
Abstract
The fossil record of Chamaeleonidae is very scarce and any new specimen is therefore considered important for our understanding of the evolutionary and biogeographic history of the group. New specimens from the early Miocene of Aliveri (Evia Island), Greece constitute the only fossils of these lizards from southeastern Europe. Skull roofing material is tentatively attributed to the Czech species Chamaeleo cf. andrusovi, revealing a range extension for this taxon, whereas tooth-bearing elements are described as indeterminate chamaeleonids. The Aliveri fossils rank well among the oldest known reptiles from Greece, provide evidence for the dispersal routes of chameleons out of Africa towards the European continent and, additionally, imply strong affinities with coeval chamaeleonids from Central Europe.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Chameleons (Squamata, Chamaeleonidae) constitute a rather diverse group with more than 200 species that are now distributed in Africa, Madagascar and several other Indian Ocean islands, southern Asia, Cyprus and southern parts of Mediterranean Europe (Glaw 2015). Cryptic diversity appears to be prominent within the group, with several new species having been described only during the current decade, mostly on the basis of molecular data (Gehring et al. 2011; Crottini et al. 2012; Greenbaum et al. 2012; Stipala et al. 2011, 2012; Tilbury and Tolley 2015). The size range of chamaeleonids is also broad and astonishing, with the larger members of the family surpassing 600 mm in total length (Glaw and Vences 1994), whereas the smallest species rank well among the tiniest known reptiles, both extant (Brookesia micra, attaining only 29 mm [Glaw et al. 2012]) and extinct (e.g. Jucaraseps grandipes [Bolet and Evans 2012]). The ability of some species to ‘change’ colour, along with their peculiar external morphology, have gained the attention of the public for centuries and render them readily distinguishable even for non-specialists (Teyssier et al. 2015). Moreover, their bizarre external appearance, acrodont dentition, unique skeletal anatomy, as even their specialized feeding and locomotor habits have long lead herpetologists to consider chameleons as a monophyletic group (Camp 1923; Estes 1983). Their relationships to other squamates, however, are yet not fully resolved, mostly because of the uncertainty of the position of Iguania, but it is widely accepted that Chamaeleonidae constitute a group within the iguanian clade Acrodonta (Estes et al. 1988; Townsend et al. 2004; Kumazawa 2007; Conrad 2008; Vidal and Hedges 2009; Gauthier et al. 2012; Wiens et al. 2012; Pyron et al. 2013; Reeder et al. 2015), even if their affinities with Agamidae are a matter of debate, with different topologies arising on the basis of morphological and molecular data (Bolet and Evans 2013; Tolley and Menegon 2013).
Despite the broad geographic distribution among extant taxa, the fossil record of Chamaeleonidae is rather patchy (Bolet and Evans 2013). A Cretaceous origin of the group is commonly accepted based on molecular data (Tolley et al. 2013), but with their first fossil record dating back to the early Miocene (Moody and Roček 1980; Pickford 1986), such molecular divergence date cannot be thoroughly tested. The dispersal of chamaeleonids to Europe and Asia seems to have occurred later (Bolet and Evans 2013; Tolley et al. 2013); indeed, their Neogene record on the European continent is rather scanty with few, scattered occurrences only across the early to middle Miocene of Central Europe (Moody and Roček 1980; Böhme and Ilg 2003; Čerňanský 2010; Bolet and Evans 2013).
Europe currently harbours two extant taxa of chamaeleonids: Chamaeleo chamaeleon (Linnaeus, 1758) and Chamaeleo africanus Laurenti, 1768 (Dimaki et al. 2000, 2008, 2015). The Greek distribution of the former species now solely comprises Samos island, as populations inhabiting Crete and Chios seem to have gone extinct recently. It is worth noting, however, that Sillero et al. (2014) reported this species as still present in Crete, considering it allochthonous. The distribution of C. africanus in Greece (restricted only to a small region near the town of Pylos, Peloponnese) is the sole European population for this African species and most probably is a product of human introduction during early antiquity (Fig. 2) (Böhme et al. 1998; Kosuch et al. 1999; Dimaki et al. 2000, 2008).
Despite their occurrence in the extant herpetofauna of the country, chamaeleonids have never been previously described from Greek fossil localities. Here, we report on three new chamaeleonid fossils found in the early Miocene (MN 4) locality of Aliveri, Evia Island, which indicate the Neogene existence of these lizards in Greece. One of these specimens is tentatively attributed to the species Chamaeleo cf. andrusovi Čerňanský, 2010, which was previously known only from the early Miocene of the Czech Republic, whereas the other two specimens represent indeterminate chamaeleonids.
Geological setting
Aliveri is a fossiliferous locality, located within the Neogene sedimentary basin of Aliveri/Kymi, in Evia (also spelled Euboea) Island in Central Greece. Along with the Gavathas site, Lesvos Island, it has yielded the oldest known Greek Neogene land mammal fauna (Koufos 2006a, b; van den Hoek Ostende et al. 2015). Aliveri was discovered in 1977 and was initially dated to the MN 3 zone (de Bruijn et al. 1980). Nevertheless, there is a general consensus now that it is referable to MN 4 (Doukas 2003; Koufos 2006b; van den Hoek Ostende et al. 2015). More specifically, as was recently stated by van den Hoek Ostende et al. (2015:276), ‘although undeniably an MN 4 assemblage, Aliveri is one of the oldest localities from that MN unit, and presumably co-eval with many of the MN 3 assemblages from western and central Europe’. The age of Aliveri is tentatively estimated between 18 and 17.5 Ma, and corresponds to the Burdigalian stage (Orleanian Continental Stage) (Koufos 2006b; van den Hoek Ostende et al. 2015). A detailed description of the geology of Aliveri is provided by de Bruijn et al. (1980).
The micromammal assemblage of Aliveri is rather diverse and has been the subject of several studies (de Bruijn et al. 1980; van der Meulen and de Bruijn 1982; Schmidt-Kittler 1983; Klein Hofmeijer and de Bruijn 1985, 1988; Doukas 1986; López Martinez 1986; Álvarez Sierra et al. 1987). It includes erinaceids, dimylids, heterosoricids, talpids, soricids, ochotonids, eomyids, many sciurids, cricetids, anomalomyids, spalacids and glirids. Larger mammals are less known, but palaeogalids, equids, palaeomerycids, cervids and bovids are present, as well as the viverrid Euboictis (van den Hoek Ostende et al. 2015). As for the herpetofauna of Aliveri, anurans, crocodylians, anguids, lacertids, as well as indeterminate lizards, snakes and turtles are present but have not been described. The study of these amphibians and reptiles will be the subject of a forthcoming publication.
Material and methods
All three chamaeleonid specimens described herein belong to the collections of the Department of Earth Sciences in the University of Utrecht and are accessioned under the repository numbers UU AL 3501–3503. All specimens were found in the early Miocene locality of Aliveri in three different field seasons, in 1977 (UU AL 3502), 1978 (UU AL 3501), and 1980 (UU AL 3503), led by the University of Utrecht. As with the micromammal material that has also been collected from this locality, all chamaeleonid fossils have been recovered from the underclay of the lignite occurring North of Aliveri, along the eastern edge of the basin where the lignite wedges out against Mesozoic limestones (de Bruijn et al. 1980). For comparison purposes, skeletal material of extant chamaeleonids was studied in the herpetological collections of the University of Torino and the Muséum national d’Histoire naturelle, Paris. Photos of the fossil specimens were taken using the Leica M205 microscope and the Leica application suite V 3.3.0, in the University of Torino.
Results
Systematic palaeontology
Reptilia Laurenti 1768
Squamata Oppel, 1811
Iguania Cope, 1864
Acrodonta Cope, 1864
Chamaeleonidae Gray, 1825
Chamaeleo Laurenti, 1768
Chamaeleo cf. andrusovi Čerňanský, 2010
(Fig. 1a, b, c, d)

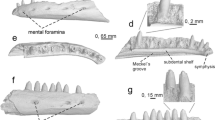
Fossil chamaeleonids from Aliveri. a–d Chamaeleo cf. andrusovi: skull roofing bone, UU AL 3501, in external (a) and internal (b) views, and its interpretative drawing in external (c) and lateral (d) views. e–h Chamaeleonidae indet.: tooth-bearing bones, UU AL 3502 (e–f) and UU AL 3503 (g–h), in lateral (e, g) and medial (f, h) views. Scale bars represent 1 mm
Material: The specimen here referred to Chamaeleo cf. andrusovi is a skull roofing bone fragment (UU AL 3501).
Description: UU AL 3501 is a skull roofing bone fragment about 3 × 3 mm in diameter. A comparison with extant chamaeleonid material, stored in the collections of the University of Torino and the Muséum national d’Histoire naturelle, Paris, suggests that the fragment can be part of the circumorbital region (i.e. either a prefrontal or a postorbital), but its fragmentary status prevents a more precise determination. The fragment is flattened and its external surface is covered by an ornamentation made by various small tubercles grouped in four, well-separated and rounded clusters. Two of the four clusters are aligned along one of the margins of the fragment, which is a natural border and not the result of breakage (as the other margins). If our interpretation of the fragment as part of a circumorbital bone is correct, this would be the orbital margin. The inner surface of the specimen is smooth.
Chamaeleonidae indet.
(Fig. 1e–h)
Material: Two additional specimens from the same locality are assigned to Chamaeleonidae, but cannot be attributed to the species level. These two specimens are both tooth bearing bones (UU AL 3502 and UU AL 3503).
Description: UU AL 3502 and UU AL 3503 are very fragmentary tooth-bearing bones. However, they clearly bear acrodont, large and triangular teeth, which are well-spaced and located on the edge of the bones. Two of them are preserved in UU AL 3502 and three of them are visible in UU AL 3503, even if in the latter specimen only one is well preserved. Small accessory cusps are recognizable anteriorly and posteriorly to the main one in one tooth of UU AL 3502 and in two teeth of UU AL 3503. Ventrally directed interdental grooves are present on the lateral surface. Due to the fragmentary nature of both specimens, it is currently impossible to determine whether these two tooth-bearing elements originate from the dentary or the maxilla.
Taxonomic identification
The specimens described herein were preliminarily described in an unpublished report by Delfino (unpublished), who attributed them to Chamaeleonidae indet. On the other hand, Böhme and Ilg (2003) cited for the same locality the presence of Chamaeleo cf. C. caroliquarti in their online database, without further details or describing any material.
Because of the complex ornamentation and the well-spaced and complex clusters of tubercles (= pustular protuberances of Čerňanský (2010)), the skull bone fragment (UU AL 3501) from Aliveri can be referred to Chamaeleo cf. andrusovi. In the original diagnosis provided for this species, Čerňanský (2010:609) stated that C. andrusovi was a small chamaeleonid, differing from all other taxa in several features, but most notably by ‘its typical strongly pustular ornamentation of the external surfaces of the skull roofing bones. Differs from adult Chamaeleo calyptratus, Triceros (sic) hoehneli or Calumma globifer in regard to the concentration, distribution and shape of the protuberances—mostly complicated pustular protuberances are moderately spaced and rather evenly distributed, covering the otherwise smooth external surface’. This diagnostic feature appears also in the Greek specimen; therefore, the skull roofing bone is attributed to Chamaeleo cf. andrusovi.
The two tooth-bearing elements from Aliveri (UU AL 3502 and UU AL 3503) can be attributed to the Chamaeleonidae on the basis of the well-spaced and apically located acrodont teeth (Delfino et al. 2008 and references therein). The identification of chamaeleonid tooth-bearing bones is severely hampered by the widespread, homoplastic features surrounding their anatomy. Generally, the dentaries of Acrodonta share several common features that should prompt us being rather cautious when dealing with taxonomic assignments to either agamids or chamaeleonids (Augé 1997; Delfino et al. 2008; Rage and Bailon 2011). Even if the two fragmentary tooth-bearing bones cannot be identified as portions of maxillae or dentaries, it is worth noting that Čerňanský (2010) has shown that dentaries of chamaeleonids are not diagnostic to the species level. Assigning the tooth-bearing bones to Chamaeleo cf. andrusovi on just the base of the taxonomic allocation of the skull roofing bone seems premature for the moment, as fossil evidence has shown that distinct chameleon species could have co-existed in sympatry: different morphologies that have been observed among the dentaries of C. caroliquarti (Čerňanský 2010) could indicate the existence of more than one chameleon at Dolnice. The fact that C. caroliquarti was a very large species (around 500 mm in total length) (Bolet and Evans 2013), whereas C. andrusovi was originally described as ‘a small species of Chamaeleo’ (Čerňanský 2010, p. 609), prompts us to leave open the possibility that Dolnice could harbour at least two different chamaeleonid taxa. The idea of two or more sympatric chamaeleonid species in the same locality is reminiscent of a similar situation in several extant species (Raselimanana and Rakotomalala 2003; Tolley and Menegon 2013). Such higher species diversity within a single region has been also recently inferred for fossil chamaeleonids (Dollion et al. 2015). On the other hand, many regions harbour only monospecific communities of extant chamaeleonids (Tolley and Menegon 2013). Taking all these into consideration, whether Aliveri hosted a monospecific chameleon assemblage, or whether multiple sympatric species coexisted, remains only to be revealed on the light of new and more complete specimens from the locality. As such, we refrain from attributing the two tooth-bearing elements to the same species as the skull roofing bone (Chamaeleo cf. andrusovi) and we thus tentatively consider them as ‘Chamaeleonidae indet.’
Discussion
The taxonomy of European Neogene chamaeleonids
Six Miocene species have already been named on the basis of rare European fossil remains, which mostly do not comprise overlapping material (Bolet and Evans 2013): C. caroliquarti Moody and Roček, 1980, Chamaeleo bavaricus Schleich, 1983, Chamaeleo pfeili Schleich, 1984, Chamaeleo simplex Schleich, 1994, Chamaeleo sulcodentatus Schleich, 1994 and Chamaeleo andrusovi Čerňanský, 2010. Several anatomical characters appear to be ubiquitous within chamaeleonids, rendering the validity of most of these taxa as tentative or even dubious (Čerňanský 2011). This applies especially to characters in the dentaries of these squamates, which appear to be widespread and indistinguishable from extant representatives. Čerňanský (2010) revised the first named European fossil taxon C. caroliquarti and regarded it as a nomen dubium, as he could not differentiate the holotype dentary from several extant taxa, whereas the paratype dentary appeared to belong to a distinct morphotype. We acknowledge here, however, that on several occasions, even fragmentary fossils can bear diagnostic characters. This is the situation, for example, when geometric morphometric approaches are used (Bastir et al. 2014; Cornette et al. 2015), a case study that has also been recently applied to fossil chamaeleonids (Dollion et al. 2015). The latter authors characteristically noted that even small fragments such as parts of the maxilla and the parietal could be useful in taxonomic identification. It is, however, beyond the scope of this paper to evaluate the status of C. caroliquarti, and acknowledge that a complete phylogenetic analysis which will, in fact, include enough dentary characters from multiple extant taxa, is needed in order to decipher the systematics of European Neogene chameleons.
The Aliveri chameleons
The identification of the Aliveri specimens described herein, as chamaeleonids, confirms the presence of this squamate group in the fossil record of Greece. Another purported acrodont taxon, Uromastyx spinipes (Daudin, 1802) [= Uromastyx aegyptia (Forskål, 1775)] was reported by Mangili (1980) from the Late Pleistocene of Grotta Simonelli in Crete, on the basis of an isolated, 43 mm long, tibia that was not figured and not described in detail, hindering any evaluation of its identity. The location of this fossil is currently unknown. However, since the comparative morphology of lizard tibiae is poorly known, this specimen should be considered an indeterminate squamate. As such, the specimens from Aliveri comprise the sole record of fossil chameleons from Greece.
The taxonomic assignment of the Aliveri skull roofing element as Chamaeleo cf. andrusovi provides a significant range extension for that species. C. andrusovi was until now known only from the early Miocene (MN 4) of Dolnice, Czech Republic (Čerňanský 2010). This new record indicates that this species was widely distributed in the early Miocene of Central and Southern Europe. Such a broad distribution is congruent with that of several extant members of the genus Chamaeleo (e.g. C. chamaeleon and C. africanus are widespread species with populations being adapted to different environments and ecological settings [Dimaki et al. 2000, 2008]). However, the high degree of cryptic speciation in modern chamaeleonids has led to the ‘split’ of several widespread extant taxa, mostly on the base of molecular data, but with few or no morphological characters that could distinguish them (e.g. Andreone et al. 2001; Nečas et al. 2003, 2005, 2009; Gehring et al. 2010, 2011; Crottini et al. 2012; Greenbaum et al. 2012; Stipala et al. 2011, 2012; Tilbury and Tolley 2015). This fact is further problematic for the fossil record of chamaeleonids, as for their identification we have to rely only on morphological evidence, and no assumption about cryptic speciation within extinct species can be suggested.
It is worth noting that the faunal assemblage of Aliveri is chronologically older than other MN 4 faunas in Europe (van den Hoek Ostende et al. 2015) and that the Greek skull roofing bone assigned to Chamaeleo cf. andrusovi is therefore older than the conspecific specimens from the type locality of Dolnice, Czech Republic. As such, the Aliveri specimen ranks among the oldest named chamaeleonids of Europe, being only slightly younger than C. caroliquarti which has been recorded from the early Miocene (MN 3) localities of Merkur Nord (Czech Republic) and Wintershof West (Germany).
The locality of Aliveri is well known for its rich micromammal fauna. The overall diverse and abundant fauna of flying squirrels suggest a forest biotope (de Bruijn et al. 1980). A relatively humid environment for the locality has also been suggested (van den Hoek Ostende et al. 2015). The new identification of Chamaeleo cf. andrusovi in Aliveri further confirms the already proposed ecological settings of the locality, as the presence of chamaeleonids is well known to be indicative of warm climates (Moody and Roček 1980) and further suggests relatively dense vegetation (Maul et al. 2011), although several extant genera are known to occur in both closed and open habitats (Tolley et al. 2008; Dollion et al. 2015).
Palaeobiogeography of the Chamaeleonidae
The biogeographic history of chamaeleonids is hampered by their extremely poor fossil record. Most of the material recovered is fragmentary, thus not permitting identification beyond the family level. Notable exceptions, however, are known to occur, with few exceptional Miocene specimens from Kenya documenting the morphological conservativeness and a relative evolutionary stasis within the group throughout time (Hillenius 1978a; Rieppel et al. 1992). Stem representatives of Chamaeleonidae are not known or at least they have not yet been identified, rendering their early evolutionary history tantalizing and obscure (Bolet and Evans 2013; Simões et al. 2015). Molecular data support a Cretaceous dichotomy between chamaeleonids and other Acrodonta, although the exact dates vary among different researches (Raxworthy et al. 2002; Wiens et al. 2006; Townsend et al. 2011; Bolet and Evans 2013; Dollion et al. 2015). Africa and Madagascar have long been proposed as potential centres of origin for chamaeleonids (Hillenius 1959, 1978b; Blanc 1972; Klaver 1977; Tolley et al. 2013; Dollion et al. 2015). More specifically, Tolley et al. (2013) suggested, on the base of molecular data, that chamaeleonids originated in Africa around the Late Cretaceous, after the break-up of the supercontinent Gondwana, with two independent oceanic dispersals to Madagascar (their modern biodiversity hotspot) during the Palaeocene and the Oligocene. Frustratingly, however, no Cretaceous and Palaeogene fossils are known, and it is characteristic that the first definite known chamaeleonid fossils are documented only in the early Miocene of Africa and Europe (Bolet and Evans 2013; Dollion et al. 2015).
Several enigmatic Palaeogene taxa from Asia bear superficial resemblance with chamaeleonids; however, they are now thought to represent convergent stem acrodonts (Bolet and Evans 2013). It is worth noting here that the taxonomic status of Anqingosaurus brevicephalus Hou, 1976 (variously cited also under the incorrect spelling Anquingosaurus), an enigmatic species from the Palaeocene of China, is pivotal to our understanding of chamaeleonid evolution. Bolet and Evans (2013, p. 184) doubted the status of A. brevicephalus as a chamaeleonid, mentioning characteristically that ‘there is nothing to suggest that it is a chameleon’. That being said, there is currently no definite record of a chamaeleonid from the Palaeogene or even the Neogene of Asia. In fact, the only known Asian fossil chamaeleonids are recorded in the Middle Pleistocene of Israel and the Palaeolithic of Lebanon (Haas 1952; Hooijer 1961; Maul et al. 2011; Bolet and Evans 2013), but palaeogeographically Middle East was part of Gondwana and not Asia.
Neogene chameleons in Europe appear to be restricted, both geographically and chronologically, with only few published occurrences from the early to middle Miocene of Czech Republic, Germany and Switzerland, and perhaps also Poland and Spain (Fig. 2) (Moody and Roček 1980; Roček 1984; Schleich 1984; Bolliger 1992; Fejfar and Schleich 1994; Mörs 2002; Böhme and Ilg 2003; Böhme 2010; Čerňanský 2010, 2011; Bolet and Evans 2013). The earliest occurrence of a chamaeleonid in Europe is documented in the early Miocene (MN 3) of Merkur Nord in Czech Republic and Wintershof West in Germany (Čerňanský 2010; Bolet and Evans 2013). A still undescribed chamaeleonid from the early Miocene (MN 3/MN 4) of Agramon, Spain, cited in the online database of Böhme and Ilg (2003), needs further investigation about its exact taxonomic affinities; if it is indeed a chamaeleonid though, then it represents one of the earliest European records of the group. Interestingly, chameleons are neither recovered from the well-known MN 2 squamate fauna of Wiesbaden-Amöneburg (Germany) nor in any other MN 2 European locality, leading Čerňanský et al. (2015) to suggest that their absence in the earliest Miocene is genuine and they only dispersed into Europe shortly thereafter. The presence of chamaeleonids in Central Europe during the early Miocene coincides with the Miocene Climatic Optimum (Böhme 2003), which enabled several other squamate groups to achieve their northernmost distribution (e.g. varanids and cordylids; Bolet and Evans 2013; Delfino et al. 2013). One or more widespread and/or even distinct, sympatric species could be present in the early Miocene of Central Europe at the time of this climatic optimum, as high temperatures favour the diversification of squamates. In any case, the permanence of Miocene chamaeleonids in Europe did not last long as suggested by their last published occurrence in the MN 6 of Ornberg in the Molasse Basin, Switzerland (Bolliger 1992; Čerňanský 2011), whereas an even later occurrence for the group is listed in the online database of Böhme and Ilg (2003), where an indeterminate reptile that was initially described by Młynarski et al. (1982) from the middle Miocene (MN 7) of Poland is there re-identified as a chamaeleonid. Whatever their last appearance datum, they likely went extinct due to climatic deterioration in Europe and/or ecological competition with newly arrived squamates. Interestingly, a new land bridge has been suggested to exist between Afro-Arabia and Asia during MN 6, which, apparently, favoured a new wave of dispersal of immigrants to Europe (Koufos et al. 2005). Indeed, several African groups are known to have dispersed by this land bridge (e.g. aardvarks) (Koufos et al. 2005; Lehmann 2009), a situation that could have applied also to African squamates. No chamaeleonid fossils are found after the middle Miocene, but a latest immigration event seems to have taken place much more recently, involving the extant species C. chamaeleon, which still inhabits several parts of the Mediterranean Europe: a sporadic fossil occurrence from the Holocene of Spain (Talavera and Sanchíz 1983) probably supports this recent wave of dispersal. This latest dispersal event seemingly concerns also the extant Greek populations of C. chamaeleon in the islands of Samos and Chios, right adjacent to the coasts of Anatolia; the (recently extinct) population of C. chamaeleon in Crete, as also the other extant species occurring in Greece, C. africanus, is most probably a product of human introduction during antiquity (Böhme et al. 1998; Dimaki et al. 2015).

Fossil chamaeleonids from Europe. See Online Resource for a list of localities and taxa identified. Grey and encircled areas represent the range of extant chameleons in Europe and Western Anatolia, including autochthonous, introduced and recently extinct populations; the dashed circle indicates the probably introduced in antiquity population of C. africanus near Pylos, whereas the remaining areas refer to C. chamaeleon. Distributional data come from Gasc et al. (1997), Sindaco and Jeremčenko (2008), Sperone et al. (2010) and Sillero et al. (2014)
With such a scarce fossil record and a rather small temporal and regional European distribution, the dispersal routes of the earliest chamaeleonids remain unclear. Čerňanský et al. (2015) suggested that chamaeleonids (probably along with cordylids) entered Europe no earlier than the MN 3, but they did not suggest any possible route from Africa to Europe. Here we propose three different possible routes (Fig. 3): (i) a marine dispersal from northwestern Africa to southwestern Europe; (ii) a land dispersal from Afro-Arabia to Asia and then via Anatolia to southeastern Europe; and (iii) a land dispersal from Afro-Arabia to Asia and then northwards and eastwards, with an entrance in Europe through Central Asia.

Map showing the three biogeographic scenarios described in the text: the first one is represented by a black arrow, the second one by white arrows and the third one by grey arrows. Palaeogeographic map modified from Rögl 1999
The first scenario involves a marine dispersal from Africa to Europe across the Mediterranean. Such oceanic dispersals are well documented in chamaeleonid biogeographic history with molecular data suggesting multiple waves of colonization in Madagascar, Comoros, Seychelles and Socotra (Tolley et al. 2013; Tolley and Menegon 2013). Squamates in general are considered to be very efficient in trans-oceanic dispersal, with multiple lineages colonizing even the furthest and most remote of the Pacific Ocean islands (Beckon 1992; Fisher 1997; Pregill and Steadman 2004; Zug et al. 2011). These data, along with the extant distribution of chamaeleonids overseas in the Indian Ocean, suggest that the Palaeogene and Early Neogene Mediterranean and Paratethys seas (especially in the western part that is estimated to be rather narrower [Rögl 1999]) should not appear as a severe obstacle for these capable marine navigators. However, their absence from Palaeogene and early Miocene localities from Western Europe could possibly indicate that chamaeleonids did not disperse from this direction. Indeed, the extensive sampling of Western European localities for over a century could probably attest that this absence of chamaeleonids from this region is genuine and not sample biased. The supposed record of an indeterminate chamaeleonid from the early Miocene (MN 3/MN 4) of Agramon is tantalizing, since it could represent the only exception; however, the specimen has not been formally described and its whereabouts are currently unknown (Zbigniew Szyndlar, personal communication, November 2015). We further note here that short-time existing land connections between Africa and Western Europe have also been suggested (Gheerbrandt and Rage 2006; Zarcone et al. 2010), however, without unveiling any dispersal of chamaeleonids to Europe. A natural or ecological barrier could have prevented these squamates from immigrating to Western Europe either through water rafting or any hypothetical land bridge.
The second scenario involves a land dispersal from Afro-Arabia to Asia and then via Anatolia to Europe. This is supposed to take place in the early Miocene through the so-called Gomphotherium Landbridge, a large landmass that arose after the collision of Afro-Arabia with Asia during the MN 3 stage (Rögl 1999; Koufos et al. 2005). This collision provided a connection between Africa and Asia, dividing the huge sea that previously prevented dispersal of terrestrial taxa from Africa to Eurasia. The Afro-Arabian plate acted therefore as a platform for these terrestrial groups that could afterwards use Anatolia and then the Balkans in order to disperse northwards to Central Europe. A similar route had already been mentioned for chameleons by Barbadillo et al. (1997). According to this scenario, Greece seems to have acted as the first ‘step’ of these African and/or Anatolian immigrants into Europe. Through the ‘Gomphotherium Landbridge’, numerous terrestrial tetrapod groups are documented to have dispersed out of Africa, most eminently proboscideans (Rögl 1999; Koufos et al. 2003, 2005). If chameleons used this dispersal scenario, then they were among the first immigrants out of Africa, accompanying proboscideans and other mammal groups; they then rapidly dispersed to Central Europe in a short time interval, as it is shown by the presence of chamaeleonids in Germany and the Czech Republic already by the MN 3 (Roček 1984; Fejfar and Schleich 1994). Interestingly, several other tetrapod groups (e.g. aardvarks, hominoids) dispersed out of Africa much later, using another land bridge that arose during the middle Miocene (MN 6) (Lehmann 2009), but this datum is younger than the majority of fossil occurrences of chamaeleonids in Europe. Different ecological barriers probably existed that prevented or favoured the dispersal of certain groups at different periods.
The third scenario involves a land dispersal of chamaeleonids from Afro-Arabia to Asia and then northwards and entrance in Europe through Central Asia. This should involve again the usage of the ‘Gomphotherium Landbridge’ but then after dispersal eastwards and not westwards to the Balkans. We note here that the collision of the Afro-Arabian plate with Asia divided the Paratethys Sea and isolated the Kotsakhurian Sea in the eastern Paratethys region (Fig. 6, in Rögl 1999). This would further put rather severe obstacles in the spread of the African immigrants to the North. Therefore, only a westwards and an eastwards direction for these immigrants would seem possible. If chamaeleonids took the eastward direction, that would mean that they had to disperse to southern Asia, spread to Central Asia and then, after getting around the huge perimeter of the Kotsakhurian Sea, enter Central Europe. That way, their entrance into Europe should take place at the level of the northwestern edge of the Kotsakhurian Sea. However, the ubiquitous absence of chamaeleonid fossils in the several known early Miocene localities from this region does not seem to favour this hypothesis.
We acknowledge that the available fossil material is limited in order to fully support any of the proposed dispersal routes. However, the new Greek specimens provide evidence in favour of our second proposed scenario: a route from Greece towards Central Europe. Moreover, the slightly older age of the Aliveri fauna compared to all other MN 4 European localities, and more specifically from the also chamaeleonid-bearing Dolnice, could indicate a step-wise pathway of these squamates from south to northwards, and hints as Greece being the first European territory to which the group initially arrived from Africa via Anatolia. The presence of chamaeleonids in MN 3 Central European localities (Merkur Nord and Wintershof West), which are slightly older than Aliveri, could indicate that their dispersal and colonization of Europe took place quite fast. The purported record of the chamaeleonid from Agramon (MN 3/MN 4) (Böhme and Ilg 2003), if verified, could hint for very similar dispersal routes of these squamates to proboscideans: the latter group of large mammals are known to have used the Gomphotherium Landbridge at the MN 3 stage and rapidly dispersed into the European continent, reaching the Iberian Peninsula at the beginning of MN 4 (Tassy 1990; Koufos et al. 2005). The idea, however, of a combination of both the first and the second biogeographic scenarios should not be ruled out: the presence of chamaeleonids from Aliveri in the Balkan Peninsula, along with that from Agramon in the Iberian Peninsula, may indicate that these squamates could have used different pathways to Europe, from both northwestern and northeastern Africa. Such complex, double or even multiple dispersal events are far from unknown for southern European squamates. To the contrary, they have been well documented for extant gekkotan lizards (Rato et al. 2010, 2011) and Neogene elapid snakes (Szyndlar and Rage 1990). As far as it concerns the latter, the cobra Naja iberica from Spain has been described as ‘undoubtedly an African invader’, which, however, most likely never crossed the Pyrenees barrier and spread to the rest of Europe, which was inhabited by different congeners (Szyndlar and Rage 1990, p. 398). Such barrier could have also prevented early Miocene Iberian chamaeleonids from dispersing northwards, if they indeed ever reached the Peninsula during the early Neogene.
The Aliveri fauna bears a close resemblance to several coeval early Miocene ones from Anatolia, indicating biogeographic correlation and possible dispersal routes (van den Hoek Ostende et al. 2015). Notably, the viverrid Euboictis from Aliveri seems to share close affinities with a rather primitive congener form from Sabuncubeli in Anatolia (early MN 3), pointing towards an Asian origin for this mammal group (Koufos et al. 2005; van den Hoek Ostende et al. 2015). In contrast, Aliveri shares with coeval Central European faunas several micromammals, such as the eomyids and the flying squirrels. Curiously, eomyids are not present in the Turkish fossil record and therefore may indicate the existence of an ecological barrier between southern Balkan Peninsula and Anatolia (van den Hoek Ostende et al. 2015). It is also characteristic that the Aliveri rodent Anomalomys aliveriensis shares congener species with the early Miocene of Dolnice, Czech Republic, which is also the type locality of C. andrusovi. It is therefore obvious that the Aliveri fauna comprised a faunal mosaic of Anatolian and Central European taxa, acting as a biogeographic passage between Anatolia and Central Europe. Frustratingly, however, no chamaeleonid has ever been reported from any of the Anatolian localities (van den Hoek Ostende et al. 2015). More specimens from the early Miocene of Greece and generally from the Balkans and Anatolia are needed in order to fully test our hypothesis about the dispersal routes of chamaeleonids. This is unfortunately hindered by the scarcity of such localities in Greece (Koufos 2006b), as also by the generally limited interest of palaeontologists in fossil reptile finds in the area (Georgalis and Kear 2013). The confirmed presence of chamaeleonids in the early Miocene of Greece, however, allows us to expect that other localities between the MN 3 and MN 4 time intervals in the southern Balkans and Anatolia will also yield similar fossil forms that will only then allow a more accurate reconstruction of the dispersal routes of these charismatic lizards.
Chamaeleo cf. andrusovi from Aliveri represents only the third reptile species identified from the early Miocene of Greece. Mesozoic and Palaeogene reptiles are yet unknown from this country. Chamaeleo cf. andrusovi, the pleurodire turtle Nostimochelone lampra and the pythonid snake Python euboicus are therefore the oldest reptiles in the Greek fossil record (Römer 1870; Georgalis et al. 2013; Georgalis and Kear 2013). As the Aliveri specimens confirm the presence of Chamaeleonidae in the area, it is possible to speculate that several other reptile groups that have a fossil record in the early Neogene of Central Europe, such as cordylids, elapids, boines and choristoderans (Rage 1984; Szyndlar and Rage 1990, 2003; Szyndlar 1991; Evans and Klembara 2005; Rage and Szyndlar 2005; Čerňanský 2012), may be recovered from the Greek localities in the future. Especially for cordylids, it will be rather interesting to test whether this African lizard group (that has an almost identical, restricted fossil distribution in the early Miocene of Central Europe with chamaeleonids [Roček 1984; Čerňanský 2012]) used the same dispersal routes and if they can be identified in Greece and the rest of southeastern Europe as well. Unlike chameleons, however, cordylids are known to have also a Palaeogene fossil record from Europe; therefore, their presence in the same early Miocene localities with chamaeleonids during the climatic optimum could suggest either a continuous presence from the Palaeogene onwards to the Miocene or indeed a Palaeogene extinction and a later dispersal event along with chameleons (Čerňanský 2012). More fossil specimens and a revision of the already named taxa under a modern phylogenetic context are definitely needed in order to shed more light on the European squamate palaeobiogeography.
References
Álvarez Sierra MA, Daams R, van der Meulen AJ (1987) The mammals from the Lower Miocene of Aliveri (Island of Evia, Greece). 7. The Eomyidae. Proc Koninkl Nederl Akad Wetensc Ser B 90(1):47–56
Andreone F, Mattioli F, Jesu R, Randrianirina JE (2001) Two new chameleons of the genus Calumma from north-east Madagascar, with observations on hemipenial morphology in the Calumma furcifer group (Reptilia, Squamata, Chamaeleonidae). Herpetol J 11(2):53–68
Augé M (1997) A consideration of the phylogenetic significance of acrodonty. Herpetol J 7:111–113
Barbadillo LJ, García-París M, Sanchiz B (1997) Orígenes y relaciones evolutivas de la herpetofauna ibérica. In: Pleguezuelos JM (ed.) Distribución y Biogeografía de los Anfibios y Reptiles en España y Portugal. Monografías de Herpetología, Vol. 3. Asociación Herpetológica Española y Universidad de Granada, pp 47–100
Bastir M, Böhme M, Sanchiz B (2014) Middle Miocene remains of Alytes (Anura, Alytidae) as an example of the unrecognized value of fossil fragments for evolutionary morphology studies. J Vertebr Paleontol 34:69–79
Beckon WM (1992) The giant Pacific geckos of the genus Gehyra: morphological variation, distribution, and biogeography. Copeia 1992(2):443–460
Blanc CP (1972) Les reptiles de Madagascar et des îles voisines. In: Battistini R, Vindard G (eds) Biogeography and ecology in Madagascar. Junk, The Hague, The Netherlands, pp 501–614
Böhme M (2003) The Miocene climatic optimum: evidence from ectothermic vertebrates of Central Europe. Palaeogeogr Palaeoclimatol Palaeoecol 195:389–401
Böhme M (2010) Ectothermic vertebrates (Actinopterygii, Allocaudata, Urodela, Anura, Crocodylia, Squamata) from the Miocene of Sandelzhausen (Germany, Bavaria) and their implications for environment reconstruction and palaeoclimate. Paläontol Z 84:3–41
Böhme M, Ilg A (2003) fosFARbase, www.wahre-staerke.com/ (accessed 1 November 2015)
Böhme W, Bonetti A, Chiras G (1998) The chameleons of the Greek mainland: taxonomic allocation and conservation needs of a second European species (Squamata: Sauria: Chamaeleonidae). Herpetozoa 11(1/2):87–91
Bolet A, Evans SE (2012) A tiny lizard (Lepidosauria, Squamata) from the Lower Cretaceous of Spain. Palaeontol 55(3):491–500
Bolet A, Evans SE (2013) Fossil history of chameleons. In: Tolley KA, Herrel A (eds) The biology of chameleons. University of California Press, Berkeley, pp 175–192
Bolliger T (1992) Kleinsäugerstratigraphie der miozänen Hörnilschüttung (Ostschweiz). Dokum Natur 75:1–297
Camp CL (1923) Classification of the lizards. Bull Am Mus Nat Hist 48:289–481
Čerňanský A (2010) A revision of chamaeleonids from the Lower Miocene of the Czech Republic with description of a new species of Chamaeleo (Squamata, Chamaeleonidae). Geobios 43:605–613
Čerňanský A (2011) A revision of the chameleon species Chamaeleo pfeili Schleich (Squamata; Chamaeleonidae) with description of a new material of chamaeleonids from the Miocene deposits of southern Germany. Bulletin of Geosciences, Czech Geological Survey 86(2):275–282
Čerňanský A (2012) The oldest known European Neogene girdled lizard fauna (Squamata, Cordylidae), with comments on Early Miocene immigration of African taxa. Geodiv 34(4):837–848
Čerňanský A, Rage J-C, Klembara J (2015) The Early Miocene squamates of Amöneburg (Germany): the first stages of modern squamates in Europe. J Syst Palaeont 13(2):97–128
Conrad JL (2008) Phylogeny and systematics of Squamata (Reptilia) based on morphology. Bull Am Mus Nat Hist 310:1–182
Cope ED (1864) On the characters of the higher groups of Reptilia Squamata—and especially of the Diploglossa. Proc Acad Nat Sci Philad 1864:224–231
Cornette R, Herrel A, Stoetzel E, Moulin S, Hutterer R, Denys C, Baylac M (2015) Specific information levels in relation to fragmentation patterns of shrew mandibles: do fragments tell the same story? J Archaeol Sci 53:323–330
Crottini A, Miralles A, Glaw F, Harris DJ, Lima A, Vences M (2012) Description of a new pygmy chameleon (Chamaeleonidae: Brookesia) from central Madagascar. Zootaxa 3490:63–74
Daudin FF (1802) Histoire Naturelle, Générale et Particulière des Reptiles; ouvrage faisant suit à l’Histoire naturelle générale et particulière, composée par Leclerc de Buffon; et rédigee par C.S. Sonnini, membre de plusieurs sociétés savantes, vol. 4, F Dufart, Paris
de Bruijn H, van der Meulen AJ, Katsikatsos G (1980) The mammals from the Lower Miocene of Aliveri (Island of Evia, Greece). Part 1: the Sciuridae. Proc Koninkl Nederl Akad Wetensc Ser B 83(3):241–261
Delfino M, Kotsakis T, Arca M, Tuveri C, Pitruzzella G, Rook L (2008) Agamid lizards from the Plio-Pleistocene of Sardinia (Italy) and an overview of the European fossil record of the family. Geodiv 30(3):641–656
Delfino M, Rage J-C, Bolet A, Alba DM (2013) Early Miocene dispersal of the lizard Varanus into Europe: reassessment of vertebral material from Spain. Act Palaeontol Pol 58(4):731–735
Dimaki M, Valakos ED, Legakis A (2000) Variation in body temperatures of the African Chameleon, Chamaeleo africanus Laurenti, 1768 and the Common Chameleon, Chamaeleo chamaeleon (Linnaeus, 1758). Belg J Zool 130(Suppl):89–93
Dimaki M, Hundsdörfer AK, Fritz U (2008) Eastern Mediterranean chameleons (Chamaeleo chamaeleon, Ch. africanus) are distinct. Amphibia-Reptilia 29:535–540
Dimaki M, Chondropoulos B, Legakis A, Valakos E, Vergetopoulos M (2015) New data on the distribution and population density of the African Chameleon, Chamaeleo africanus and the Common Chameleon, Chamaeleo chamaeleon in Greece. Hyla 2015(1):36–43
Dollion AY, Cornette R, Tolley KA, Boistel R, Euriat A, Boller E, Fernandez V, Stynder D, Herrel A (2015) Morphometric analysis of chameleon fossil fragments from the Early Pliocene of South Africa: a new piece of the chamaeleonid history. Sci Nat 102:2
Doukas CS (1986) The mammals from the Lower Miocene of Aliveri (Island of Evia, Greece). Part 5. The insectivores. Proc Koninkl Nederl Akad Wetensc Ser B 89(1):15–38
Doukas CS (2003) The MN4 faunas of Aliveri and Karydia (Greece). Coloq Paleontol 1(2003):127–132
Estes R (1983) Sauria Terrestria, Amphisbaenia. Encyclopedia of Paleoherpetology, part 10a. Gustav Fisher Verlag, Stuttgart/New York
Estes R, de Queiroz K, Gauthier J (1988) Phylogenetic relationships within Squamata. In: Estes R, Pregill G (eds) Phylogenetic relationships of the lizard families: essays commemorating Charles L Camp. Stanford University Press, Stanford, pp 99–118
Evans SE, Klembara J (2005) A choristoderan reptile (Reptilia: Diapsida) from the Lower Miocene of Northwest Bohemia (Czech Republic). J Vertebr Paleont 25:171–184
Fejfar O, Schleich HH (1994) Ein Chamäleonfund aus dem unteren Orleanium des Braunkohlen-Tagebaus Merkur-Nord (Nordböhmen). Cour Forschungsinst Senck 173:167–173
Fisher RN (1997) Dispersal and evolution of the Pacific Basin gekkonid lizards Gehyra oceanica and Gehyra mutilata. Evol 51(3):906–921
Forskål P (1775) Descriptiones animalium, avium, amphibiorum, piscium, insectorum, vermium; quae in itinere Orientali observavit Petrus Forskål. Möller, Haunia
Gasc JP, Cabela A, Crnobrnja-Isailovic J, Dolmen D, Grossenbacher K, Haffner P, Lescure J, Martens H, Martinez Rica JP, Maurin H, Oliviera E, Sofianidou TS, Veith M, Zuiderwijk A (1997) Atlas of amphibians and reptiles in Europe. Societas Europea Herpetologica and Museum National d’Histoire Naturelle, Paris
Gauthier JA, Kearney M, Maisano JA, Rieppel O, Behlke ADB (2012) Assembling the squamate tree of life: perspectives from the phenotype and the fossil record. Bull Peab Mus Nat Hist 53:3–308
Gehring P-S, Pabijan M, Ratsoavina FM, Köhler J, Vences M, Glaw F (2010) A Tarzan yell for conservation: a new chameleon, Calumma tarzan sp. n., proposed as a flagship species for the creation of new nature reserves in Madagascar. Salamandra 46(3):167–179
Gehring P-S, Ratsoavina FM, Vences M, Glaw F (2011) Calumma vohibola, a new chameleon species (Squamata: Chamaeleonidae) from the littoral forests of eastern Madagascar. Afr J Herpetol 60(2):130–154
Georgalis GL, Kear BP (2013) The fossil turtles of Greece: an overview of taxonomy and distribution. Geobios 46:299–311
Georgalis GL, Velitzelos E, Velitzelos D, Kear BP (2013) Nostimochelone lampra gen. et sp. nov., an enigmatic new podocnemidoidean turtle from the Lower Miocene of northern Greece. In: Brinkman D, Holroyd P, Gardner J (eds) Morphology and evolution of turtles: papers in honor of Eugene S Gaffney. Vol 3. Pleurodires. Springer, Dordrecht, The Netherlands, pp 277–287
Gheerbrandt E, Rage J-C (2006) Palaeobiogeography of Africa: how distinct from Gondwana and Laurasia. Palaeogeogr Palaeoclimatol Palaeoecol 241:224–246
Glaw F (2015) Taxonomic checklist of chameleons (Squamata: Chamaeleonidae). Vertebr Zool 65(2):167–246
Glaw F, Vences M (1994) A field guide to amphibians and reptiles of Madagascar. 3rd ed, Vences and Glaw Verlag, Koln, Germany
Glaw F, Köhler JR, Townsend TM, Vences M (2012) Rivaling the world’s smallest reptiles: discovery of miniaturized and microendemic new species of leaf chameleons (Brookesia) from northern Madagascar. PLoS ONE 7(2):e31314
Gray JE (1825) A synopsis of the genera of reptiles and amphibia, with a description of some new species. An Phil 10:193–217
Greenbaum E, Tolley KA, Joma A, Kusamba C (2012) A new species of chameleon (Sauria: Chamaeleonidae: Kinyongia), from the northern Albertine Rift, Central Africa. Herpetol 68(1):60–75
Haas G (1952) The fauna of layer B of the Abu Usba Cave. Isr Expl J 2:35–47
Hillenius D (1959) The differentiation within the genus Chamaeleo Laurenti 1768. Beaufortia 8(89):1–92
Hillenius D (1978a) Notes on chameleons. IV: A new chameleon from the Miocene of Fort Ternan, Kenya (Chamaeleonidae, Reptilia). Beaufortia 28:9–15
Hillenius D (1978b) Notes on chameleons. V: the chameleons of North Africa and adjacent countries, Chamaeleo chamaeleon (Linnaeus) (Sauria, Chamaeleonidae). Beaufortia 28:37–55.
Hooijer DA (1961) The fossil vertebrates of Ksâr’Akil, a Palaeolithic rock shelter in the Lebanon. Zool Verh 49:3–67
Hou L (1976) New materials of Palaeocene lizards of Anhui. Vertebr Palasiat 14(1):48–52
Klaver CJJ (1977) Comparative lung-morphology in the genus Chamaeleo Laurenti, 1768 (Sauria: Chamaeleonidae) with a discussion of taxonomic and zoogeographic implications. Beaufortia 25(327):167–199
Klein Hofmeijer G, de Bruijn H (1985) The mammals from the lower Miocene of Aliveri (Island of Evia, Greece)—the Spalacidae and Anomalomyidae. Proc Koninkl Nederl Akad Wetensc Ser B 88:185–198
Klein Hofmeijer G, de Bruijn H (1988) The mammals from the Lower Miocene of Aliveri (Island of Evia, Greece). Part 8: the Cricetidae. Proc Koninkl Nederl Akad Wetensc Ser B 91(2):185–204
Kosuch J, Vences M, Böhme W (1999) Mitochondrial DNA sequence data support the allocation of Greek mainland chameleons to Chamaeleo africanus. Amph-Rept 20:440–443
Koufos GD (2006a) Palaeoecology and chronology of the Vallesian (late Miocene) in the Eastern Mediterranean region. Palaeogeogr Palaeoclimatol Palaeoecol 234(2–4):127–145
Koufos GD (2006b) The Neogene mammal localities of Greece: faunas, chronology, and biostratigraphy. An Geol Pays Hel 4:183–214
Koufos GD, Zouros N, Mourouzidou O (2003) Prodeinotherium bavaricum (Proboscidea, Mammalia) from Lesvos island, Greece; the appearance of deinotheres in the Eastern Mediterranean. Geobios 36:305–315
Koufos GD, Kostopoulos D, Vlachou T (2005) Neogene/Quaternary mammalian migrations in Eastern Mediterranean. Belg J Zool 135(2):181–190
Kumazawa Y (2007) Mitochondrial genomes from major lizard families suggest their phylogenetic relationships and ancient radiations. Gene 388:19–26
Laurenti JN (1768) Specimen medicum, exhibens synopsin reptilium emendatam cum experimentis circa venena et antidota reptilium austracorum, quod authoritate et consensu. Joan Thomae, Vienna
Lehmann T (2009) Phylogeny and systematics of the Orycteropodidae (Mammalia, Tubulidentata). Zool J Linn Soc 155:649–702
Linnaeus C (1758) Systema naturae per regna tria naturae, secundum classes, ordines, genera, species, cum characteribus, differentiis, synonymis, locis, 10th edn. Laurentius Salvius, Stockholm
López Martinez N (1986) The mammals from the lower Miocene of Aliveri (Island of Evia, Greece). 6. The ochotonid lagomorph Albertona balkanica n. gen. n. sp. and its relationships. Proc Koninkl Nederl Akad Wetensc Ser B 89(2):177–194
Mangili G (1980) Fossils reptiles of Simonelli cave. Accad Naz Lincei Probl Attu Sc Cult Quaderno 249:121–122
Maul LC, Smith KT, Barkai R, Barash A, Karkanas P, Shahack-Gross R, Gopher A (2011) Microfaunal remains at Middle Pleistocene Qesem Cave, Israel: preliminary results on small vertebrates, environment and biostratigraphy. J Hum Evol 60(4):464–480
Młynarski M, Szyndlar Z, Estes R, Sanchiz B (1982) Lower vertebrate fauna from the Miocene of Opole (Poland). Estud Geol 38:103–119
Moody S, Roček Z (1980) Chamaeleo caroliquarti (Chamaeleonidae, Sauria) a new species from the Lower Miocene of central Europe. Věstnik Ústředniho Ustavu Geologického 55:85–92
Mörs T (2002) Biostratigraphy and paleoecology of continental Tertiary vertebrate faunas in the Lower Rhine Embayment (NW-Germany). Neth J Geosc, Geol Mijn 81:177–183
Mörs T, von der Hocht F, Wutzler B (2000) Die erste Wirbeltierfauna aus der miozänen Braunkohle der niederrheinischen Bucht (Ville-Schichten, Tagebau Hambach). Paläontol Z 74:145–170
Nečas P, Modry D, Slapeta JR (2003) Chamaeleo (Trioceros) narraioca n. sp. (Reptilia: Chamaeleonidae), a new chameleon species from a relict montane forest of Mount Kulal, northern Kenya. Trop Zool 16(1):1–12
Nečas P, Modry D, Slapeta JR (2005) Chamaeleo (Triceros) ntunte n. sp., a new chamaeleon species from Mt. Nyiru, northern Kenya (Squamata: Sauria: Chamaeleonidae). Herpetozoa 18(3/4):125–132
Nečas P, Sindaco R, Kořený L, Kopečná J, Malonza PK, Modry D (2009) Kinyongia asheorum sp. n., a new montane chameleon from the Nyiro Range, northern Kenya (Squamata: Chamaeleonidae). Zootaxa 2028:41–50
Oppel M (1811) Die Ordnungen, Familien, und Gattungen der Reptilien als Prodrom einer Naturgeschichte derselben. Joseph Lindauer, München
Pickford M (1986) Sediment and fossil preservation in the Nyanza Rift system of Kenya. Geol Soc Spec Pub 25:345–362
Pregill GK, Steadman DW (2004) South Pacific iguanas: human impacts and a new species. J Herpetol 38(1):15–21
Prieto J, Böhme M, Maurer H, Heissig K, Abdul-Aziz H (2009) Biostratigraphy and sedimentology of the Fluviatile Untere Serie (Early and Middle Miocene) in the central part of the North Alpine Foreland Basin: implications for palaeoenvironment and climate. Int J Earth Sci 98(7):1767–1791
Pyron RA, Burbrink FT, Wiens JJ (2013) A phylogeny and revised classification of Squamata, including 4161 species of lizards and snakes. BMC Evol Biol 13:93
Rage J-C (1984) Serpentes. Handbuch der Palaeoherpetologie, Part 11. Gustav Fischer Verlag, Stuttgart/New York
Rage J-C, Bailon S (2011) Amphibia and Squamata. In: Harrison T (ed) Paleontology and geology of Laetoli: human evolution in context, vol 2, Fossil hominins and the associated fauna. Springer, New York, pp 467–478
Rage J-C, Szyndlar Z (2005) Latest Oligocene-Early Miocene in Europe: Dark Period for booid snakes. Compt R Pal 4:428–435
Raselimanana AP, Rakotomalala D (2003) Chamaeleonidae, chamaeleons. In: Goodman SM, Benstead JP (eds) The natural history of Madagascar. University of Chicago Press, Chicago, pp 960–969
Rato C, Carranza S, Perera A, Carretero MA, Harris DJ (2010) Conflicting patterns of nucleotide diversity between mtDNA and nDNA in the Moorish gecko, Tarentola mauritanica. Mol Phyl Evol 56:962–971
Rato C, Carranza A, Harris DJ (2011) When selection deceives phylogeographic interpretation: the case of the Mediterranean house gecko, Hemidactylus turcicus (Linnaeus, 1758). Mol Phyl Evol 58:365–373
Raxworthy CJ, Forstner MRJ, Nussbaum RA (2002) Chameleon radiation by oceanic dispersal. Nature 415:784–787
Reeder TW, Townsend TM, Mulcahy DG, Noonan BP, Wood PL Jr, Sites JWJ, Wiens JJ (2015) Integrated analyses resolve conflicts over squamate reptile phylogeny and reveal unexpected placements for fossil taxa. PLoS ONE 10(3):e0118199
Rieppel O, Walker A, Odhiambo I (1992) A preliminary report on a fossil chamaeleonine (Reptilia: Chamaeleoninae) skull from the Miocene of Kenya. J Herpet 26(1):77–80
Roček Z (1984) Lizards (Reptilia: Sauria) from the Lower Miocene locality Dolnice (Bohemia, Czechoslovakia). Rozpr Českosl Akad věd, řad matem přír věd 94(1):3–64
Rögl F (1999) Mediterranean and Paratethys. Facts and hypotheses of an Oligocene to Miocene paleogeography (short overview). Geol Carpat 50:330–349
Römer F (1870) Über Python euboicus, eine fossile Riesenschlange aus tertiärem Kalkschiefer von Kumi auf der Insel Euboea. Z Deuts Geol Ges 22:582–590
Schleich HH (1983) Die mittelmiozane Fossil-Lagerstätte Sandelzhausen 13. Chamaeleo bavaricus sp. nov., ein neuer Nachweis aus dem Jungtertiär Süddeutschlands. Mitt Bayer Staatssamml Paläontol Hist Geol 23:77–81
Schleich HH (1984) Neue Reptilienfunde aus dem Tertiär Deutschlands 2. Chamaeleo pfeili sp. nov., von der untermiozänen Fossilfundstelle Rauscheröd/Niederbayern (Reptilia, Sauria Chamaeleonidae). Mitt Bayer Staatssamml Paläontol Hist Geol 24:97–103
Schleich HH (1994) Neue Reptilienfunde aus dem Tertiär Deutschlands 15. Neue Funde fossiler Chamäleonen aus dem Neogen Süddeutschlands. Cour Forschungsinst Senckenb 173:175–195
Schmidt-Kittler N (1983) The mammals from the Lower Miocene of Aliveri (Island of Evia, Greece). 3. On a new species of Sivanasua Pilgrim, 1931 (Feliforma, Carnivora) and the phylogenetic position of this genus. Proc Koninkl Nederl Akad Wetensc Ser B 86(3):301–318
Sillero N, Campos J, Bonardi A, Corti C, Creemers R, Crochet P-A, Crnobrnja Isailović J, Denoël M, Ficetola GF, Gonçalves J, Kuzmin S, Lymberakis P, de Pous P, Rodríguez A, Sindaco R, Speybroeck J, Toxopeus B, Vieites DR, Vences M (2014) Updated distribution and biogeography of amphibians and reptiles of Europe. Amphibia-Reptilia 35:1–31
Simões TR, Wilner E, Caldwell MW, Weinschütz LC, Kellner AWA (2015) A stem acrodontan lizard in the Cretaceous of Brazil revises early lizard evolution in Gondwana. Nat Com 6:8149
Sindaco R, Jeremčenko V (2008) The reptiles of the western Palearctic. Vol. 1: annotated checklist and distributional atlas of the turtles, crocodiles, amphisbaenians and lizards of Europe, North Africa, Middle East and Central Asia. In: Belvedere (ed) Monografie della Societas Herpetologica Italica - I
Sperone E, Crescente A, Brunelli E, Paolillo G, Tripepi S (2010) Sightings and successful reproduction of allochthonous reptiles in Calabria. Act Herpetol 5(2):265–273
Stipala J, Lutzmann N, Malonza PK, Borghesio L, Wilkinson P, Godley B, Evans MR (2011) A new species of chameleon (Sauria: Chamaeleonidae) from the highlands of northwest Kenya. Zootaxa 3002:1–16
Stipala J, Lutzmann N, Malonza PK, Wilkinson P, Godley B, Nyamache J, Evans MR (2012) A new species of chameleon (Squamata: Chamaeleonidae) from the Aberdare Mountains in the central highlands of Kenya. Zootaxa 3391:1–22
Szyndlar Z (1991) A review of Neogene and Quaternary snakes of Central and Eastern Europe. Part I: Scolecophidia, Boidae, Colubrinae. Est Geol 47:103–126
Szyndlar Z, Rage J-C (1990) West Palearctic cobras of the genus Naja (Serpentes: Elapidae): interrelationships among extinct and extant species. Amphibia-Reptilia 11:385–400
Szyndlar Z, Rage J-C (2003) Non-erycine Booidea from the Oligocene and Miocene of Europe. Institute of Systematics and Evolution of Animals. Polish Academy of Sciences, Cracow
Talavera R, Sanchíz F (1983) Restos pliocénicos de Camaleón común, Chamaeleo chamaeleon (L.) de Málaga. Bol Real Soc Esp Hist Nat (Geolo) 81:81–84
Tassy P (1990) The “Proboscidean datum event”: how many proboscideans and how many events. In: Lindsay EH, Fahlbusch V, Mein P (eds) European Neogene mammal chronology. Plenum Press, New York, pp 237–252
Teyssier J, Saenko SV, van der Marel D, Milinkovitch MC (2015) Photonic crystals cause active colour change in chameleons. Nat Com 6:6368
Tilbury CR, Tolley KA (2015) Contributions to the herpetofauna of the Albertine Rift: two new species of chameleon (Sauria: Chamaeleonidae) from an isolated montane forest, south eastern Democratic Republic of Congo. Zootaxa 3905:345–364
Tolley KA, Menegon M (2013) Evolution and biogeography of chameleons. In: Tolley KA, Herrel A (eds) The biology of chameleons. University of California Press, Berkeley, pp 131–147
Tolley KA, Chase BM, Forest F (2008) Speciation and radiations track climate transitions since the Miocene climatic optimum: a case study of southern African chameleons. J Biogeogr 35:1402–1414
Tolley KA, Townsend TM, Vences M (2013) Large-scale phylogeny of chameleons suggests African origins and Eocene diversification. Proc R Soc B 280:20130184
Townsend T, Larson A, Louis EJ, Macey JR (2004) Molecular phylogenetics of Squamata: the position of snakes, amphisbaenians, and dibamids, and the root of the squamate tree. Syst Biol 53:735–757
Townsend TM, Mulcahy DG, Noonan BP, Sites JW Jr, Kuczynski CA, Wiens JJ, Reeder TW (2011) Phylogeny of iguanian lizards inferred from 29 nuclear loci, and a comparison of concatenated and species-tree approaches for an ancient, rapid radiation. Mol Phyl Evol 61:363–380
van Den Hoek Ostende LW, Mayda S, Oliver A, Madern A, Hernández-Ballarín V, Peláez-Campomanes P (2015) Aliveri revisited, a biogeographical appraisal of the early Miocene mammals from the eastern Mediterranean. Palaeobiod Palaeoenv 95:271–284
van der Meulen AJ, de Bruijn H (1982) The mammals from the Lower Miocene of Aliveri (Island of Evia, Greece). Part 2. The Gliridae. Proc Koninkl Nederl Akad Wetensc Ser B 85(4):485–524
Vidal N, Hedges SB (2009) The phylogeny of squamate reptiles (lizards, snakes, and amphisbaenians) inferred from nine nuclear protein-coding genes. C R Biol 328:1000–1008
Wiens JJ, Brandley MC, Reeder TW (2006) Why does a trait evolve multiple times within a clade? Repeated evolution of snake-like body form in squamate reptiles. Evol 61:123–141
Wiens JJ, Hutter CR, Mulcahy DG, Noonan BP, Townsend TM, Sites JW, Reeder TW (2012) Resolving the phylogeny of lizards and snakes (Squamata) with extensive sampling of genes and species. Biol Lett 8:1043–1046
Zarcone G, Petti FM, Cillari A, Di Stefano P, Guzzetta D, Nicosia U (2010) A possible bridge between Adria and Africa: new palaeobiogeographic and stratigraphic constraints on the Mesozoic palaeogeography of the Central Mediterranean area. Earth-Sci Rev 103:154–162
Zug GR, Hamilton AM, Austin CC (2011) A new Emoia samoensis group lizard (Squamata: Scincidae) from the Cook Islands, South-central Pacific. Zootaxa 2675:47–57
Acknowledgments
We are grateful to Wilma Wessels (University of Utrecht) and Hans de Bruijn (University of Utrecht) for the extended loan of the specimens. Walter Joyce (University of Fribourg), Dimitris Kostopoulos (Aristotle University of Thessaloniki), George Koufos (Aristotle University of Thessaloniki) and Zbigniew Szyndlar (Institute of Systematics and Evolution of Animals, Polish Academy of Sciences, Krakow) provided important comments that improved the manuscript. Andrej Čerňanský (Museum für Naturkunde, Berlin) and three anonymous reviewers gave significant input that enhanced the quality of the paper. Gaetano Pitruzzella prepared the interpretative drawing of UU AL 3501 showed in Fig. 1. Project supported by Fondi di Ateneo (ex 60 %) 2013–2014 dell’ Università di Torino and Generalitat de Catalunya (2014 SGR 416 GRC). Access to the collections of the Muséum national d’Histoire naturelle, Paris, has been made possible thanks to a Synthesys grant (FR-TAF-5007) to AV. GLG acknowledges travel funding from the University of Fribourg.
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by: Sven Thatje
Electronic supplementary material
Below is the link to the electronic supplementary material.
ESM 1
(PDF 101 kb)
Rights and permissions
About this article

Cite this article
Georgalis, G.L., Villa, A. & Delfino, M. First description of a fossil chamaeleonid from Greece and its relevance for the European biogeographic history of the group. Sci Nat 103, 12 (2016). https://doi.org/10.1007/s00114-016-1336-5
Received:
Revised:
Accepted:
Published:
DOI: https://doi.org/10.1007/s00114-016-1336-5