested whether, or not, the Rp1, Rpp9 and RppQ, were clustered in the same chromosome region. The research on the allelic relation between RppQ and Rpp9 is undergoing in our laboratory.
References
Akagi H, Yokozeki Y, Inagaki A, Fujimura T (1996) Microsatallite DNA markers for rice chromosomes. Theor Appl Genet 93:1071–1077
Bailey BA, Schun W, Fred-eriksen RA, Bockholt AJ (1987) Identification of slow-rusting resistance to Puccinia polysora in maize inbreds and single crosses. Plant Dis 71:518–521
Chen CX, Yang DE, Yu YJ, Wang ZL, Wang B (2003) Studies on southern corn rust and its resistance. Acta Phytopathol Sin 33:86–87
Chin ECL, Senior ML, Smith JSC (1996) Maize simple repetitive DNA sequences: abundance and allele variation. Genome 39:866–873
Duan DR, He HZ (1984) Description of a rust Puccinia polysora on corn in Hainan Island. Acta Mycol Sin 3:125–126
Futrell MC (1975) Puccinia polysora on maize associated with cropping practice and genetic homogeneity. Phytopathology 65:1041–1042
Futrell MC, Hooker AL, Scott GE (1975) Resistance on maize to corn rust, controlled by a single dominant gene. Crop Sci 15:597–599
Hillel J, Schapp T, Haberfield A, Jeffreys AJ, Plotzky Y, Cahaner A, Lave U (1990) DNA fingerprints applied to gene introgression in breeding program. Genetics 124:783–789
Hoisington DA, Khairallah MM, Gonzales-de-Leon D (1994) Laboratory protocols. CIMMYT Applied Molecular Genetics Laboratory, CIMMYT, Hisfoa, Mexico, DF
Holland JB, Uhr DV, Jeffers D, Goodman MM (1998) Inheritance of resistance to southern corn rust in tropical-by-corn-belt maize populations. Theor Appl Genet 96:232–241
Hooker AL, Yarwood CE (1966) Culture of Puccinia sorghi on detached leaves of corn and Oxalis corniculata. Phytopathology 56:536–539
Hospital F, Chevalet C, Mulsant P (1992) Using markers in gene-introgression breeding programs. Genetics 132:1199–1210
Hulbert SH, Bennetzen JL (1991) Recombination at the Rp1 locus of maize. Mol Gen Genet 226:377–382
Kosambi DD (1944) The estimation of map distance from recombination values. Ann Eugen 12:172–175
Lander ES, Green P, Abrahamson J, Barlow A, Daly MJ, Lincoln SE, Etoh T (1987) MAPMAKER: an interactive computer package for constructing primary genetic linkage maps of experimental and natural populations. Genomics 1:174–181
Liu YY, Wang J (1999) Southern corn rust occurred in Hebei Province in 1998. Plant Protect:53
Lubberstedt T, DuBle C, Melchinger A (1998) Application of microsatellites from maize to teosinte and other relatives of maize. Plant Breed 117:447–450
McMullen MD, Simcox K (1995) Genomic organization of disease- and insect-resistance genes in maize. Mol Plant-Microbe Interact 8:811–815
Melching JS (1975) Corn rust: type, races and destructive potential. In: The American Seed Trade Association, pp 90–115
Rhind D, Waterston JM, Deighton FC (1952) Occurrence of Puccinia polysora Underw in West Africa. Nature 169:631
Rodrigues-Ardon. R, Scott GE, Hennen JF (1980) Maize yield losses caused by southern corn rust. Crop Sci 20:812–814
Scott GE, King SB, Armour JW (1984) Inheritance of resistance to southern corn rust in the maize population. Crop Sci 24:265–267
Ullstrup AJ (1965) Inheritance and linkage of a gene determining resistance in maize to an American race of Puccinia polysora. Phytopathology 55:425–428
Visscher PM, Haley CS, Thompson R (1996) Marker-assisted introgression in backcross breeding programs. Genetics 144:1923-1932
Vos PR, Hogers MB (1995) AFLP: a new technique for DNA fingerprinting. Nucleic Acids Res 23:4407–4414
Wang SL (1999) Southern corn rust was epidemic in the Province of Shandong, Henan and Jiangsu. Plant Protect Technol Dissemin 3:40–41
Ye JC (2000) The first inbred line Qi319 immune to maize southern rust bred in china. Sci Agricult Sin 33:110
Zummo N (1988) Components contributing to partial resistance in maize to Puccinia polysora. Plant Dis 72:157–160
Acknowledgements
This work was supported by the 10th Five-Year Science and Technology Research Program of China (2001BA511B04-01) and the State High-Tech Program of China (2001AA211111).
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by H. F. Linskens
Rights and permissions
About this article
Cite this article
Chen, C.X., Wang, Z.L., Yang, D.E. et al. Molecular tagging and genetic mapping of the disease resistance gene RppQ to southern corn rust. Theor Appl Genet 108, 945–950 (2004). https://doi.org/10.1007/s00122-003-1506-7
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00122-003-1506-7