
Abstract
Previously, the presence of metals in arthropod mandibles has been linked with harder cuticle, and in termites, a 20% increase in hardness has been found for mandibles containing major quantities of zinc. The current study utilises electron microscopy and energy-dispersive X-ray microanalysis to assess incidence and abundance of metals in all extant subfamilies of the Isoptera. The basal clades contain no zinc and little to no manganese in the cutting edge of the mandible cuticle, suggesting that these states are ancestral for termites. However, experimentation with mandibles in vitro indicates the presence of some elements of the cuticular biochemistry necessary to enable uptake of zinc. The Termopsidae, Serritermitidae, Rhinotermitidae and Termitidae all contain minor quantities of manganese, while trace to minor quantities of zinc occur in all except the Serritermitidae. In contrast, all Kalotermitidae or drywood termites contain major levels of zinc in the mandible edge. Diet and life type are explored as links to metal profiles across the termites. The presence of harder mandibles in the drywood termites may be related to lack of access to free water with which to moisten wood. Scratch tests were applied to a set of mandibles. The coefficient of friction for Cryptotermes primus (Kalotermitidae) mandibles, when compared with species from other subfamilies, indicates that zinc-containing mandibles are likely to be more scratch resistant.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
A range of metals including zinc (Zn) and manganese (Mn) are found in insects and other arthropods concentrated within the cuticle (or exoskeleton) of mandibles, mouth hooks, claws and ovipositors (Hillerton and Vincent 1982; Hillerton et al. 1984; Quicke et al. 1998; Fontaine et al. 1991; Schofield 2001; Lichtenegger et al. 2003; Morgan et al. 2003; Schofield 2005; Birkedal et al. 2006). A number of studies on insects establish links between Zn enrichment and enhanced mechanical properties such as hardness, for example in grasshoppers, Schistocerca gregaria and Locusta migratoria (Orthoptera) and the leaf-cutter ant, Atta sexdens rubipilosa (Hymenoptera) (Edwards et al. 1993; Schofield et al. 2002; Hillerton et al. 1982). Zn in the jaws has also been linked with increased local hardness (and stiffness) in the marine polychaete worm Nereis limbata (Lichtenegger et al. 2003). In termites, the metals, Zn and Mn, occur only in the edge of the mandibles and not below in the general cuticle of the mandible (Cribb et al. 2007). Recently, it was found that the mandible edges of termites which contain major quantities of Zn are 20% harder than mandibles without this metal (Cribb et al. 2007). Interestingly, of the six species of termite investigated for hardness in this study, the two species found to contain Zn were from the same group, the Kalotermitidae or drywood termites. Beyond the six species analysed in that study, only four other species of termites have been investigated for metals in mandibles and while some show minor quantities of Mn but no Zn (Fawke et al. 1997; Yoshimura et al. 2002, 2005), the two others are drywood termite species and show Zn in the mandible edge (Ohmura et al. 2007).
Because Zn-enriched mandibles are substantially harder, their presence may relate to capacity to inflict damage to wood and, hence, pest status. Presence may also relate to specific life type characteristics. The question arises as to the distribution of Zn occurrence within the termites. In the current study, we survey all extant subfamilies in the Isoptera: the Mastotermitidae, Hodotermitidae, Termopsidae, Serritermitidae, Rhinotermitidae and Termitidae. It should be noted that we have chosen to maintain the classification schema used prior to Inward et al. (2007) due to the as yet unresolved debate initiated by Lo et al. (2007).
Materials and methods
Samples were assessed for elemental composition using a JEOL JSM 6460 LA low vacuum analytical scanning electron microscope equipped with an integrated JEOL Hyper mini-cup, 133eV resolution, ultra thin window (UTW), SiLi crystal, energy-dispersive x-ray spectrometer (EDS) following Cribb et al. (2007). X-ray analysis was carried out in low vacuum mode to obviate a metal coating. Standardless quantitation software comprised the JEOL Analysis Station (V3.51). Acquisition conditions on the scanning electron microscope (SEM) were 20 kV, 10 mm working distance and 30 s live time acquisition at approximately 10–15% dead time. Because no suitable standard is available to match the material being assessed, standardless quantitation was used with a phi-rho-z correction and elemental mass reported qualitatively as trace (<0.5 mass % dry weight), minor (a few percent) or major (>5–≥20% mass weight; following Newbury 1991).
Samples from all extant subfamilies of the Isoptera were investigated for metal content at the working mandible edge. Multiple samples for each species were assessed with 3–30 worker individuals measured. The exception was Coptotermes formosanus where only a single worker was available; however, this species has been investigated previously (Yoshimura et al. 2002, 2005). A minimum of three EDS measurements were taken from each sample. Individuals selected for sampling were well tanned (showed dark head capsules and mandibles).
Soil samples were taken from seven termite collection sites around South East Queensland (Australia). These were dried at 60°C and sieved to >63 μm, <2 and <2 mm. Samples (1 g) were digested using aqua regia following the Australian Standard method (AS 4479.2). The digest solutions were filtered and made up with 1% nitric acid. Analysis of Mn and Zn were performed by inductively coupled plasma optical emission spectrometry. Standard reference materials used were River Sediments NCS CRF NCS DC78301 and GBW08301. All analyses were performed by Queensland Health Scientific Services National Association of Testing Authorities registered (ISO 17025-accredited) Inorganic Chemistry laboratory at Coopers Plains, Queensland, Australia.
A colony that was known to be devoid of metals at the mandible edge of worker individuals was selected for study. Thirty Mastotermes darwiniensis (Mastotermitinae) mandibles were harvested and treated in one of three ways. One set was exposed only to water for the duration of the experiment, the second set was exposed to 1 M aqueous zinc acetate (C2H3O2)2Zn: Sigma Aldrich) for 12 h and the third set was pretreated with 1 M (4%) aqueous sodium hydroxide (NaOH: Ajax) at 60°C for 1 h followed by 2 h at room temperature before being washed in deionised water and then exposed to the 1 M zinc acetate for 12 h at room temperature. All samples were then washed in deionised water to remove external Zn and air-dried. Mandible edges were assessed using X-ray analysis (EDS) as above.
Using the Hysitron Triboindentor, scratch tests were applied to a set of termite mandibles comprising three each of Cryptotermes primus (Kalotermitidae), Coptotermes acinaciformis ((Rhinotermitidae) and M. darwiniensis (Mastotermitidae). Samples were embedded in Daystar resin, polymerised at room temperature then polished flat into the longitudinal plane of the tip region. All tests were at room temperature, and the samples were firmly fixed to the holder. A conical diamond probe tip with a tip radius of 1 μm and a cone angle of 90° was applied with a normal load of 300 μN and a scratch length of 10 μm. Penetration into the sample varied from 150 to 200 nm and SEM observation on samples after testing showed no pileup and no gouging. Mass percentage of elements (e.g. Zn and Mn) was confirmed after scratch testing using EDS. Scratch morphology was investigated using secondary electron imaging on a JEOL 6300 field emission SEM.
Results
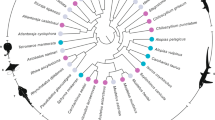
Metal content in the mandible edges is consistent within certain groups (Table 1). A summary of the metal content by subfamily and life type is presented in Fig. 1. Major Zn occurrence appears to be present across the whole Kaloterminitidae (drywood termites). It should be noted that representatives of all major groups in the drywood termites have been assessed (Fig. 2).

Majority rule consensus phylogenetic tree to subfamily level for termites (following Eggleton 2001; Lo et al. 2007) with metal profile and life history data for workers (following Abe 1987; Eggleton and Tayashu 2001; unpublished collection data). Metal concentration for Zn and Mn is mass, reported qualitatively as trace (<0.5 mass % dry weight), minor (a few percent) or major (>5 to ≥20% mass weight)

Australian lineage of the Kalotermitidae or drywood termites (following Thompson et al. 2000) indicating the branches investigated for metal incorporation (bolded). All branches studied showed major Zn occurrence in the mandible edges of workers. Data on the metal incorporation is provided in Table 1
Soil sample data were collected from seven sites where Rhinotermitidae, Termitidae and Kalotermitidae were taken. All sites showed presence of both Mn and Zn, but concentrations varied from 17 to 1,988 mg/kg (dry weight) for Mn and 5–211 mg/kg (dry weight) for Zn. However, there was no correlation between metals in the mandibles of the termites and soil. The Pearson’s correlation coefficients are as follows: for Mn in the mandible against Mn in the soil r 9 = −0.078, P = 0.841; for Zn in the mandible against Zn in the soil r 9 = −0.040, P = 0.920.
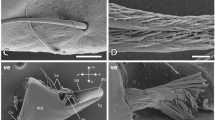
Representative X-ray analysis (EDS) spectra are presented in Fig. 3. The halogen chlorine (Cl) is always associated with the presence of Zn (e.g. Fig. 3b: Cryptotermes brevis) whereas, when Mn is present alone, while silicon (Si) is still seen, Cl is not always present (e.g. Fig. 3c: Nasutitermes magnus). The mandibles of M. darwiniensis investigated for Zn uptake in vitro did not contain Mn or Zn (Fig. 3d); however, some regions showed minor quantities of sulphur (S). Representatives from other colonies of M. darwiniensis showed trace Mn (Table 1, Fig. 1). The grinding plate, cutting edge and tooth or teeth characteristic of wood-feeding termites are visible in the mandible of M. darwiniensis (Fig. 4a). Sections through the apical tip region and grinding plate were assessed for Zn uptake using EDS and backscattered electron microscopy (Figs 3d–g, 4b–d). In drywood termites, the cuticle in these regions was seen to contain multiple small pore canals as well as larger pore canals (see Cribb et al. 2007). These large canals are also evident in transverse regions of M. darwiniensis mandible edges (Fig. 4b–d).

Elemental composition as representative spectra from energy-dispersive X-ray analysis of termite mandibles; a Daystar polyester resin mounting-medium (control); b Spectrum demonstrating Zn (major quantity) in Cryptotermes brevis; c Mn (minor) in Nasutitermes magnus; d no metals in this sample from Mastotermes darwiniensis; e M. darwiniensis mandible tip treated with NaOH prior to zinc acetate (see Fig. 4c for image) f M. darwiniensis mandible grinding plate treated with NaOH prior to zinc acetate g M. darwiniensis mandible below edge, treated with NaOH prior to zinc acetate

Mandibles of Mastotermes darwiniensis. a Scanning electron micrograph showing the posterior end a grinding plate (G), anterior cutting edges (C) and tooth (T). Bar: 100 μm. b–d Transverse sections through mandible tips and grinding plate. Images obtained using backscattered electron imaging: atomic number is correlated with contrast (see J. Goldstein et al. 2003). Arrows indicate large pore canals. b Mandible tip: treated with zinc acetate alone. Bar: 10 μm. c Mandible tip: treated with sodium hydroxide prior to zinc acetate. Metal-enriched areas are whiter than surrounding regions. Bar: 10 μm. d Grinding plate: treated with zinc acetate alone. Bar: 20 μm
Mandibles treated with aqueous zinc acetate, or immersed only in water, did not take up Zn within the cuticle (see Figs. 3d, 4b,d). Only mandibles pretreated with 4% NaOH solution before exposure to aqueous zinc acetate solution (an uncomplexed form of zinc) were seen to bind Zn (Figs. 3e, 4c), and the enrichment was not homogeneous. X-ray analysis (EDS) demonstrated that major Zn uptake occurred principally in the mandible edge with minor uptake throughout the adjacent insect cuticle (contrast Fig. 3e,f,g).
The coefficient of friction was assessed for termite mandibles using a scratch test. This coefficient is defined as the ratio of tangential to normal force (F t /F n ). Values varied as follows—Cr. primus (Kalotermitidae) averaged 0.25 with a range of 0.24–0.27, M. darwiniensis (Mastotermitidae) averaged 0.27 with a range of 0.25–0.28 and C. acinaciformis (Rhinotermitidae) averaged 0.29 with a range of 0.25–0.30. The data sets from each of the three species tested showed overlap but the lowest friction measurements were observed from mandibles of Cr. primus. Microanalysis via X-ray analysis confirmed that the Cr. primus mandibles contained Zn at major levels, the C. acinaciformis mandibles contained Mn at minor levels and the M. darwiniensis mandibles contained no Zn or Mn.
Discussion
Incorporation of Zn and Mn occurs throughout phylogenies within the Arthropoda suggesting its advent predates the divergence of the Insecta (Schofield et al. 2003). Termites (Isoptera) are ancestrally related to wood-feeding cockroaches and have existed for over 100 million years (Pearce 1997; Abe et al. 2000; Eggleton and Tayashu 2001; Inward et al. 2007; Lo et al. 2007). However, while we have shown that termite mandibles generally contain metals, cockroaches that have been investigated do not incorporate Zn or Mn into their mandibles (Hillerton and Vincent 1982). Within the Isoptera, our study shows metal incorporation maps onto phylogenetic position and life type. Mastotermitidae and Hodotermitidae contain no Zn and little to no Mn (Fig. 2) suggesting that these states are ancestral for termites: the two taxa are considered basal clades (Pearce 1997). The remaining five subfamilies, in addition to accumulating low levels of Mn, all show the capacity for acquisition of Zn in the mandible edge, except for the Serritermitidae (found only in Brasil). The feeding habit of the Serritermitidae is not known and may be atypical, involving predation and scavenging (Emerson and Krishna 1975). These unusual termites have reduced grinding features on the mandibles unlike those seen in wood-feeders (Emerson and Krishna 1975). Here, the absence of Zn may represent a secondary loss and is possibly related to adaptation to a different life history; alternatively, the data also support multiple acquisitions for Zn in the phylogenetic tree because some members of the sister groups, Rhinotermitidae and Termitidae, also show an absence of Zn in the mandibles.
There are examples from other taxa suggesting similar loss of metal acquisition. In the Hymenoptera, mandibles from wasps in the Vespidae and Pelecinidae lack Zn (Quicke et al. 1998). In the Coleoptera, lack of metals in Carabid mandibles may be a secondary loss (Hillerton et al. 1984). However, dietary deficiency cannot be ruled out as a cause for absence of metals (Morgan et al. 2003). To address this possibility, our study considered dietary sufficiency. We found colonies within the Rhinotermitidae (Coptotermes acinaciformis) that lacked Zn but were co-located with a colony from the Kalotermitidae (Cryptotermes primus) which contained major amounts of Zn in the mandible edges. Both species were feeding on the same wood and presumably had access to equivalent dietary Zn, which, for the kalotermitids, was a sufficient food source on its own for Zn enrichment.
Some data that indicate concentration of available Mn in the environment may influence uptake in insects (Morgan et al. 2003 ). We sampled and tested soil from termite collection sites to determine whether soil concentration might be linked to metals present in mandibles. Presumably, vegetation supported by the soil would have had access to the mineral profile but the plant material was not evaluated. Sites tested for a range of termites collected in South East Queensland (Australia) all showed presence of Mn and Zn, but concentrations varied and there was no correlation between soil concentration and qualitative concentration in the mandibles. This suggests that metal uptake is not linked to environmental levels, beyond the need for the metal to be present at some concentration.
Interestingly, within a few of the species sampled, the metal profile was seen to vary. For example, some collections of C. acinaciformis showed minor levels of Zn in their mandibles whereas others lacked the metal (Table 1). However, evidence based on the mitochondrial cytochrome oxidase gene suggests this species probably represents a complex (Lo et al. 2006). So, such diversity within our sample set may explain the difference in the observed metal accumulation. The disparity highlights possible physiological differences. It is worth noting, though, that the species complex is expected to feed in a similar fashion so those members accumulating Zn at a low level would not be likely to represent a change in feeding strategy.
Because high Zn levels have been linked with enhanced hardness, acquisition within the Insecta suggests adaptation to extreme environments where improvements in ‘tools’ is an advantage. An elegant study by Schofield et al. (2002) using leaf-cutter ants supports this hypothesis. The authors show that ants only cut leaves when the mandible edges increase in hardness, which is accompanied by the incorporation of Zn. Other examples also exist. Some beetles, like Sitophilus granarius, the granary beetle, have Mn in adult mandibles but not in the larval mandible (Morgan et al. 2003). In this species, the adult penetrates the seed for the larva, laying the egg within the grain (Morgan et al. 2003). Ichneumonid wasps like Megarhyssa spp, which contain Mn in the ovipositor, take an hour to drill an oviposition hole and lay one egg (Quicke et al. 1998). On average, such a wasp spends 20 h actively drilling. The first instar of the bug Leptoglossus occidentalis, which feeds on foliage, lack Mn, but the later instar and adult contain Mn in the rostrum and are seed feeders (Fontaine et al. 1991). It seems likely that the presence of major quantities of Zn in the drywood termites might also be linked to a life history or survival strategy found in this group.
Most termites require contact with water but the Kalotermitidae are referred to as the drywood termites because they consume sound wood in the absence of free water. They live in small family nests (a few hundred individuals) within single pieces of wood often well above the soil. Single-piece nesters, as this life type has been termed (Abe 1987), are small, confined colonies restricted in foraging choice because they do not leave the nest region to visit the soil or to scavenge on a range of cellulose sources. The most likely advantage of the specialised mandibles, therefore, relates to water use. Workers from other termite groups have access to free water and can use this. For example Reticulitermes santonensis (Rhinotermitidae) have been shown to regurgitate water onto substrates from their water sacs (labial gland reservoirs; Grube and Rudolph 1999). They are, therefore, able to bring water to dry wood surfaces under harvest. These glands also produce a general signal in termites that results in gnawing, feeding and aggregations at the labeled food site (Reinhard and Kaib 2001). In contrast, the Kalotermitidae are water limited because they only obtain water metabolically (Pearce 1997). We hypothesise that whereas most termites can feed on water-softened food and, hence, have no need for mandibles with increased hardness, drywood termites with no access to free water instead harden their mandibles. This may have resulted in two different strategies in the Isoptera to cope with a difficult food source: soften it with water and if this cannot be achieved, develop better tools.
However, groups other than the drywood termites also show uptake or accumulation of Zn, for example some genera within the higher termites or Termitidae, but these incorporate Zn to a lesser extent. This does indicate that a mechanism for Zn incorporation exists outside the Kalotermitidae. At this stage, no evaluation has been made of mandible hardness for minor concentrations of Zn. There can only be an assumption on the basis of studies from other arthropods that there is a correlation between hardness and concentration. The presence of a mechanism for Zn incorporation outside the Kalotermitidae allows that it could be acted upon by selective pressure in these organisms to produce higher concentrations (as seen in the kalotermitids) and presumably harder mandibles over evolutionary time.
But are termites that show no Zn uptake in possession of this mechanism also? The bonding characteristics for Zn in cuticle are poorly understood at present (Schofield 2005) with evidence from marine worms pointing towards possible involvement of proteins, specifically the metal-binding imidazole side chain of histidine (Lichtenegger et al. 2003, Broomell et al. 2007). The microstructure is better documented. Schofield et al. (2003) noted that arthropod cuticle with high densities of pore canals is associated with metal uptake and Compère et al. (1993) demonstrated experimentally that pore canals are the means by which biomineralisation precursors are transferred to the cuticle, in this case, calcium and magnesium ions, in crabs. So, presence of these canals can be taken as an indicator of cuticle that is associated with increasing metal mass. Cribb et al. (2007) noted pore canals in the mandible edges of Kalotermitidae. Both small and large canals were identified. In the current study, the mandible edges of Mastotermes darwiniensis appear similar. But this is not surprising because trace amounts of Mn have been found in the edges of some mandibles from this species. The canals are likely to be the physical transport system for translocation of metals. Active binding sites must also be present for incorporation to occur.
To explore further, we undertook an experiment aimed at testing whether mandibles of M. darwiniensis could take up Zn under in vitro conditions. The specimens used were initially devoid of metals. Results demonstrated that uptake occurs but only if the cuticle is pretreated with NaOH. Because there was no obvious degradation in the microstructure at the mandible edge, presumably, the treatment did not extract structural elements. Cuticle is comprised of chitin chains and surrounding proteins; however, a harsher protocol (5% NaOH at 100°C) than that used would be needed to remove those proteins attached directly to the chitin fibres (Vincent and Wegst 2004) or cause chitin to undergo significant deacetylation (45% NaOH for 30 min at 15 psi and 121°C; No et al. 2000). It is, therefore, likely that proteins were principally affected.
Uptake of Zn only after cuticle pretreatment with NaOH is likely to occur because any protein present will be in anionic form at the pH of 4% NaOH solution (1 M and pH >12) and likely to exceed the isoelectric point of the protein (Sloane and York 1969). Zinc in acetate solution is an uncomplexed form and will, therefore, bind to any protein activated in anionic form.
What is interesting is that Zn enrichment occurred in the cuticle in a similar pattern to enrichment in the drywood termites (as presented in Cribb et al. 2007). That is, rather than occurring homogeneously throughout the cuticle, the major enrichment occurred only at the edge. This is despite the edge being heavily tanned and, therefore, less prone to cleaving of molecular bonds. The data suggest a specific biochemistry, presumably associated with the protein complex suited to metal-bonding in this area. Schofield (2001) has argued that evidence from arthropods indicates there are two distinct processes, one associated with Zn uptake and a second with Mn and Ca uptake, so an ability to bind one does not necessarily imply an ability to bind the other. Whatever the case, currently, only the drywood termites show the capacity to take up major quantities of Zn into the mandible edge in live individuals, and the presence of this metal in major quantities has been linked to possession of harder mandibles (Cribb et al. 2007).
Cribb et al. (2007) demonstrated that mandibles of termites containing Zn are 20% harder than those containing Mn or no metal. This raised the prospect that mandibles containing Zn may undergo less wear. We applied a scratch test to a number of termite mandibles principally to investigate scratch resistance. Such tests mimic interaction of the mandibles with hard materials in their environment such as during feeding and nest building activities. Wei and Bhushan (2006) highlight that little work has been done in the area of scratch tests for biomaterials and canvas the complexity of comparing materials with differing hardness values. However, it should be noted that in the scratch test, unlike some samples from the work of Wei and Bhushan (2006) which caused difficulties in comparison, there was no evidence of the mandible tissues having been gouged or cut into by the tip during scratching—in fact, there was no evidence of the scratch after the force was removed. Considering termite mandibles containing Zn are harder than those with low levels of Mn or no metals (Cribb et al. 2007), it is not surprising that the smallest values (down to 0.24) for coefficient of friction were found in this Zn-containing group.
It must not be forgotten that hydration level of materials is also significant. Schöberl and Jäger (2006) have shown that more material is removed by wear on wet cuticle than dry cuticle (of the marine worm, Nereis) despite being less hard. This cuticle also has a lower friction as measured by lateral force microscopy (a qualitative assessment) than dry cuticle. Although a larger data set would be beneficial, including one that assesses the use of a range of normal force application and hydration states, the current data obtained for termite mandibles can be interpreted as showing that dry Zn-containing mandibles are likely to be more scratch resistant than mandibles lacking Zn, hence, possibly more resistant to environmental degradation. In contrast, lack of metals or presence of low mass percentage of Mn does not appear to provide an advantage in terms of coefficient of friction. We must continue to look elsewhere for an explanation for why Mn is incorporated into cuticle at these low levels.
References
Abe T (1987) Evolution of life types in termites. In: Kawano S, Connell JH, Hidaka T (eds) Evolution and coadaptation in biotic communities. University of Tokyo Press, Japan, pp 126–148
Abe T, Bignell DE, Higashi M (2000) Termites: evolution, sociality, symbioses, ecology. Kluwer Academic, The Netherlands
Birkedal H, Khan RK, Slack N, Broomell C, Lichtenegger HC, Zok F, Stucky GD, Waite JH (2006) Halogenated veneers: protein cross-linking and halogenation in the jaws of Nereis, a marine polychaete worm. Chem Bio Chem 7:1392–1399
Broomell CC, Khan RK, Moses DN, Michael AM, Pontin G, Stucky GD (2007) Mineral minimization in nature’s alternative teeth. J R Soc Interface 4:19–31
Compère P, Morgan JA, Goffinet G (1993) Ultrastructural location of calcium and magnesium during mineralization of the cuticle of the shore crab, as determined by the K-pyroantimonate method and X-ray microanalysis. Cell Tissue Res 274:567–577
Cribb BW, Stewart A, Huang H, Truss R, Noller B, Rasch R, Zalucki MP (2007) Insect mandibles—comparative mechanical properties and links with metal incorporation. Naturwissenschaften. DOI 10.1007/s00114-007-0288-1
Edwards AJ, Fawke JD, McClements JG, Smith SA, Wyeth P (1993) Correlation of zinc distribution and enhanced hardness in the mandibular cuticle of the leaf-cutting ant Atta sexdens rubropilosa. Cell Biol Int 17:697–698
Eggleton P (2001) Termites and trees: a review of recent advances in termite phylogenetics. Insectes Soc 48:187–193
Eggleton P, Tayashu I (2001) Feeding groups, lifetypes and the global ecology of termites. Ecol Res 16:941–960
Emerson E, Krishna K (1975) The Termite Family Serritermitidae (Isoptera). Am Mus Novit 2570:1–31
Fawke JD, McClements JG, Wyeth P (1997) Cuticular metals: quantification and mapping by complementary techniques. Cell Biol Int 21:675–678
Fontaine AR, Olsen N, Ring RA, Singla CL (1991) Cuticular metal hardening of mouthparts and claws of some forest insects of British Columbia. J Entomol Soc Brit Columbia 88:45–55
Goldstein J, Newbury DE, David C, Joy DC, Lyman CE, Echlin P, Lifshin E, Sawyer LC, Michael JR (2003) Scanning Electron Microscopy and X-ray Micronalysis, 3rd edn. Kluwer Academic, New York
Grube S, Rudolph D (1999) The labial gland reservoirs (Water sacs) in Reticulitermes santonensis (Isoptera: Rhinotermitidae): Studies of the functional aspects during microclimatic moisture regulation and individual water balance. Sociobiology 33:307–323
Hillerton JE, Vincent JFV (1982) The specific location of zinc in insect mandibles. J Exp Biol 101:333–336
Hillerton JE, Reynolds SE, Vincent JFV (1982) On the indentation hardness of insect cuticle. J Exp Biol 96:45–52
Hillerton JE, Robertson B, Vincent JFV (1984) The presence of zinc or manganese as the predominant metal in the mandibles of adult, stored-product beetles. J Stored Prod Res 20:133–137
Inward D, Beccaloni G, Eggleton P (2007) Death of an Order: a comprehensive molecular phylogenetic study confirms that termites are eusocial cockroaches. Biol Lett 3:331–335 [And Suppl. Mat.]
Lichtenegger HC, Schoberl T, Ruokolainen JT, Cross JO, Heald SM, Birkedal H, Waite JH, Stucky GD (2003) Zinc and mechanical prowess in jaws of Nereis, a marine worm. Proc Natl Acad Sci U S A 100:9144–9149
Lo N, Eldridge RH, Lenz M (2006) Phylogeny of Australian Coptotermes (Isoptera: Rhinotermitidae) species inferred from mitochondrial COII sequences. Bull Entomol Res 96:433–437
Lo N, Engel MS, Cameron S, Nalepa CA, Tokuda G, Grimaldi D, Kitade O, Krishna K, Klass K-D, Maekawa K, Miura T, Thompson GJ (2007) Save isoptera: a comment on Inward et al. Biol Lett 3:562–563
Morgan TD, Baker P, Kramer KJ, Basibuyuk HH, Quicke DLJ (2003) Metals in mandibles of stored products insects: do zinc and manganese enhance the ability of larvae to infest seeds. J Stored Prod Res 39:65–75
Newbury DE (1991) Standardless quantitative electron-excited x-ray microanalysis by energy-dispersive spectrometry: what is its proper role. Microsc Microanal 4:585–597
No HK, Cho YI, Kim HR, Meyers SP (2000) Effective deacetylation of chitin under conditions of 15 psi/121 degrees C. J Agric Food Chem 48(6):2625–2627
Ohmura W, Matsunaga H, Yoshimura T, Suzuki Y, Imaseki H (2007) Zinc distribution on the mandible cutting edges of two drywood termites, Incisitermes minor and Cryptotermes domesticus (Isoptera: Kalotermitidae). Sociobiology 50(3):1035–1040
Pearce MJ (1997) Termites biology and pest management. CAB, UK
Quicke DLJ, Wyeth P, Fawke JD, Basibuyuk HH, Vincent JFV (1998) Manganese and zinc in the ovipositors and mandibles of hymenopterous insects. Zool J Linn Soc 124:387–396
Reinhard J, Kaib M (2001) Food exploitation in termites: indication for a general feeding-simulating signal in labial gland secretion of Isoptera. J Chem Ecol 27:189–201
Schöberl T, Jäger IL (2006) Wet or dry—hardness, stiffness and wear resistance of biological materials on the micron scale. Adv Eng Mater 11:1164–1169
Schofield RMS (2001) Metals in cuticular structures. In: Brownell P, Polis G (eds) Scorpion Biology and Research. Oxford University Press, Oxford, pp 234–256
Schofield RMS (2005) Metal-halogen biomaterials. Am Entomol 51:45–47
Schofield RMS, Nesson MH, Richardson KA (2002) Tooth hardness increases with zinc-content in mandibles of young adult leaf-cutter ants. Naturwissenschaften 89:579–583
Schofield RMS, Nesson MH, Richardson KA, Wyeth P (2003) Zinc is incorporated into cuticular “tools” after ecdysis: the time course of the zinc distribution in “tools” and whole bodies of an ant and a scorpion. J Insect Phys 49:31–44
Sloane NH, York JL (1969) Chapter 5. In Review of biochemistry. Macmillan, Toronto, pp 37–42
Thompson GT, Miller LR, Lenz M, Crozier RH (2000) Phylogenetic analysis and trait evolution in Australian lineages of drywood termites (Isoptera, Kalotermitidae). Mol Phylogenet Evol 17:419–429
Vincent JFV, Wegst UGK (2004) Design and mechanical properties of insect cuticle. Arthropod Struct Dev 33:187–199
Wei G, Bhushan B (2006) Nanotribological and nanomechanical characterization of human hair using a nanoscratch technique. Ultramicroscopy 106:742–754
Yoshimura T, Kagemori N, Kawai S, Sera K, Futatsugawa S (2002) Trace elements in termites by PIXE analysis. Nucl Instrum Methods Phys Res B 189:450–453
Yoshimura T, Kagemori N, Sugiyama J, Kawai S, Sera K, Futatsugawa S, Yukawa M, Imazeki H (2005) Elemental analysis of worker mandibles of Coptotermes formosanus (Isoptera: Rhinotermitidae). Sociobiology 45:255–259
Acknowledgements
We gratefully acknowledge supply of termite samples from Prof. Reginaldo Constantino, Universidade de Brasília, Brasil; Mrs. Vivienne Uys, Plant Protection Research Institute, Queenswood, South Africa and Dr. Gerhard Prinsloo, Agricultural Research Council-Plant Protection Research Institute, Pretoria, South Africa; Dr. Michael K. Rust, Department of Entomology, University of California, Riverside, CA, USA; Dr. B. Peters, Queensland Department of Primary Industries and Fisheries; Mr. Greg Daniels, The University of Queensland Insect Collection, Brisbane, Australia; and Mrs. Rachel Hancock Narangbar, Queensland, Australia. We thank Mr. Yueqin Wu, University of Queensland, for his experimental assistance in scratching tests and Dr. Michael Lenz, Commonwealth Scientific and Industrial Research Organisation-Entomology, Canberra, and Dr. David Merritt and Dr. Lyn Cook, University of Queensland for their helpful comments on the manuscript when in preparation. The experiments comply with the current laws of the country in which they were performed.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Cribb, B.W., Stewart, A., Huang, H. et al. Unique zinc mass in mandibles separates drywood termites from other groups of termites. Naturwissenschaften 95, 433–441 (2008). https://doi.org/10.1007/s00114-008-0346-3
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00114-008-0346-3