
Abstract
Recent functional and zooarchaeological studies conducted on the archeological finds of Pirro Nord (PN13) produced new, reliable data on early European hominid subsistence activities. The age of the site is estimated to be ~ 1.3–1.6 Ma, based on bio-chronological data, and the archeological excavation of the Pirro Nord 13 fissure led to the discovery of more than 300 lithic artifacts associated with thousands of vertebrate fossil remains of the final Villafranchian (Pirro Nord Faunal Unit). The analysis of the fossil faunal remains allowed for the identification of anthropogenic traces linked to the exploitation of different animal carcass (cut marks and intentional bone breakages). Use-wear traces were also observed on some flint artifacts and have been interpreted as the result of the exploitation of animal resources by early hominids and carnivores. It has not been possible to identify the type of access that hominins developed on the carcasses, although it has been established that the hominins competed with carnivores for animal resources. The stone tools and faunal remains with anthropogenic traces recovered in the PN13 fissure represent among the earliest evidence of hominin faunal exploitation in Europe.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
The process of human dispersal from Africa started before 2 Ma (Coppens 2017; Han et al. 2017a; Malassé Dambricourt et al. 2016). The oldest evidence of the Eurasian peopling seems to suggest a scenario in which the Asian continent was occupied before Europe. The oldest human evidence comes from China and dates from more than 2 Ma (Boëda and Hou 2011; Han et al. 2017b). Around 1.9–1.6 Ma, human presence is attested to in most of Southern Asia (Dennell 2008a, b; Zhu et al. 2008), in Southeast Asia (Falguères et al. 2016; Swisher et al. 1994), in Northern China (Dennell 2015), and in Georgia (Baena et al. 2010; Gabunia 2000; Garcia et al. 2010; Mgeladze et al. 2010; Vekua 2002).
In Southern Europe, several sites display the arrival of Homo before and around 1 Ma (Álvarez et al. 2015; Arzarello et al. 2007, 2015; Barsky et al. 2010; Bermúdez de Castro et al. 2010; Carbonell et al. 2008; de Lombera-Hermida et al. 2016; De Lumley et al. 1988; Despriée et al. 2006, 2010; Huguet et al. 2017; Lopez-Garcìa et al. 2015; Moyano et al. 2011; Muttoni et al. 2011; Parés et al. 2006), but very few human remains have been discovered and actually attributed to Homo sp. (Bermúdez de Castro et al. 2010; Toro-Moyano et al. 2013).
Among these European archeological sites, Pirro Nord 13 (PN13) represents evidence of the oldest human presence in Europe (Lopez-Garcìa et al. 2015). The site, located inside a karst fissure, holds faunal remains in association with lithic artifacts. The nature and characteristics of the fissure filling allow for the statement that the faunal and archeological remains are in a secondary position and that the sediment, within the bones and lithics, has been transported inside the fissure, mainly by water and gravitational processes (Giusti and Arzarello 2016; Karampatsou 2017). Despite site formation processes, the homogeneity of the deposits and the contemporaneity of the artifacts and faunal remains have been clarified by several works (Bagnus 2011; Giusti and Arzarello 2016; Karampatsou 2017), but the mutual relations between the faunal remains and human activity have never been proved before. The aim of this work is to define the relationship between the faunal remains and the lithic industry of Pirro Nord 13 through zooarchaeological study of the faunal remains and use-wear analysis of the lithic industry.
Historical and background of research
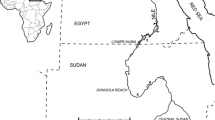
The fossiliferous area of Pirro Nord (also known as Cava Pirro or Cava Dell’Erba) is located at the northwestern margin of the Gargano promontory, close to the village of Apricena (FG, Apulia, Italy; 41°4800700 N, 15°2300500 E) (Fig. 1). The paleontological findings come from several karst fissures situated at the top of a Mesozoic limestone formation that has been the object of exploitation by numerous limestone quarries (Fig. 1). During the Pleistocene, these fissures were part of a very complex interconnected karst system (Pavia et al. 2012). Each fissure containing Villafranchian paleontological remains has been named “P” or “PN,” followed by a progressive number. Paleontological studies have been conducted there since the 1970s (Freudenthal 1971), and systematic field investigations have been carried out by several research teams (De Giuli et al. 1986). In the PN13 fissure, some lithic industry was found, together with vertebrate fossils related to the Pirro Nord Faunal Unit (Arzarello et al. 2009, 2012, 2015; Giusti and Arzarello 2016).

Site location: left—position of Pirro Nord fossiliferous area; right—position of the fissure of PN13 inside the quarry (aerial view)
Geomorphological and stratigraphic settings
The deposit that preserves the paleontological and archeological remains of PN13 fills a vertical karst fissure that was exposed by the mining activities of the limestone quarry. The fracture opens at the stratigraphic boundary between the Mesozoic limestone and the Pleistocene calcarenite formation, in the wilderness zone of a more complex karst system located in the Dell’Erba quarrying complex (Pavia et al. 2012). The deposit is more than 4 m thick and is characterized by a sandy loam matrix with clasts and blocks of local lithology, showing a chaotic arrangement and distribution. On lithological and textural bases, four sedimentary units (SUs) have been distinguished (Fig. 2). Unit A consists of a thin sandy-clay layer that covers the underlying units almost for their entire lateral extension. Its formation was due to vertical percolation of water that led to the deposition of secondary clay at the top of the sequence; the same process filled the empty interstices between the large blocks that characterize the underlying SUs, determining the deposition of clay and fine sand layers. Units B to D are composed of sandy loam sediment among limestone and calcarenite cobbles and boulders, which show poor sorting and a varying degree of sizes and roundness. Units C and D show an abrupt increase in the number and dimension of clasts and large blocks and a significant increase in the number of fossils and lithic artifacts as well (Giusti and Arzarello 2016). The formation of the deposit was due to a first collapse of the top of the fissure, which resulted in communication of the karst system with the ground surface, which determined the fall of the large blocks of calcarenite and limestone. An applied set of spatial analyses (Giusti and Arzarello 2016), along with recent geochemical analyses and geoarchaeological revision of the site (Karampatsou 2017), suggested that the formation of SUs B, C, and D was the result of a gravitational selection of the chaotic materials carried into the fissure during one or more subsequent events of mud flow or earth flow, triggered by humid and arid weathering.

PN13 stratigraphy. a PN13. Stratigraphic layout across squares 1 and 6 (*): labels indicate the stratigraphic units (SUs), and lines mark their bases. b Details of SU B: chaotic structure of many sub-rounded, less than decimetric clasts in sandy muddy matrix. c Details of SU C: chaotic structure of varying sizes of decimetric clasts and boulders in brown-green muddy sand matrix—pin indicates a Mn-rich lens. d Details of SU D: same as c but with larger boulders and common sandy lenses—see the pin—sometimes structured in thin layers with alterations of sand and mud
Considering the stratigraphy and the taphonomical processes, models that describe the formation of the site have been proposed by Bagnus (2011), Berruti (2017), and Giusti and Arzarello (2016). These models assume that animals died closed to the karst sinkhole, and the action of heavy rains transported the faunal remains and lithic artifacts into the fixture, in which they were rapidly buried with mass-wasting processes, such as mud flow or earth flow, also carrying rocks. No post-depositional reworking or mixture of the sedimentary deposit has been observed.
Considering this geomorphological and stratigraphical setting and the re-deposition processes that affected the deposit, the faunal remains and the lithic artifacts found in the PN13 fissure have been studied as a unique assemblage (Arzarello et al. 2009, 2012, 2015; Bagnus 2011; Berruti 2017; Giusti and Arzarello 2016; Lopez-Garcìa et al. 2015).
Faunal remains and biochronology
The PN13 large mammal assemblage is typical of the Pirro Nord Faunal Unit (FU) (De Giuli et al. 1986; Gliozzi et al. 1997; Masini and Sala 2007, 2011). This FU is the last of the Late Villafranchian, and it denotes a first faunal renewal related to the beginning of arid climatic conditions that prevailed during the Galerian. It registers the first occurrence (FAD) of Bison (Eobison) degiulii, Canis (Xenocyon) lycaonoides, Canis mosbachensis, and Equus altidens (Gliozzi et al. 1997; Masini and Sala 2007; Petronio and Marcolini 2013).
The Pirro Nord local fauna is characterized by numerous taxa of reptiles, amphibians, birds, and mammals (Alberdi and Palombo 2013; Angelone 2013; Bedetti 2003; Bedetti and Pavia 2013; Blain et al. 2016; Delfino and Atzori 2013; Delfino and Bailon 2000; Marcolini et al. 2013; Petrucci et al. 2013; Rook and Sardella 2013; Salari et al. 2013). Among large mammals, PN13 is characterized by an assemblage dominated by Pseudodama, cf. P. farnetensis, with, among ungulates, Bison (Eobison) degiulii, Equus altidens, a large size cervid (Praemegaceros?), and Stephanorhinus, cf. S. hunsheimensis. At this site is also present Mammuthus meridionalis and, among carnivores, Canis mosbachensis, Canis, cf. C. (Xenocyon) lycaonoides, Vulpes, cf. V. alopecoides, Pachycrocuta brevirostris, Ursus etruscus, Homotherium sp., Lynx issiodorensis, Felis, cf. F. lunensis, Meles sp., and Martes sp. (Colopi 2016). Among small mammals, the Pirro Nord local fauna is related to the Early Biharian mammal age and the “Mimomys pusillus-Mimomys savini zone” (Gliozzi et al. 1997; Lopez-Garcìa et al. 2015; Masini and Sala 2007). According to the other Pirro Nord localities, the only arvicolid species present at PN13 is Allophaiomys ruffoi. Although no direct dates are available yet, this species has been related to a chronological range between 1.3 and 1.6 Ma, allowing for the hypothesis that Pirro Nord is older than other Western European sites where an ancient human presence has been attested (Lopez-Garcìa et al. 2015).
Lithic assemblage
From 2006 to 2015, 340 lithic artifacts were found in the PN13 fissure related to anthropic activities (231 flakes, 37 cores, and 72 debris) (Table 1). The raw material was collected no more than 7 km from PN13, in secondary positions (riverbeds or slope deposits). The exploited flint pebbles and cobbles have different morphologies and sizes, and they come from the Gargano Cretaceous succession (Arzarello et al. 2009, 2015, 2016; Giusti and Arzarello 2016). In the PN13 lithic assemblage, two main reduction sequences were adopted: an “opportunistic” débitage based on the exploitation of multiple striking platforms (max 5) to produce flakes with different morphologies but always with at least one cutting edge; and a “centripetal” débitage to produce flakes with convergent cutting edges. The “opportunistic” reduction sequence has been adopted for the reduction of largest flint cobbles. This method is an adaptation of the raw material shape to obtain the greatest quantity of blanks with a sharp cutting edge and with the least number of gestures (Arzarello et al. 2015). The “centripetal” reduction sequence could be compared more simply to some S.S.D.A. cores described by Forestier (Forestier 1993)—a “centripetal” debitage for, in most cases, the production of flakes with convergent cutting edges (Arzarello et al. 2015). There is not a clear relationship between the type of flint and the method of débitage adopted for the reduction of the cores, but a relationship between the raw material morphology/size and the knapping method has been underlined (Arzarello et al. 2009, 2015; Giusti and Arzarello 2016). The technique used is always direct percussion by a hard hammer. The retouched instruments are very rare, and only four flakes, issued by an opportunistic method, are characterized by an intentional modification of the cutting edges. All of them were broken, and two show a bending fracture (Arzarello et al. 2009, 2015; Giusti and Arzarello 2016). The macroscopic analysis attests that a good state of preservation characterizes the whole series of artifacts. At this level of analysis, the artifacts have sharp edges and do not show macroscopic evidence of transportation: 35% carry traces of superficial Fe-Mn oxide patinas, as observed on more than 70% of the faunal remains. The only post-depositional alteration identified consists of fractures that affect 20% of the lithic material (Arzarello et al. 2015; Giusti and Arzarello 2016).
Materials and methods
Zooarchaeological study
The material is stored and available for the study at the Dipartimento di Studi Umanistici, Sezione di Scienze preistoriche ed antropologiche of the University of Ferrara under the permission of the Soprintendenza Archeologia, Belle Arti e Paesaggio per le province di Barletta-Andria-Trani e Foggia. Previous works have shown that the PN13 faunal remains are characterized by frequent post-depositional alterations, such as cracks, hydric abrasion (rounding/polishing), carnivores’ modifications, ferromanganese oxide deposition, and chemical alterations (Arzarello et al. 2012, 2015, 2016; Bagnus 2011; Giusti and Arzarello 2016).
The zooarchaeological study was conducted on 1285 remains from the 2012–2015 excavation campaigns; the taxonomic attribution of this sample refers to Colopi (Colopi 2016), except for the remains belonging to the Testudines order, which are under study.
The remains were grouped into five size categories according to the system of classification developed by Bunn (1986) for the Olduvai faunal assemblages: very large size (> 1000 kg), large size (300–1000 kg), medium size (100–300 kg), small size (20–100 kg), and very small size (< 20 kg) (Bunn et al. 1980; Bunn 1986). When possible, the age of death of the animals (age AT death) was defined. This analysis was based on the stages of eruption of the teeth, the dental wear (Ervynck 1997; Grant 1982; Stiner 1998) and the epiphyseal fusion stages (Brown and Chapman 1991; Eisenmann 1979). The following indices were calculated: NISP (number of identified specimens), MNE (minimum number of skeletal elements), MNI (minimum number of individuals), and %MAU (minimum number of animal units) (Binford 1981; Lyman 1994).
The study of the sample was conducted to detect all taphonomic modifications, paying special attention to hominin and carnivore modifications (cut marks, bone breakage, and tooth marks); these elements were analyzed according to the works of different scholars (Andrews 1995; Behrensmeyer 1978; Cáceres 2002; Fernández-jalvo and Andrews 2003; Fernández-Jalvo and Andrews 2016; López-González et al. 2006; Marín-Arroyo et al. 2014). Butchery traces, with striations of variable length and width, were analyzed using the criteria described by Binford (1981), Shipman and Rose (1983), Nilssen (2000), Domínguez-Rodrigo et al. (2009), Domínguez-Rodrigo et al. (2017), and Soulier and Costamagno (2017).
A bone breakage study was conducted using the method developed by Villa and Mahieu (1991), which considers the type of fracture, the angle, and the edge texture. To discern the breakage traces linked to anthropic activity, the presence of percussion notches and impact flakes, in accordance with the works of Capaldo and Blumenschine (1994) and Pickering and Egeland (2006), was considered. Carnivore tooth marks (pits and scores) were analyzed in view of the type of the tooth marks, the location of these marks on the bones, the type of the bone tissue affected (shaft or end), and the dimensions (length and breadth) of each tooth mark (Delaney-Rivera et al. 2009; Domínguez-Rodrigo and Barba 2006; Domínguez-Rodrigo and Piqueras 2003; Pobiner 2015; Pobiner et al. 2008; Saladié et al. 2014). The zooarchaeological study was conducted with the aid of two different stereomicroscopes: Optika (SZM-2 EWF × 10/× 20, magnification × 0.7–× 4.5) for all of the material and a Hirox digital microscope (K-H 8700) for the study of butchery traces with striations of variable length and width.
Use-wear analysis
This study began with the preliminary evaluation of the entire artifact sample found in the fissure of PN13 during the excavation campaigns from 2006 to 2015 (340 artifacts). Two criteria were applied to select artifacts for the use-wear analysis: presence of at least one useful edge (selected according to the criteria developed by Terradillos-Bernal and Rodríguez-Álvarez (2017); and surface preservation (absence of marked PDSMs (post-depositional surface modifications) (Levi Sala 1986)). The PN13 evaluated sample is finally composed of 63 flint flakes (15 centripetal and 48 opportunistic flakes). For the analysis, each selected artifact was gently washed with warm water and soap, and then it was washed for 3 min in a mixture of demineralized water (75%) and alcohol (25%) in an ultrasonic tank and was open air dried. The use-wear analysis of the PN13 assemblage was conducted with an integrated approach that uses a low power approach (Odell and Cowan 1986) in combination with a high power approach (Keeley 1980). Several works (Berruti and Daffara 2014; Beyries 1987; Burbidge et al. 2014; Cruz and Berruti 2015; Hardy 2004; Lemorini et al. 2014a, b; Moss 1983; Van Gijn 2014; Visentin et al. 2015; Wilkins et al. 2015) have shown, in fact, that the use of both methodologies integrated is more effective and productive.
The analysis of the macro-traces, or the low power approach, provides information about the potential activities carried out (e.g., cutting, scraping, piercing, etc.) and general interpretation of the hardness of the worked materials (Lemorini et al. 2006; Lemorini et al. 2014a; Odell 1981; Semenov 1964; Tringham et al. 1974). The analysis of the micro-traces, or the high power approach, was conducted to provide a more detailed understanding of the activities carried out with the lithic artifacts and to support the diagnosis of the processed materials (Keeley 1980; Lemorini et al. 2006; Lemorini et al. 2014a; Rots 2010; Van Gijn 2014; Ziggioti 2005). The analysis of the lithic artifacts was conducted using three different types of microscope: a Seben Incognita III stereoscopic microscope with magnification from × 20 to × 80 (for the first scanning of the collection), a Leica EZ4 HD stereoscopic microscope with magnification from × 8 to × 40 (for the low power approach analysis) and an Optika B 600 Met metallographic microscope supplied with 5 PLAN IOS MET objectives (× 5–10–20–50–100) (for the high power approach analysis). Adobe Photoshop CS6 Portable (© Adobe) software was used for the treatment of the images since it allows a single image to be built up from several photos taken at different depths of field. To better identify the traces of use present on the artifacts, a reference collection with flakes made of the same raw material used in Pirro Nord was produced (it was also used for other papers (Buccheri et al. 2016)). Several specific activities were then conducted on different materials (skinning, filleting, woodworking, etc.) with the experimental lithic tools to link the use-wear features to tool motions and to the processed materials. In particular, 10 tools were used to carry out butchering activities on wild boar and rabbit carcasses. During the experimentation, un-retouched flakes obtained using the S.S.D.A. method and the hard percussion technique were used. For each of them, the time of use, the direction of the gesture, and the material worked were recorded. After the experiments, each artifact was gently washed with warm water and soap, and then it was washed for 3 min in a mixture of demineralized water (75%) and alcohol (25%) in an ultrasonic tank and was open air dried. Finally, each object was studied using the same method used for the archeological collection.
Results
Zooarchaeology results
Of the 1285 fossil remains present in the sample, 482 were identified anatomically and taxonomically, likely due to the condition of the bones and, in particular, the high fragmentation rate. The large mammal assemblage is coherent with the associations found in other Pirro Nord localities, and it is not contaminated by faunal remains coming from Quaternary karstic fissures, while in the small mammal assemblage, three remains related to the Terre Rosse endemic assemblages (Messinian-Zanclean) have been detected (Colopi 2016; Lopez-Garcìa et al. 2015).
During the study, the remains were divided into five size categories. The very large size regroups adult Mammuthus meridionalis and Stephanorhinus, cf. S. hundsheimensis. The large size contains adult Equus altidens, Bison (Eobison) degiulii, Ursus etruscus, Homotherium sp., young Stephanorhinus, cf. S. hundsheimensis, and Homotheirum sp. In the medium size group were placed young Equus altidens, Ursus etruscus, and Praemegaceros and adults of Pachycrocuta brevirostris and Pseudodama, cf. P. fartenensis. The small size group includes young of Pachycrocuta brevirostris, Canis mosbachensis, Hystrix refossa, Pseudodama, cf. P. farnetensis, and Lynx issiodirensis, and adult of Canis cf., C. (Xenocyon) lycaonoides. Finally, the very small size group contains young Canis, cf. C. (Xenocyon) lycaonoides, Meles sp., Martes sp., Vulpes, cf. V. alopecoides, Felis, cf. F. lunensis, and Testudines. Despite taxonomic variability, the MNIs (Table 2) shows the predominance of fallow deer with 12 individuals, while the other genera present in the sample do not exceed an NMI of 7.
Related to division for age groups, considering the entire sample (Table 2), the finds attributable to adult specimens are the most represented, with 29 individuals, while the juveniles are 15, the infantile are 9, and the senile are 10. In terms of skeletal representation, appendicular and cranial elements are the most common, while axial parts are relatively rare (Table 3).
The PN13 assemblage is composed of different sizes of ungulates, mainly deer, equines, and other sizes of very large animals, such as rhinoceroses and proboscideans. Anatomically, all of the segments were conserved in this set, and the appendicular skeleton is best represented, followed by cranial and axial segments. Deer (Pseudodama farnetensis and Praemegaceros) had more complete skeletal representation. Isolated teeth are best represented, followed by unidentifiable long bones, metapods, vertebrae, phalanges, ribs, tarsals, tibiae, mandibles, skulls, femurs, humeri, radii, scapulae, antlers, pelvises, ulnae, carpals, patellae, sterna, and horns. The majority of MNE are represented by large animals, followed by medium-sized, small-sized, very small-sized, and finally very large-sized animals. According to %MAU (Table 4), a biased representation was observed in all of the taxa in the assemblage. The best-represented elements of larger animals are those belonging to the medial and proximal appendicular segments, followed by the axial and cranial, with relatively high proportions. The well-represented elements in the medium-sized category are those inherent to the medial appendicular skeleton, followed by axial and cranial ones. The small-sized animals are greatly represented by elements appertaining to the axial segment, followed by the cranial and appendicular ones, with small proportions of representation.
The PN13 faunal remains assemblage is affected by several taphonomic modifications: manganese oxide patina (97.12%), rounding (57.27%), polishing (33.23%), iron oxides (51.83%), and root-etching marks (5.83%) (Fig. 3).

Taphonomic alterations observed on the analyzed fossils of Pirro Nord 13
Despite the post-depositional modifications, evidence of human activities (cut marks and breakage patterns) and that of carnivores (tooth marks) were identified in this assemblage, although hominin faunal exploitation evidence is less abundant than carnivore activity. During the study, 14 cut-marked bones were identified (1.08%) (Table 5). This analysis was based on the diagnostic criteria of Domínguez-Rodrigo (Domínguez-Rodrigo et al. 2009; Domínguez-Rodrigo et al. 2012; Domínguez-Rodrigo and Yravedra 2009; Egeland 2012; Merritt 2012), which were used to precisely characterize these cut marks and to distinguish them from those produced by taphonomic processes (such as trampling and weathering): location, orientation, trajectory of the groove, and the presence or absence of micro-striations inside the groove, the shape of the groove (narrow V or wide V), and the presence of flaking on the shoulder. All of them are slicing marks, with lengths varying between 21.42 and 1.36 mm. The majority are obliquely oriented in relation to the major axis of the bone and only in few cases are transverse. A cluster of obliquely oriented cut marks were found on proximal metaphysis of a left metatarsal, previously recognized as unidentified (Colopi 2016) and now attributed to cf. Equus (12D17B4S66). This bone is not well preserved, and it lacks any secure morphological features that allow for species attribution. Nevertheless, part of the small facet for the mesentocuneiform (Hussain 1975) is present, and on the proximal diaphysis portion, a typical Perissodactyla transverse incision for tendon attachment is visible (Fig. 4). On two shaft metatarsal bones of Pseudodama, cf. P. farnetensis, on one Praemegaceros diaphysis shaft metatarsal and on one lumbar vertebra of Pseudodama were found cut marks obliquely oriented (Fig. 5). Three other cut marks are situated on three different rib diaphyses attesting to transverse and oblique orientation (Fig. 6). Finally, four obliquely oriented incisions are identified on four long bone shafts.

Cut marks: PN13, PN13-D17B4S66; 1 and 2, several oblique incisions, grouped on a left metatarsal of Equus; 3, carnivore tooth mark (score)

Cut marks. a PN13-D22B2S69, linear incision, oblique on the lateral dorsal part of the body of a lumbar vertebra of Pseudodama, cf. P. farnetensis. b PN13-D22B1 (1-1), linear, oblique and deep incision on the diaphysis of a metatarsal of Pseudodama

Cut marks. a PN13-RIM 1-2, two incisions, parallel, oblique scutted by the edge of the fracture on the diaphysis of a long bone of a medium-sized animal. b PN13-D19B5S73, two incisions, parallel, obliques on limb mid-shaft of a medium-sized animal. c PN13-D19D1S05, cut mark, oblique linear on the lateral part metatarsal of Praemegaceros
During the analysis of the sample evidence, intentional bone breakage documented on eight remains was found (Fig. 7): two fragments of limb bone shafts with percussion notches (of two medium-sized animals); four impact flakes, two of them referable to a medium-sized animal (metatarsal bone and humeri of deer); and another one referable to large-sized animal.

Bone breakage evidence. a PN13-D24C2-34, impact flake (large-sized animal). b PN13-D24C2-16, impact flake (medium-sized animal). c PN13-D21B2S59 limb mid-shaft showing percussion notches (medium-sized animal). d PN13-D22B2S70, proximal epiphysis+diaphysis of deer metatarsal bone demonstrating flake parasitism by percussion. e PN13-D18B3S93, limb mid-shaft large-sized animal with percussion notches
On the faunal remains, several carnivore tooth marks (pits and scores) were observed (5.56% of the whole sample) (Table 6 and Figs. 8 and 9). The remains referable at the appendicular skeleton (stylopodial and zeugopodial) are those that appear more affected by the carnivore tooth marks. The score dimensions show a range of length of 7.61 mm and a breadth of 1.1 mm, while those of the pits are 3.7 mm for the length and 3.31 mm for the breadth.

Carnivore modifications. a PN13-D19C4S116, score on long bone metaphysis (large-sized animal). b PN13-RIM 1-2-2015, Pit on bone fragment (small-sized animal). c PN13-C11B2S02, pit on the dorsal part of a vertebra

Carnivore modifications. a PN13-D20C4S125. b PN13-C10B5S82, cervical vertebrates (medium-sized animal) showing consumption traces. c PN13-D22B1S84, digested fragment of metapodial diaphysis (medium-sized animal). d PN13-D22C6S184, hyena coprolith. e PN13-D23C5S119, femur mid-shaft (medium-sized animal) presenting pitting
There is also a cut-marked bone (equid metatarsal) that preserves cut marks associated with one score mark made by a carnivore tooth (Fig. 4).
The results obtained analyzing the epiphyses/diaphysis ratio and the percentage of deterioration show a highly significant negative correlation, indicating intense modification caused by carnivores at PN13, although in the case of medium-sized remains, this percentage is low (Table 6).
Use-wear analysis results
The microscopic analysis of the surfaces of the lithic finds, conducted with the aid of a stereomicroscope, showed that 296 of the 340 lithic artifacts studied present post-depositional alterations (PdAs) or PDSMs (Levi Sala 1986): 259 edges crumbling, 208 Fe-Mn oxide patinas, 190 rounding of edges and ridges, 277 polished surfaces, and 58 white patinas. The different PDSMs registered on the lithic artifacts have different intensities, combinations and distributions (Fig. 10).

PDSM alteration on a PN 14 Beta 2007. a On this surface, it is possible to see the presence of white patina (W), polished patina (P), rounding of the edge (R), and Fe-Mn patina (F). b Post-depositional polish on the edge of the artifact
Therefore, only 63 artifacts have both criteria (useful edge and surface preservation) needed to be suitable for use-wear analysis.
Among the 63 artifacts selected for the use-wear analysis, only five showed use-wear traces (Fig. 11). They are medium- and small-sized flakes, measuring between 20 and 43 mm in length, 30 and 15 mm in width, and 6–15 mm thick. Four of them show only one used edge, while the other has two active edges. In total, five different used flakes have been found with six functional edges. Some of the functional edges identified present edge removals not due to intentional retouching but rather micro-flaking produced during use (Odell 1981; Semenov 1964). The main difference between the traces of use and post-depositional edge removals is the combination of the trace attributes; the contact with the worked material produces specific combinations of attributes, which rarely are replicated by post-depositional agents (Lemorini et al. 2014a). In any case, considering the antiquity of the site and the formation process of this archeological deposit, only the edges that presented edge removals in association with polishes (Keeley 1980) were considered used. The use wear traces identified on the edges of the artifacts are sufficiently developed to determine the kinematics of the tool use (motion) and the type of the worked material with a satisfactory degree of reliability. Small, diagonally oriented edge removals (attributable at cutting motions) were recognized on the four functional edges belonging to the flakes with only one functional edge, and all of them also presented polishes linked to butchering activities. The polishes identified affect the edge surfaces gradually, and their extension decreases from the outside to the inner part; furthermore, the width of these polishes on the faces of the edges is limited. The limited polish extension on the surfaces usually indicates that the position of the tool relative to the worked material was characterized by a high angle of work. The longitudinal motion and a high angle of work are typical features of cutting actions, which are also typical of butchering activities. Part of the recorded polishes on the edges appears like a rough band associated with, in some cases, diagonal smears that could be the result of contact with soft animal tissue. Furthermore, on the same edge surfaces, there are areas showing small, smooth, and flat spots of polish that could be linked to contact with bone (Fig. 11). On the last two functional edges identified were recognized sub-perpendicular and diagonally oriented edge removals and rough bands of polishes with, in some cases, sub-perpendicular smears, likely linked to mixed actions on soft animal tissue (Fig. 11). Similar characteristics are found on the margins of the instruments used during the experimentation to perform butchering actions on rabbit and wild boar carcasses (Fig. 12), as well as the data presented in numerous scientific works dealing with the same type of traces (e.g., Lemorini et al. 2006; Lemorini et al. 2016; Iwase 2016; Pawlik 2013; Solodenko et al. 2015; Lemorini et al. 2014b).

Use wear traces on PN13 artifacts: 13ϒ2007 (scale bar 1 cm): (a) micro-use wear interpreted as the result of a mixed motion (× 30); (b, c) details of the edge area (× 100, × 200) with a band of rough polish (contact with fleshy tissues). 19ε2007 (scale bar 1 cm); (a) micro-use wear interpreted as the result of a mixed motion (× 30); (b) details of the edge area (× 100) with a band of rough polish (contact with fleshy tissues?). 140B4C72011 (scale bar 1 cm); (a) micro-use wear interpreted as the result of a longitudinal motion (30x); (b, c) details of the edge area (× 100, × 200) with rough polish (contact with fleshy tissues). S107C6D212012 (scale bar 1 cm); (a) details of the area in the square (× 100) with rough polish (contact with fleshy tissues); (b) details of the area in the square (× 200), with rough polish (contact with fleshy tissues). S11B3A2008 (scale bar 2 cm); (a) micro-use wear interpreted as the result of a mixed motion of butchering (× 50); (b; c) details of the area in the square (× 100, × 200) with small, smooth, and flat spots of polish (contact with bone)

Use wear traces on experimental artifacts: Sper. 8 (scale bar 1 cm): (a) this use-wear is the result of transversal motion during a butchering activity on a wild boar carcass (× 30); (b, c) details of the edge area (× 100, × 200) with small, smooth and flat spots of polish due to contact with bone. Similar to the traces found on the artifact S11B3A2008. Sper.5 (scale bar 1 cm): (a) this use wear is the result of longitudinal motion during a butchering activity on a rabbit carcass (× 30); (b, c) details of the edge area with a band of rough polish (× 100, × 200) by contact with fleshy tissues. Similar to the traces found on the archeological artifacts 13ϒ2007, 19ε2007, 140B4C2011, and S107C6D212012
Discussion
The available data about the first European peopling are not very numerous, but every little piece of information is fundamental to understanding human behavior. As already mentioned, Europe was peopled for the first time before 1 Ma. Some of the oldest sites can give us data only about technical behavior since the bones are not preserved (Peretto et al. 1998a, b; Despriée et al. 2006), but some others, including Pirro Nord 13, allow us to also have information about hominid-carnivore interactions. Because PN13 is not a primary deposition site, the archeological and paleontological assemblage can reflect different origins (Colopi 2016; Giusti and Arzarello 2016; Lopez-Garcìa et al. 2015). The presence of several types of post-depositional alterations on the bones and lithic surfaces confirms that the remains were transported from their original place into the fissure (Arzarello et al. 2012, 2015; Bagnus 2011; Berruti 2017; Giusti and Arzarello 2016). Therefore, in the fissure were collected faunal remains and lithics in relation to each other and findings that have only a chronological relation to each other. All of the materials contained in the PN13 fissure, lithic artifacts and fauna remains, are related to the same time span that can be placed at the end of the Villafranchian (Colopi 2016; Pavia et al. 2012; Bagnus 2011; Berruti 2017; Giusti & Arzarello 2016). Indeed, the paleontological and geoarchaeological studies confirmed that this period was the only time when the material in the fissure accumulated (Berruti 2017; Colopi 2016; Giusti and Arzarello 2016; Karampatsou 2017; Lopez-Garcìa et al. 2015). The relation between fauna and lithics is demonstrated only for a reduced part of the assemblage (Bagnus 2011; Giusti and Arzarello 2016). For these reasons, the analyses conducted on the PN13 remains are mainly qualitative, while the quantitative aspects of these studies have relative meaning. Regarding the use-wear analysis of the lithic assemblage, the only activity that it was possible to identify on the PN13 lithic artifacts is the exploitation of animal resources to obtain soft animal tissues, e.g., muscle, viscera, brains, and marrow. Indeed, the presence of two different types of polishes on the same edge, one linked to contact with bone and the other linked to contact with fleshy tissues, can be related to meat extraction from the animals’ carcasses. The presence of this type of use-wear traces on lithics tools referable to ancient sites is not new; indeed, use-wear traces referable to butchering activities have been found on artifacts from many of the oldest African sites: Kanjera South (± 2.0 Ma, Kenya (Lemorini et al. 2014a), Aïn Hanech (± 1.78 Ma, Algeria, (Sahnouni et al. 2013), and Koobi Fora at ± 1.5 Ma (Keeley and Toth 1981). The presence of use-wear traces linked to butchering activities on lithic tools has also been attested in many ancient European and Asian sites: Monte Poggiolo (Longo 1994; Peretto et al. 1998a, b), level TD6 of Gran Dolina (Carbonell et al. 1999) and Xiaochangliang (Shen and Chen 2000).
Despite the post-depositional modifications and numerous carnivore tooth marks, evidence of human activities (cut marks and breakage patterns) has been recognized in the considered faunal assemblage. All of the cut marks identified are slicing marks discovered on medium and large-sized animals’ bones, mainly equids and cervids. According to the criteria developed by Binford (1981), Nilssen (2000), and Soulier and Costamagno (2017), all of the identified cut marks can be linked to the realization of two different slaughtering activities: defleshing and skinning. The presence of traces linked to intentional bone breakage (such as percussion notches and impact flakes) might testify to the practice of marrow extraction. Interaction between carnivores and hominins at PN13 is demonstrated by a cut-marked bone (equid metatarsal) that preserves cut marks associated with one score mark made by a carnivore (Egeland 2007). Even if different methods to identify the access type (i.e., primary or secondary access by hominids) have been applied (Blasco and Rosell 2009; Blumenschine and Selvaggio 1998; Capaldo 1995; Domínguez-Rodrigo and Barba 2006; Egeland et al. 2004, 2014; Selvaggio 1998) in the considered faunal assemblage, overlapping of cut marks and teeth marks, as well as evidence of anthropic evisceration on the faunal remains, is not present. However, the combined evidence of butchering activities on metapodial parts, axial skeletons, the absence of cut marks on stylopodial and zeugopodial limb mid-shafts (Domínguez-Rodrigo and Barba 2006; Domı́nguez-Rodrigo 1997; Egeland 2012; Galán and Domínguez-Rodrigo 2013; Merritt 2012; Nilssen 2000; Soulier and Costamagno 2017; Val et al. 2017) and the presence of impact flakes and percussion notches on limb mid-shafts (Blumenschine and Selvaggio 1998; Capaldo and Blumenschine 1994; Domı́nguez-Rodrigo 1997; Gaudzinski 1993, 2004; Pickering and Egeland 2006; Yeshurun et al. 2011), seem to attest to secondary access to the carcasses by the hominids of PN13 (Gaudzinski 1993, 2004; Yeshurun et al. 2011). Nevertheless, we do not completely exclude the primary access because of the presence of cut marks mid-shaft on unidentifiable long bones.
Conclusions
The scarcity of faunal remains with anthropic and carnivore marks and the nature of the PN13 fissure do not permit determining with confidence the type of access to the carcasses, although it has been established that hominins competed with carnivores for animal resources. Given the small number of lithic tools with use-wear traces and bone with cut marks, for this assemblage, it was not possible to correlate the results.
Not forgetting the available data and remembering that the evidence is not numerous in any of the oldest European sites, it seems that human behavior was not the same for all sites. Around 1 Ma, human groups preferred primary, secondary and mixed access to animal carcasses. Abundant cut marks have been found on the large mammal bones of Fuente Nueva-3 and Barranco León, and opportunistic scavenger behaviour has been underscored for the hominin populations (Rodríguez-Gómez et al. 2016). In Bois-de-Riquet, anthropic marks have been identified for two long bone shafts, and probable secondary access to the carcass has been supposed (Bourguignon et al. 2016). In Level TE9c of Sima del Elefante, cut marks and breakages have been found on Equidae, Bovidae, and Cervidae specimens; even in this case, skinning, defleshing, and anthropic breakage are the recognized activities. Unlike what is observed for the other sites, in Sima del Elefante, the hominins had primary access to the prey (Huguet et al. 2017). The Vallonnet cave is another example of primary access to herbivores of middle size, but in the site, secondary access is also documented for larger animals and, in particular, for Bison schoetensacki (Echassoux 2009).
In conclusion, due to the nature of the accumulation, we cannot assume that all specimens represent a single event; moreover, the post-depositional alteration made it difficult to obtain exhaustive data. Of the approximately 1200 fossils analyzed, a small number (not much more than 1%) present cut marks, most likely made during skinning, meat removal and breakage of the bones. The use-wear analysis, conducted on 300 lithic specimens, allowed for identifying only 5 artifacts that preserved traces (important data for a such old site) and were used during removal of soft tissues. These findings indicate a similar trend to the archaeozoological study and generally with the actual knowledge about this topic, i.e., that early members of the genus Homo were meat consumers. However, in the case of Pirro Nord, whether they had primary (hunters) or secondary (scavengers) access to carcasses cannot be determined.
In Europe, around 1 Ma, we have a homogeneous substratum for what concerns lithic production, but we have some peculiarities, for example, the shaped elements, the retouched blanks and the utilization of centripetal debitage methods. It is very difficult to define a detailed picture when we face numerically reduced evidence, but at the current state of knowledge, we can certainly underscore how human behavior during the first European peopling was strongly influenced and conditioned by the type and availability of resources.
Change history
28 July 2020
In the original publication of this article, one of the author names was incorrectly captured. The first name should be Razika, then family name should be Chelli���Cheheb.
References
Alberdi MT, Palombo MR (2013) The Early Pleistocene Equidae from Pirro Nord (Apricena, Southern Italy). Palaeontographica Abt A: Palaeozoology - Stratigraphy 298(1–6):147–167
Álvarez C, Parés JM, Granger D, Duval M, Sala R, Toro I (2015) New magnetostratigraphic and numerical age of the Fuente Nueva-3 site (Guadix-Baza basin, Spain). Quat Int 389:224–234. https://doi.org/10.1016/j.quaint.2015.04.044
Andrews P (1995) Experiments in Taphonomy. J Archaeol Sci 22(4):147–153
Angelone C (2013) Fossil Lagomorpha from Pirro Nord (Apulia, southern Italy). Palaeontogr Abt A 298(1–6):95–105. https://doi.org/10.1127/pala/298/2013/95
Arzarello M, Marcolini F, Pavia G, Pavia M, Petronio C, Petrucci M, Rook L, Sardella R (2007) Evidence of earliest human occurrence in Europe: the site of Pirro Nord (southern Italy). Naturwissenschaften 94:107–112
Arzarello M, Marcolini F, Pavia G, Pavia M, Petronio C, Petrucci M, Rook L, Sardella R (2009) L’industrie lithique du site Pléistocéne inférieur de Pirro Nord (Apricena, Italie du sud): une occupation humaine entre 1,3 et 1,7 Ma. Anthropologie 113(1):47–58. https://doi.org/10.1016/j.anthro.2009.01.004
Arzarello M, Pavia G, Peretto C, Petronio C (2012) Evidence of an Early Pleistocene hominin presence at Pirro Nord (Apricena, Foggia, southern Italy): P13 site. Quat Int 267:56–61. https://doi.org/10.1016/j.quaint.2011.01.042
Arzarello M, Peretto C, Moncel MH (2015) The Pirro Nord site (Apricena, Fg, southern Italy) in the context of the first European peopling: convergences and divergences. Quat Int 389:255–263. https://doi.org/10.1016/j.quaint.2014.08.051
Arzarello M, De Weyer L, Peretto C (2016) The first European peopling and the Italian case: Peculiarities and “opportunism.”. Quat Int 393:41–50. https://doi.org/10.1016/j.quaint.2015.11.005
Baena J, Lordkipanidze D, Cuartero F, Ferring R, Zhvania D, Martín D et al (2010) Technical and technological complexity in the beginning: the study of Dmanisi lithic assemblage. Quaternary International Oldest Human Expansions in Eurasia: Favouring and Limiting Factors 223–224:45–53
Bagnus, C. (2011). Analisi tafonomica delle associazioni a vertebrati del Pleistocene inferiore di Pirro Nord. Turin
Barsky D, Celiberti V, Cauche D, Grégoire S, Lebègue F, de Lumley H, Toro-Moyano I (2010) Raw material discernment and technological aspects of the Barranco León and Fuente Nueva 3 stone assemblages (Orce southern Spain). Quat Int 223–224:201–219. https://doi.org/10.1016/j.quaint.2009.12.004
Bedetti C (2003) Le avifaune fossili del Plio-Pleistocene italiano: sistematica, paleoecologia ed elementi di biocronologia. Università “La Sapienza” Roma
Bedetti C, Pavia M (2013) Early Pleistocene birds from Pirro Nord (Puglia, southern Italy). Palaeontographica Abt. A: Palaeozoology - Stratigraphy 298:31–53. https://doi.org/10.1127/pala/298/2013/31
Behrensmeyer AK (1978) Taphonomic and ecologic information from bone weathering. Paleobiology 4(2):150–162
Bermúdez de Castro JM, Martinón-Torres M, Robles AG, Prado L, Carbonell E (2010) New human evidence of the Early Pleistocene settlement of Europe, from Sima del Elefante site (Sierra de Atapuerca, Burgos, Spain). Quat Int 223–224:431–433. https://doi.org/10.1016/j.quaint.2009.07.024
Berruti GLF (2017) Use-wear analysis of discoid-conception lithic industries. Universidade de Trás-os-Montes e Alto Douro, Vila Real
Berruti GLF & Daffara S (2014) Theory and methods of the use wear analyisis of the lithic tools. The example of the arrowhead from the Morgado superior Cave (Tomar, PT). Antrope, 221–242
Beyries, S. (1987). Variabilité de l’industrie lithique au Moustérien: approche fonctionnelle sur quelques gisements français. Oxford: British Archaeological Reports International Series 328
Binford LR (1981) Bone ancient men and modern myths. Academic Press, London
Blain H-A, Delfino M, Berto C, Arzarello M (2016) First record of Pelobates syriacus (Anura, Amphibia) in the early Pleistocene of Italy. Palaeobiodiversity and Palaeoenvironments 96(1):111–124. https://doi.org/10.1007/s12549-015-0220-1
Blasco R, Rosell J (2009) Who was the first? An experimental application of carnivore and hominid overlapping marks at the Pleistocene archaeological sites. Comptes Rendus Palevol 8(6):579–592
Blumenschine RJ, Selvaggio MM (1998) Percussion marks on bone surfaces as a new diagnostic of hominid behaviour. Nature 34(6):565–607. https://doi.org/10.1006/jhev.1998.0216
Boëda É, Hou Y-M (2011) Analyse des artefacts lithiques du site de Longgupo. L’Anthropologie 115(1):78–175. https://doi.org/10.1016/j.anthro.2010.12.005
Bourguignon L, Crochet J-Y, Capdevila R, Ivorra J, Antoine P-O, Agustí J, Barsky D, Blain HA, Boulbes N, Bruxelles L, Claude J, Cochard D, Filoux A, Firmat C, Lozano-Fernández I, Magniez P, Pelletier M, Rios-Garaizar J, Testu A, Valensi P, de Weyer L (2016) Bois-de-Riquet (Lézignan-la-Cèbe, Hérault): a late Early Pleistocene archeological occurrence in southern France. Quat Int 393:24–40. https://doi.org/10.1016/j.quaint.2015.06.037
Brown WAB, Chapman NG (1991) The dentition of red deer (Cervus elaphus): a scoring scheme to assess age from wear of the permanent molariform teeth. J Zool 224(4):519–536. https://doi.org/10.1111/j.1469-7998.1991.tb03783.x
Buccheri F, Bertè DF, Luigi G, Berruti F, Cáceres I, Volpe L et al (2016) Tapphonomic analysis on fossil remains from the Ciota Ciara (Piedimont, Italy) and a new evidence of cave bear and wolf exploitation with simple quartz flakes by Neanderthal. Research in Paleontology and Stratigraphy 122(November):41–54
Bunn HT (1986) Patterns of skeletal representation and hominid subsistence activities at Olduvai Gorge, Tanzania, and Koobi Fora, Kenya. J Hum Evol 15(8):673–690. https://doi.org/10.1016/S0047-2484(86)80004-5
Bunn H, Harris JWK, Isaac G, Kaufulu Z, Kroll E, Schick K et al (1980) FxJj50: an Early Pleistocene site in northern Kenya. Taylor & Francis, Ltd., World Archaeology. https://doi.org/10.2307/124398
Burbidge CI, Trindade MJ, Dias MI, Oosterbeek L, Scarre C, Rosina P, Cruz A, Cura S, Cura P, Caron L, Prudêncio MI, Cardoso GJO, Franco D, Marques R, Gomes H (2014) Luminescence dating and associated analyses in transition landscapes of the alto ribatejo, Central Portugal. Quat Geochronol 20:65–77. https://doi.org/10.1016/j.quageo.2013.11.002
Cáceres I (2002) Tafonomía de yacimientos antrópicos en Karst. In: Complejo Galería (Sierra de Atapuerca, Burgos), Vanguard Cave (Gibraltar) y Abric Romaní (Capellades, Barcelona). University of, Tarragona
Capaldo SD (1995) Inferring hominid and carnivore behavior from dual patterned archaeofaunal assemblages. Rutgers University
Capaldo SD, Blumenschine RJ (1994) A quantitative diagnosis of notches made by hammerstone percussion and carnivore gnawing on bovid long bones. Am Antiq 59(04):724–748. https://doi.org/10.2307/282345
Carbonell E, García-Antón MD, Mallol C, Mosquera M, Ollé, a, Rodríguez, X. P., et al. (1999) The TD6 level lithic industry from Gran Dolina, Atapuerca (Burgos, Spain): production and use. J Hum Evol 37:653–693. https://doi.org/10.1006/jhev.1999.0336
Carbonell E, Bermúdez de Castro JM, Parés JM, Pérez-González A, Cuenca-Bescós G, Ollé A, Mosquera M, Huguet R, van der Made J, Rosas A, Sala R, Vallverdú J, García N, Granger DE, Martinón-Torres M, Rodríguez XP, Stock GM, Vergès JM, Allué E, Burjachs F, Cáceres I, Canals A, Benito A, Díez C, Lozano M, Mateos A, Navazo M, Rodríguez J, Rosell J, Arsuaga JL (2008) The first hominin of Europe. Nature 452(7186):465–469. https://doi.org/10.1038/nature06815
Colopi F (2016) Le faune villafranchiane di Pirro 13 (Apricena, Puglia). Università di Ferrara
Coppens Y (2017) Les derniers préhumains et les premiers humains. Comptes Rendus Palevol 16(2):196–199. https://doi.org/10.1016/J.CRPV.2016.04.001
Cruz, A., & Berruti, G. L. F. (2015). Use-wear analysis of the knapped lithic grave goods from Gruta Do Morgado Superior ( Tomar , Portugal ), (October), 81–94
De Giuli F, Masini F, Torre D (1986) The latest Villafranchian faunas in Italy: the Pirro Nord Fauna (Apricena, Gargano). Palaeontographia Italica 74:51–62
de Lombera-Hermida A, Rodríguez-Alvarez X-P, Peña L, Sala Ramos R, Despriée J, Moncel M-H et al (2016) The lithic assemblage from Pont-de-Lavaud (Indre, France) and the role of the bipolar-on-anvil technique in the Lower and Early Middle Pleistocene technology. J Anthropol Archaeol 41:159–184. https://doi.org/10.1016/j.jaa.2015.12.002
De Lumley H, Fournier A, Krzepkowska J, Echassoux A (1988) L’industrie du Plèistocène inférieur de la grotte du Vallonet, Roquebrune-Cap-Martin, Alpes-Maritimes. L’anthropologie 92:501–614
Delaney-Rivera C, Plummer TW, Hodgson JA, Forrest F, Hertel F, Oliver JS (2009) Pits and pitfalls: taxonomic variability and patterning in tooth mark dimensions. J Archaeol Sci 36(11):2597–2608. https://doi.org/10.1016/J.JAS.2009.08.001
Delfino M, Atzori M (2013) An update on the Early Pleistocene herpetofauna from Pirro Nord. Palaeontographica Abt. A: Palaeozoology - Stratigraphy 298(1–6):19–29
Delfino M, Bailon S (2000) Early pleistocene herpetofauna from Cava Dell’Erba and Cava Pirro (Apulia, Southern Italy). Herpetol J 10(3)
Dennell RW (2008a) The taphonomic record of Upper Siwalik (Pinjor stage) landscapes in the Pabbi Hills, northern Pakistan, with consideration regarding the preservation of hominin remains. Quat Int 192(1):62–77. https://doi.org/10.1016/J.QUAINT.2007.06.024
Dennell RW (2008b) June. Human Migration and Occupation of Eurasia, Episodes
Dennell RW (2015) The Jaramillo Subchron, a geochronological marker horizon in the palaeoanthropological record of China. Quat Int 389:241–254. https://doi.org/10.1016/j.quaint.2014.11.006
Despriée J, Gageonnet R, Voinchet P, Bahain JJ, Falguères C, Varache F, Courcimault G, Dolo JM (2006) Une occupation humaine au Pléistocène inférieur sur la bordure nord du Massif Central. Comptes Rendu PALEVOL 5:821–828
Despriée J, Voinchet P, Tissoux H, Arzarello M, Robin S, Bahain J-J et al (2010) Lower and middle Pleistocene human settlements in the Middle Loire River Basin, Centre Region, France. Quat Int 223:345–359. https://doi.org/10.1016/j.quaint.2009.07.019
Domı́nguez-Rodrigo M (1997) Meat-eating by early hominids at the FLK 22Zinjanthropussite, Olduvai Gorge (Tanzania): an experimental approach using cut-mark data. J Hum Evol 33(6):669–690. https://doi.org/10.1006/JHEV.1997.0161
Domínguez-Rodrigo M, Barba R (2006) New estimates of tooth mark and percussion mark frequencies at the FLK Zinj site: the carnivore-hominid-carnivore hypothesis falsified. J Hum Evol 50(2):170–194. https://doi.org/10.1016/J.JHEVOL.2005.09.005
Domínguez-Rodrigo M, Piqueras A (2003) The use of tooth pits to identify carnivore taxa in tooth-marked archaeofaunas and their relevance to reconstruct hominid carcass processing behaviours. J Archaeol Sci 30(11):1385–1391. https://doi.org/10.1016/S0305-4403(03)00027-X
Domínguez-Rodrigo M, Yravedra J (2009) Why are cut mark frequencies in archaeofaunal assemblages so variable? A multivariate analysis. J Archaeol Sci 36(3):884–894. https://doi.org/10.1016/j.jas.2008.11.007
Domínguez-Rodrigo M, de Juana S, Galán, a. B., & Rodríguez, M. (2009) A new protocol to differentiate trampling marks from butchery cut marks. J Archaeol Sci 36:2643–2654. https://doi.org/10.1016/j.jas.2009.07.017
Domínguez-Rodrigo M, Pickering TR, Bunn HT (2012) Experimental study of cut marks made with rocks unmodified by human flaking and its bearing on claims of ∼3.4-million-year-old butchery evidence from Dikika, Ethiopia. J Archaeol Sci 39(2):205–214. https://doi.org/10.1016/j.jas.2011.03.010
Domínguez-Rodrigo M, Baquedano E, Mabulla A, Mercader J (2017) Paleoecological reconstructions of the Bed I and Bed II lacustrine basins of Olduvai Gorge (Tanzania) and insights into early human behavior. Palaeogeogr Palaeoclimatol Palaeoecol 488:1–8. https://doi.org/10.1016/J.PALAEO.2017.05.009
Echassoux A (2009) Les premiers Européens et la dynamique des interactions avec leur environnement : comportement écologique et niveau de cognition. L’Anthropologie 113(1):191–197. https://doi.org/10.1016/j.anthro.2008.12.003
Egeland, C. P. (2007). Zooarchaeological and taphonomic perspectives on hominid and carnivore interactions at Olduvai Gorge, Tanzania. Anthropology, PhD(June), 313
Egeland CP (2012) The use of bone surface modifications to model hominid lifeways during the Oldowan. In: Dominguez-Rodrigo M (ed) Stone tools and fossil bones : debates in the archaeology of human origins. Cambridge University Press, Cambridge, pp 80–114
Egeland CP, Pickering TR, Domínguez-Rodrigo M, Brain CK (2004) Disentangling Early Stone Age palimpsests: determining the functional independence of hominid- and carnivore-derived portions of archaeofaunas. J Hum Evol 47(5):343–357. https://doi.org/10.1016/j.jhevol.2004.08.004
Egeland CP, Gasparian B, Arakelyan D, Nicholson CM, Petrosyan A, Ghukasyan R, Byerly R (2014) Reconnaissance survey for Palaeolithic sites in the Debed River Valley, northern Armenia. J Field Archaeol 39(4):370–386. https://doi.org/10.1179/0093469014Z.00000000096
Eisenmann V (1979) Les Mètapodes d’equus sensu lato (Mammalia, Pèrissodactyla). Geobios 12(6):863–886
Ervynck A (1997) Detailed recording of tooth wear (Grant, 1982) as an evaluation of the seasonal slaughtering of pigs? Examples from Medieval sites in Belgium. Archaeofauna 6:67–79
Falguères C, Sémah F, Saleki H, Hameau S, Tu H, Féraud G, Simanjuntak H, Widianto H (2016) Geochronology of early human settlements in Java: what is at stake? Quat Int 416:5–11. https://doi.org/10.1016/j.quaint.2015.10.076
Fernández-jalvo Y, Andrews P (2003) Experimental effects of water abrasion on bone fragments. Journal of Taphonomy 1(3):147–164
Fernández-Jalvo, Y., & Andrews, P. (2016). Atlas of taphonomic identifications. Springer Netherlands
Forestier H (1993) Le Clactonien: Mise en application d’une nouvelle méthode de débitage s’inscrivant dans la variabilité des systèmes de production lithique du Paléolithique ancien. Paleo 5:53–82
Freudenthal M (1971) Neogene vertebrates from the Gargano Peninsula, Italy. Scr Geol 3:1–10
Gabunia LK (2000) Earliest Pleistocene hominid cranial remains from Dmanisi, Republic of Georgia: taxonomy, geological setting, and age. Science 288:1019–1025
Galán AB, Domínguez-Rodrigo M (2013) An experimental study of the anatomical distribution of cut marks created by filleting and disarticulation on long bone ends. Archaeometry 55(6):1132–1149. https://doi.org/10.1111/j.1475-4754.2012.00730.x
Garcia T, Féraud G, Falguères C, de Lumley H, Perrenoud C, Lordkipanidze D (2010) Earliest human remains in Eurasia: new 40Ar/39Ar dating of the Dmanisi hominid-bearing levels, Georgia. Quat Geochronol 5(4):443–451. https://doi.org/10.1016/j.quageo.2009.09.012
Gaudzinski S (1993) Early hominid subsistence in the Levant. Taphonomical studies at the Plio/Pleistocene ‘Ubeidiya Formation (Israel): evidence from layer II-24. In: Goren-Inbar N, Speth JD (eds) Human Paleoecology in the Levantine Corridor. (Oxbow Book., pp. 75–87), Oxford
Gaudzinski S (2004) Subsistence patterns of Early Pleistocene hominids in the Levant - taphonomic evidence from the ‘Ubeidiya Formation (Israel). J Archaeol Sci 31(8):65–75. https://doi.org/10.1016/S0305-4403(03)00100-6
Giusti D, Arzarello M (2016) The need for a taphonomic perspective in spatial analysis: formation processes at the Early Pleistocene site of Pirro Nord (P13), Apricena, Italy. JASREP 8:235–249. https://doi.org/10.1016/j.jasrep.2016.06.014
Gliozzi, E., Abbazzi, L., Argenti, P., Azzaroli, A., Caloi, L., Capasso Barbato, L., et al. (1997). Biochronology of selected mammals, molluscs and ostracods from the middle pliocene to the late pleistocene in Italy. The state of the art. Rivista Italiana di Paleontologia e Stratigrafia, 103(3), 369–388. doi:https://doi.org/10.13130/2039-4942/5299
Grant A (1982) The use of tooth wear as a guide to the age of domestic animals. In: Wilson B, Grigson C, Sebastian P (eds) Ageing and sexing animal bones from archaeological sites. BAR International Series. Archaeopress, Oxford, pp 91–108
Han F, Bahain J-J, Deng C, Boëda É, Hou Y, Wei G, Huang W, Garcia T, Shao Q, He C, Falguères C, Voinchet P, Yin G (2017a) The earliest evidence of hominid settlement in China: combined electron spin resonance and uranium series (ESR/U-series) dating of mammalian fossil teeth from Longgupo cave. Quat Int 434:75–83. https://doi.org/10.1016/j.quaint.2015.02.025
Han F, Bahain J-J, Deng C, Boëda É, Hou Y, Wei G et al (2017b) The earliest evidence of hominid settlement in China: combined electron spin resonance and uranium series (ESR/U-series) dating of mammalian fossil teeth from Longgupo cave. Quaternary International, 434. https://doi.org/10.1016/j.quaint.2015.02.025
Hardy BL (2004) Neanderthal behaviour and stone tool function at the Middle Palaeolithic site of La Quina, France. Antiquity 78(301):547–565. https://doi.org/10.1017/S0003598X00113213
Huguet R, Vallverdú J, Rodríguez-Álvarez XP, Terradillos-Bernal M, Bargalló A, Lombera-Hermida A, Menéndez L, Modesto-Mata M, van der Made J, Soto M, Blain HA, García N, Cuenca-Bescós G, Gómez-Merino G, Pérez-Martínez R, Expósito I, Allué E, Rofes J, Burjachs F, Canals A, Bennàsar M, Nuñez-Lahuerta C, Bermúdez de Castro JM, Carbonell E (2017) Level TE9c of Sima del Elefante (Sierra de Atapuerca, Spain): a comprehensive approach. Quat Int 433:278–295. https://doi.org/10.1016/j.quaint.2015.11.030
Hussain S (1975) Evolutionary and functional anatomy of the pelvic limb in fossil and recent Equidae (Perissodactyla, Mammalia). Anatomia, Histologia, Embryologia: Journal of Veterinary Medicine Series C 4(3):193–222. https://doi.org/10.1111/j.1439-0264.1975.tb00637.x
Iwase A (2016) A functional analysis of the LGM microblade assemblage in Hokkaido, northern Japan: a case study of Kashiwadai 1. Quat Int 425:140–157. https://doi.org/10.1016/j.quaint.2016.04.008
Karampatsou, T. (2017). New data on stratigraphy and formation processes from Pirro 13 site of Pirro Nord ( Apulia , Italy ). Università degli Studi di Ferrara
Keeley LH (1980) Experimental determination of stone tool uses: a microwear analysis. The University of Chicago Press, Chicago
Keeley LH, Toth N (1981) Microwear polishes on early stone tools from Koobi fora, Kenya. Nature 293(5832):464–465. https://doi.org/10.1038/293464a0
Lemorini C, Stiner MC, Gopher A, Shimelmitz R, Barkai R (2006) Use-wear analysis of an Amudian laminar assemblage from the Acheuleo-Yabrudian of Qesem Cave, Israel. J Archaeol Sci 33(7):921–934. https://doi.org/10.1016/j.jas.2005.10.019
Lemorini C, Plummer TW, Braun DR, Crittenden AN, Ditchfield PW, Bishop LC, Hertel F, Oliver JS, Marlowe FW, Schoeninger MJ, Potts R (2014a) Old stones’ song: use-wear experiments and analysis of the Oldowan quartz and quartzite assemblage from Kanjera South (Kenya). J Hum Evol 72:10–25. https://doi.org/10.1016/j.jhevol.2014.03.002
Lemorini C, Venditti F, Assaf E, Parush Y, Barkai R, Gopher A (2014b) The function of recycled lithic items at late lower Paleolithic Qesem cave, Israel: an overview of the use-wear data. Quat Int 361:103–112. https://doi.org/10.1016/j.quaint.2014.07.032
Lemorini C, Bourguignon L, Zupancich A (2016) A scraper’s life history: Morpho-techno-functional and use-wear analysis of Quina and demi-Quina scrapers from Qesem cave, Israel. Quat Int 398:86–93. https://doi.org/10.1016/j.quaint.2015.05.013
Levi Sala I (1986) Use wear and post-depositional surface modification: a word of caution. J Archaeol Sci 13(3):229–244. https://doi.org/10.1016/0305-4403(86)90061-0
Longo L (1994) Le industrie litiche. L’analisi delle tracce d’uso. In: Peretto C (ed) Le industrie litiche del giacimento paleolitico di Isernia la Pineta. La tipologia, le tracce di utilizzazione, la sperimentazione. Isernia, Cosmo Iannone, pp 355–404
Lopez-Garcìa JM, Luzi E, Berto C, Peretto C, Arzarello M (2015) Chronological context of the first hominin occurrence in southern Europe: the Allophaiomys ruffoi (Arvicolinae, Rodentia, Mammalia) from Pirro 13 (Pirro Nord, Apulia, southwestern Italy). Quat Sci Rev 107:260–266. https://doi.org/10.1016/j.quascirev.2014.10.029
López-González F, Grandal-d’Anglade A, Vidal-Romaní JR (2006) Deciphering bone depositional sequences in caves through the study of manganese coatings. J Archaeol Sci 33(5):707–717. https://doi.org/10.1016/j.jas.2005.10.006
Lyman RL (1994) Vertebrate taphonomy. Cambridge University Press, Cambridge
Malassé Dambricourt A, Moigne A-M, Singh M, Calligaro T, Karir B, Gaillard C et al (2016) Intentional cut marks on bovid from the Quranwala zone, 2.6 ma, Siwalik frontal range, northwestern India. Comptes Rendus Palevol 15(3–4):317–339. https://doi.org/10.1016/J.CRPV.2015.09.019
Marcolini F, Masini F, Argenti P (2013) The rodents of the Pirro Nord fauna (Foggia, Southern Italy). Palaeontographica Abt. A: Palaeozoology - Stratigraphy 298(1–6):73–85
Marín-Arroyo AB, Landete-Ruiz MD, Seva-Román R, Lewis MD (2014) Manganese coating of the Tabun faunal assemblage: implications for modern human behaviour in the Levantine Middle Palaeolithic. Quat Int 330(1):10–18. https://doi.org/10.1016/j.quaint.2013.07.016
Masini F, Sala B (2007) Large- and small-mammal distribution patterns and chronostratigraphic boundaries from the Late Pliocene to the Middle Pleistocene of the Italian peninsula. Quat Int 160(1):43–56. https://doi.org/10.1016/j.quaint.2006.09.008
Masini F, Sala B (2011) Considerations on an integrated biochronological scale of Italian Quaternary continental mammals. Il Quatrnario 24(2):193–198
Merritt SR (2012) Factors affecting early stone age cut mark cross-sectional size: implications from actualistic butchery trials. J Archaeol Sci 39(9):2984–2994. https://doi.org/10.1016/j.jas.2012.04.036
Mgeladze A, Lordkipanidze D, Moncel M-H, Despriée J, Chagelishvili R, Nioradze M (2010) First human settlement of the Caucasus: technical behavior and raw material acquisition at Dmanisi, Georgia (1.77 Ma). Quat Int 223–224:422–425
Moss, E. H. (1983). The Functional analysis of flint implements Pincevent and Pont d’Ambon: two case studies from the French final Palaeolithic. Oxford: British Archaeological Reports International Series 177. Archaeopress
Moyano IT, Barsky D, Cauche D, Celiberti V, Grégoire S, Lebegue F, Moncel MH, de Lumley H (2011) The archaic stone tool industry from Barranco León and Fuente Nueva 3, (Orce, Spain): evidence of the earliest hominin presence in southern Europe. Quat Int 243(1):80–91. https://doi.org/10.1016/j.quaint.2010.12.011
Muttoni G, Scardia G, Kent DV, Morsiani E, Tremolada F, Cremaschi M, Peretto C (2011) First dated human occupation of Italy at {~}0.85Ma during the late Early Pleistocene climate transition. Earth Planet Sci Lett 307(3–4):241–252. https://doi.org/10.1016/j.epsl.2011.05.025
Nilssen PJ (2000) An Actualistic butchery study in South Africa and its implications for reconstructing hominid strategies of carcass acquisition and butchery in the Upper Pleistocene and Plio-Pleistocene. University of Cape Town
Odell GH (1981) The mechanism of use-breakage of stone tools: some testable hypothesis. J Field Archaeol 8:197–209
Odell GH, Cowan F (1986) Experiments with spears and arrows on animal targets. J Field Archaeol 13(2):195–212. https://doi.org/10.1179/009346986791535780
Parés JM, Pérez-González A, Rosas A, Benito A, Bermúdez de Castro JM, Carbonell E, Huguet R (2006) Matuyama-age lithic tools from the Sima del Elefante site, Atapuerca (northern Spain). J Hum Evol 50(2):163–169. https://doi.org/10.1016/j.jhevol.2005.08.011
Pavia M, Zunino M, Coltorti M, Angelone C, Arzarello M, Bagnus C, Bellucci L, Colombero S, Marcolini F, Peretto C, Petronio C, Petrucci M, Pieruccini P, Sardella R, Tema E, Villier B, Pavia G (2012) Stratigraphical and palaeontological data from the Early Pleistocene Pirro 10 site of Pirro Nord (Puglia, south eastern Italy). Quat Int 267:40–55. https://doi.org/10.1016/j.quaint.2010.12.019
Pawlik AF (2013) Detecting traits of modern behavior through microwear analysis. Asian Perspect 53:1689–1699. https://doi.org/10.1017/CBO9781107415324.004
Peretto C, Amore FO, Antoniazzi A, Aantoniazzi A, Bahain J-J, Cattani L et al (1998a) L’industrie lithique de Ca’Belvedere di Monte Poggiolo : Stratigraphie, matière première, typologie, remontages et traces d’utilisation. L’ Anthropologie 102(4):343–465
Peretto P, Despriée J, Gageonnet R, Voinchet P, Bahain J-J, Falguères C et al (1998b) Technological change and population growth. J Econ Growth 3(4):283–311. https://doi.org/10.1016/j.crpv.2006.03.001
Petronio C, Marcolini F (2013) Mammal Biocrhronology at the end of Late Villafranchian (Early Pleistocene): Pirro Faunal Unit. Palaeontographica Abt. A: Palaeozoology - Stratigraphy 298(1–6):183–191
Petrucci M, Cipullo A, Martínez-Navarro B, Rook L, Sardella R (2013) The Late Villafranchian (Early Pleistocene) carnivores (Carnivora, Mammalia) from Pirro Nord (Italy). Palaeontogr Abt A 298(1–6):113–145. https://doi.org/10.1127/pala/298/2013/113
Pickering TR, Egeland CP (2006) Experimental patterns of hammerstone percussion damage on bones: implications for inferences of carcass processing by humans. J Archaeol Sci 33(4):459–469. https://doi.org/10.1016/j.jas.2005.09.001
Pobiner BL (2015) New actualistic data on the ecology and energetics of hominin scavenging opportunities. J Hum Evol 80:1–16. https://doi.org/10.1016/J.JHEVOL.2014.06.020
Pobiner BL, Rogers MJ, Monahan CM, Harris JWK (2008) New evidence for hominin carcass processing strategies at 1.5 Ma, Koobi Fora, Kenya. J Hum Evol 55(1):103–130. https://doi.org/10.1016/J.JHEVOL.2008.02.001
Rodríguez-Gómez G, Palmqvist P, Rodríguez J, Mateos A, Martín-González JA, Espigares MP, Ros-Montoya S, Martínez-Navarro B (2016) On the ecological context of the earliest human settlements in Europe: resource availability and competition intensity in the carnivore guild of Barranco León-D and Fuente Nueva-3 (Orce, Baza Basin, SE Spain). Quat Sci Rev 143:69–83. https://doi.org/10.1016/J.QUASCIREV.2016.05.018
Rook L, Sardella R (2013) New data on the Early Pleistocene large sized porcupine from Pirro Nord (Apricena, Apulia, Italy). Palaeontogr Abt A 298(1–6):87–94. https://doi.org/10.1127/pala/298/2013/87
Rots V (2010) Prehension and hafting traces on flint tools: a methodology. Leuven University Press, Leuven
Sahnouni M, Rosell J, van der Made J, Vergès JM, Ollé A, Kandi N, Harichane Z, Derradji A, Medig M (2013) The first evidence of cut marks and usewear traces from the Plio-Pleistocene locality of El-Kherba (Ain Hanech), Algeria: implications for early hominin subsistence activities circa 1.8 Ma. J Hum Evol 64(2):137–150. https://doi.org/10.1016/j.jhevol.2012.10.007
Saladié P, Rodríguez-Hidalgo A, Huguet R, Cáceres I, Díez C, Vallverdú J, Canals A, Soto M, Santander B, Bermúdez de Castro JM, Arsuaga JL, Carbonell E (2014) The role of carnivores and their relationship to hominin settlements in the TD6-2 level from Gran Dolina (Sierra de Atapuerca, Spain). Quat Sci Rev 93:47–66. https://doi.org/10.1016/J.QUASCIREV.2014.04.001
Salari L, Kotsakis T, Petronio C (2013) Early Pleistocene bats from Pirro Nord (Apulia, Southern Italy). Palaeontogr Abt A 298(1–6):55–72. https://doi.org/10.1127/pala/298/2013/55
Selvaggio MM (1998) Evidence for a three-stage sequence of hominid and carnivore involvement with long bones at FLKZinjanthropus, Olduvai Gorge, Tanzania. J Archaeol Sci 25:191–202
Semenov SA (1964) Prehistoric technology; an experimental study of the oldest tools and artefacts from traces of manufacture and wear. Cory. Adams and Mackay, London
Shen C, Chen C (2000) A use-wear study of lithic artifacts from Xiaochangliang and hominid activities in Nihewan Basin. Acta Anthropol Sinica 19:119–125
Shipman P, Rose J (1983) Early hominid hunting, butchering, and carcass-processing behaviors: approaches to the fossil record. J Anthropol Archaeol 2:57–98. https://doi.org/10.1016/0278-4165(83)90008-9
Solodenko N, Zupancich A, Cesaro SN, Marder O, Lemorini C, Barkai R (2015) Fat residue and use-Wear found on Acheulian biface and scraper associated with butchered elephant remains at the site of Revadim, Israel. PLoS One 10(3):e0118572. https://doi.org/10.1371/journal.pone.0118572
Soulier M-C, Costamagno S (2017) Let the cutmarks speak! Experimental butchery to reconstruct carcass processing. J Archaeol Sci Rep 11:782–802. https://doi.org/10.1016/J.JASREP.2016.12.033
Stiner MC (1998) Mortality analysis of Pleistocene bears and its paleoanthropological relevance. J Hum Evol 34(3):303–326. https://doi.org/10.1006/jhev.1997.0198
Swisher CC, Curtis GH, Jacob T, Getty AG, Suprijo A, Widiasmoro (1994) Age of the earliest known hominids in Java, Indonesia. Science 263(5150):1118–1121
Terradillos-Bernal M, Rodríguez-Álvarez XP (2017) The influence of raw material quality on the characteristics of the lithic tool edges from the Atapuerca sites (Burgos, Spain). Quat Int 433:211–223. https://doi.org/10.1016/j.quaint.2015.10.122
Toro-Moyano I, Martínez-Navarro B, Agustí J, Souday C, Bermúdez de Castro JM, Martinón-Torres M, Fajardo B, Duval M, Falguères C, Oms O, Parés JM, Anadón P, Julià R, García-Aguilar JM, Moigne AM, Espigares MP, Ros-Montoya S, Palmqvist P (2013) The oldest human fossil in Europe, from Orce (Spain). J Hum Evol 65(1):1–9. https://doi.org/10.1016/j.jhevol.2013.01.012
Tringham R, Cooper G, Odell GH, Voytek B, Whitman A (1974) Experimentation in the formation of edge damage: a new approach to lithic analysis. J Field Archaeol 1:171–196
Val A, Costamagno S, Discamps E, Chong S, Claud E, Deschamps M, Mourre V, Soulier MC, Thiébaut C (2017) Testing the influence of stone tool type on microscopic morphology of cut-marks: experimental approach and application to the archaeological record with a case study from the Middle Palaeolithic site of Noisetier Cave (Fréchet-Aure, Hautes-Pyrénées, France). J Archaeol Sci Rep 11:17–28. https://doi.org/10.1016/J.JASREP.2016.11.028
Van Gijn AL (2014) Science and interpretation in microwear studies. J Archaeol Sci 48:166–169. https://doi.org/10.1016/j.jas.2013.10.024
Vekua AK (2002) A new skull of early Homo from Dmanisi, Georgia. Science 297:85–89
Villa P, Mahieu E (1991) Breakage patterns of human long bones. J Hum Evol 21(1):27–48. https://doi.org/10.1016/0047-2484(91)90034-S
Visentin D, Bertola S, Ziggiotti S, Peresani M (2015) Going off the beaten path? The Casera Lissandri 17 site and the role of the Cansiglio plateau on human ecology during the Early Sauveterrian in North-eastern Italy. Quat Int 423:213–229. https://doi.org/10.1016/j.quaint.2015.11.119
Wilkins J, Schoville BJ, Brown KS, Chazan M (2015) Kathu Pan 1 points and the assemblage-scale, probabilistic approach: a response to Rots and Plisson, “Projectiles and the abuse of the use-wear method in a search for impact.”. J Archaeol Sci 54:294–299. https://doi.org/10.1016/j.jas.2014.12.003
Yeshurun R, Zaidner Y, Eisenmann V, Martínez-Navarro Bienvenido B, Bar-Oz G (2011) Lower Paleolithic hominin ecology at the fringe of the desert: faunal remains from Bizat Ruhama and Nahal Hesi, Northern Negev, Israel. J Hum Evol 60(4):492–507. https://doi.org/10.1016/j.jhevol.2010.01.008
Zhu RX, Potts R, Pan YX, Lü LQ, Yao HT, Deng CL, Qin HF (2008) Paleomagnetism of the Yuanmou Basin near the southeastern margin of the Tibetan plateau and its constraints on late Neogene sedimentation and tectonic rotation. Earth Planet Sci Lett 272(1–2):97–104. https://doi.org/10.1016/j.epsl.2008.04.016
Ziggioti S (2005) Tecnologia e Funzione. Apporto dell’analisi funzionale delle industrie litiche dell’Epigravettiano recente nelle Prealpi orientali italiane. Consorzio universitario di Bologna, Ferrara e Parma
Acknowledgements
We thank the Gervasio family, owner of the excavation area, for its kindness and availability. We thank also Gaetano and Franco dell’Erba for their constant support for our research and the Municipality of Apricena for its economic help and general support. The work in the Pirro Nord site was possible thanks to collaboration with the Soprintendenza Archeologia, Belle Arti e Paesaggio per le Province di Barletta-Andria-Trani e Foggia. We thank M.L. Caparrini and Fabio Bertone for the English revision.
Author information
Authors and Affiliations
Contributions
Analyzed the data: RCC, IC, RH, GLFB, CB, MA. Wrote the paper: GLFB, CB, RCC, MA. Analysis of faunal remains: FC, CB, BS, GMC. Analysis of lithic tools: GLFB, MA, SD. Geomorphological analyses: SC, TK, MZ. Coordination of the excavation: MA, JA, GLFB, SD, CB, GMC, SC.
Corresponding author
Additional information
Communicated by: Mary T. Silcox
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
The oldest European traces of Homo-mamma’ interactions: new data from Pirro Nord 13 (Apricena, Italy)
Rights and permissions
About this article

Cite this article
Cheheb, R.C., Arzarello, M., Arnaud, J. et al. Human behavior and Homo-mammal interactions at the first European peopling: new evidence from the Pirro Nord site (Apricena, Southern Italy). Sci Nat 106, 16 (2019). https://doi.org/10.1007/s00114-019-1610-4
Received:
Revised:
Accepted:
Published:
DOI: https://doi.org/10.1007/s00114-019-1610-4