
Abstract
Fossil footprints are important in understanding Cretaceous avian diversity because they constitute evidence of paleodiversity and paleoecology that is not always apparent from skeletal remains. Early Cretaceous bird tracks have demonstrated the existence of wading birds in East Asia, but some pedal morphotypes, such as zygodactyly, common in modern and earlier Cenozoic birds (Neornithes) were unknown in the Cretaceous. We, herein, discuss the implications of a recently reported, Early Cretaceous (120–110 million years old) trackway of a large, zygodactyl bird from China that predates skeletal evidence of this foot morphology by at least 50 million years and includes the only known fossil zygodactyl footprints. The tracks demonstrate the existence of a Cretaceous bird not currently represented in the body fossil record that occupied a roadrunner (Geococcyx)-like niche, indicating a previously unknown degree of Cretaceous avian morphological and behavioral diversity that presaged later Cenozoic patterns.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Neornithean (modern) birds occupy and have occupied a broad diversity of ecological niches, many of which are facilitated by unusual pedal adaptations, such as zygodactyly and heterodactyly, that are presently unknown in any Mesozoic bird. Until quite recently, no extant bird clade was thought to have arisen until after the end-Cretaceous (K/Pg) extinction event (Feduccia 1995), despite predictions that Cretaceous neornitheans (terminology sensu Harris et al. 2006) must have been present based on phylogenetic and evolutionary projections (Dyke and van Tuinen 2004; Sibley and Ahlquist 1990). Definitive evidence of Mesozoic neornitheans has only recently emerged (Clarke et al. 2005; Kurochkin et al. 2002), but even these are basal (Galloanseres); no undisputed Cretaceous members of the Neoaves (all neornitheans except Galloanseres and Ratitae) are known. All Cretaceous birds for which the pedal skeleton is known are either anisodactyl (digit I reversed with respect to II–IV), the plesiomorphic state for all avians more derived than Archaeopteryx (Mayr et al. 2005, 2007), or pamprodactyl (pedal digit I un-reversed) and, in some cases, possibly totipalmate (all digits conjoined by webbing; Marsh 1880; Mayr et al. 2005). The sole reported exception is the ostensibly heterodactyl enantiornithean Dalingheornis liweii (Zhang et al. 2006), for which evidence of heterodactyly is ambiguous at best: First, the distal end of the second metatarsal in this taxon does not appear to be strongly rotated medially as it is in heterodactyl neornitheans, at least not significantly more so than in other enantiornitheans (e.g., Chiappe 1993, 2002b; Zhou and Zhang 2003a). Second, the ostensibly reversed second digit is not in articulation with the tarsometatarsus, nor are its phalanges articulated; its ungual appears to have been rotated postmortem but pre-burial, during diagenetic compaction of the specimen, or both, a phenomenon occasionally seen in the pedal digits of specimens of some other fossil bird as well [e.g., Sinornis (Sereno et al. 2002, Fig. 8.6)].
Early Cretaceous bird taxa far outnumber coeval avian ichnotaxa particularly in central and eastern Asia, which has been a locus of discovery and study of birds from this epoch (You et al. 2006; Zhou and Zhang 2004; Chiappe 2007). Although the high frequency of Asian fossil bird discoveries is partly attributable to exceptional preservation, the high incidence of bird track discoveries (Kim et al. 2006; Lockley et al. 1992, 2006a,b), in a variety of sedimentary facies, cannot be dismissed as solely a function of preservational bias. Moreover, the most numerous and diverse Early Cretaceous birds are small (crow-sized or smaller) basal pygostylians or members of the Enantiornithes, all taxa of which that are represented by remains that include the foot skeleton are perching birds with anisodactyl feet (Chiappe et al. 1999; Zhou 2004), that are generally inferred to have occupied many of the same niches as extant passeriforms and coraciiforms. Among Early Cretaceous members of the Ornithuromorpha (Chiappe 2002a), the clade that includes the Neornithes and its immediate outgroups, ostensibly basal taxa (Hou et al. 1996) were similarly perchers, but many more derived taxa display evidence of specialization for aquatic/amphibious niches (You et al. 2006; Zhou and Zhang 2006). Some Late Cretaceous ornithuromorphs occupied ground bird niches (Chiappe 2002b). The coeval bird track record is dominated by morphologies reflecting feet similar to those of modern shorebirds (Lockley et al. 1992), while footprints of ground-dwelling taxa are comparatively rare. Thus, the combined body and trace fossil record of Cretaceous birds is cumulative, not duplicative, stressing the often undervalued utility of footprints.
Fossil footprints from the Lower Cretaceous (Aptian–Albian, 120–110 million years old) Tianjialou Formation of Shandong Province, China (Fig. 1), were described and named Shandongornipes muxiai (Li et al. 2005). Although not recognized as such when first described, these tracks are unique because they indicate the presence of an unexpectedly large Early Cretaceous, zygodactyl, cursorial bird that is unknown from body fossils and occupied a niche unknown for any bird of this time period. Shandongornipes is the only zygodactyl bird track type known and is twice as old as the oldest osteological evidence of avian zygodactyly.

Type locality of Shandongornipes muxiai in Shandong Province, People’s Republic of China. a Map of China (white) showing the location (star) of the Tianjialou Formation outcrop that produced the Shandongornipes holotype in Junan County, Shandong Province, near the town of Lingquan. b Footprint-bearing Tianjialou Formation outcrop in Junan. Shandongornipes tracks are circled (dashed line indicates damaged track); non-avian theropod (Th) and ornithopod (Or) tracks are also present on several bedding planes
Materials and methods
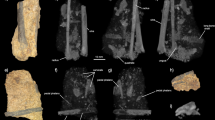
The original description of Shandongornipes (Li et al. 2005), on the basis of natural impressions (negative or concave epireliefs) and plaster replicas (casts or positive, convex hyporeliefs), was made from a single trackway preserved in situ at Houzuoshan Dinosaur Park. The entire trackway configuration was recorded by tracing each track’s outline on acetate film; tracks were enumerated sequentially in the order that they were made (the direction of the track maker’s travel); track 2 was irreparably damaged, and track 3 was partly lost after the replica of it, mentioned below, was made. Track number 4 (Fig. 2a,e,g) was replicated in plaster directly from the rock surface and designated as the holotype [LRH-DH01; abbreviations refer to the Li Rihui collection (LRH) from Houzuoshan Dinosaur Park (DH) reposited in the Qingdao Institute of Marine Geology]; track number 3 (Fig. 2a,c) was replicated the same way and designated as the only paratype (LRH-DH02; see below for more on specimen numbers). Field numbers (LRH dz66–70) were given to all five tracks, although number 2 was almost entirely missing, and numbers 1 and 5 were not replicated.

Tracks of Shandongornipes muxiai. a Map of holotype trackway (LRH-DH01). b–e Photographs and schematics of individual tracks in trackway (digit numbers labeled), including b photograph of first (left foot, LRH-dz70), c outline drawing of third (left foot, LRH-dz68), d outline drawing of fifth (right foot, LRH-dz66), and e photograph of fourth (left foot, LRH-dz67). f Segment of a trackway made by extant roadrunner G. californianus near St. George, Utah; segment includes same left–right sequence preserved in LRH-DH01 (a) and shown at same scale. g Schematic of fourth Shandongornipes print (LRH-dz67). h Individual G. californianus right footprint [from Elbroch and Marks (2001); used with permission] rotated with hallux in approximately same orientation as Shandongornipes print in (g)
In the present study, which involved additional field work (by RL, ML, and MM), tracks 1, 3, 4, and 5 were enumerated sequentially in the order in which they were made, molded in latex, cast in plaster, and reposited in the University of Colorado (CU) at Denver Dinosaur Tracks Museum; the casts are designated as CU 214.116–119, respectively. CU 214.117 and 214.118, respectively, represent copies of the “reverse” plaster casts of LRH dz68 and dz 67 made before the former was damaged. CU 214.122 is a replica of “un-reversed” track 4 (=CU 214.118), and CU 214.123 preserves a post-damage record of track 3. These tracks are reposited with 13 other bird and non-avian dinosaur track replicas (CU 214.103–115) from the same locality. In the present study, all four Shandongornipes tracks, including previously un-replicated tracks 1 and 5, are considered part of the holotype trackway in the emended description in the main text. Track outlines were drawn from a duplicate set of plaster replicas using low angle light and a fine point graphite pencil. These tracings were directly scanned and used as the basis of the revised illustrations (Fig. 2c,d,g). All measurements of footprint lengths and widths and digit divarication angles were made to the nearest 1.0 mm and 1.0° with transparent ruler and protractor and checked on latex molds, replicas, and photocopies.
Description of specimens
Ichnotaxonomy
Shandongornipodidae ichnofam. nov.
Type ichnogenus and ichnospecies
Shandongornipes muxiai Li, Lockley, et Liu 2005.
Description
Tetradactyl, paraxonic, zygodactyl tracks (i.e., with cranially directed digits II and III in opposition to caudally directed digits I and IV). Trackway narrow (about 7.0 cm), only marginally wider than foot width (about 6.0 cm), with long step (about 44 cm) averaging about 5 times foot length (about 8.5 cm).
Holotype and paratype
In the initial description (Li et al. 2005), the holotype of Shandongornipes was referred to as LRH-DH01, but this did not specify precisely what this specimen number entailed. LRH-DH01 is a plaster cast of track number 4 (a right foot) in the trackway of five sequential tracks made by a single individual, which is equivalent to the original track that was given the registration or “field photo number” LRH-dz67. The initial description (Li et al. 2005) also individually numbered each track in the trackway (LRH-dz66 through dz70). Thus, the term “holotype” was used to specify both the replica (LRH-DH01) and the actual uncollected track (LRH-dz67) in the sequence. The same applies to the paratype (LRH-DP02), which corresponds to field number LRH-dz68 (Li et al. 2005; see also “Materials and methods”). The intention of the authors, however, was to specify the entire trackway as the holotype. In tandem, both the cited paratype and the holotype represent left and right tracks (numbers 3 and 4, respectively) in a sequence made by the same individual, and together with the intervening step, they are representative of the entire morphology and gait of the ichnotaxon.
Horizon and locality
Lower Cretaceous Tianjialou Formation (Dasheng Group), DH (near the village of Houzuoshan), near Lingquan, Junan County, Shandong Province, People’s Republic of China (Fig. 1).
Known distribution
Early Cretaceous of eastern Asia.
Revised diagnosis
Tetradactyl, paraxonic, zygodactyl footprints in narrow (6–7 cm wide) trackway with long (41–46 cm) steps. Elongate hallux (digit I) oriented caudally, subparallel to trackway axis, and in opposition to the hypex between cranially directed digits II and III, which subtend a narrow mean angle of about 32°. Digit IV rotated caudolaterally to subtend a mildly acute mean angle of about 77° with the hallux. Phalangeal pad formula 2-3-3 for digits II, III, and IV, respectively (Fig. 2a–e).
Revised description
The only known specimen is a single trackway of five consecutive, tetradactyl, paraxonic, well-preserved, zygodactyl bird tracks (Figs. 1b and 2a). Shandongornipes tracks have average track length (with hallux) of 8.5 cm (range 7.8–8.8 cm) and width (measured perpendicular to trackway axis) of 5.3 cm (range 4.4–6.0 cm). All digit plantar surface traces are separate from one another, indicating that their proximal connection to the metatarsus was typically elevated above the substrate. All digit traces end in narrow, acuminate claw marks. The hallux trace is short, consisting of a single elongate claw mark averaging 1.4 cm long (range 1.0–1.8 cm) and separated proximally from all other digit traces by at least 2.0 cm (Fig. 2b–e). Digit II consists of two proximal pads (mean length 1.1 cm) and a claw trace (mean total length 3.5 cm; range 3.4–3.6 cm) directed craniomedially toward the trackway midline at an acute angle of 10°–15°. Digit III is the longest (mean 4.23 cm; range 4.1–4.4 cm) and comprised of three proximal pads and one distal claw impression; the digit is directed craniolaterally away from the trackway midline at an acute angle of 10°–15°. The length of the digit III impression demonstrates that the track maker was somewhat larger than Sapeornis, the largest Early Cretaceous bird known from osteological remains (Zhou and Zhang 2003a), which has a digit III length of roughly 3.9 cm. Digit IV is caudolaterally directed at a 10°–25° angle caudal to a line perpendicular to the trackway axis; the digit impression comprises three short pads (mean length 0.6 cm) plus a claw trace (mean length 2.95 cm; range 2.7–3.4 cm). The average divarication angles are 152.75° (range 137°–163°) for digits I and II, 32.25° (range 26°–36°) for II and III, 101.25° (range 89°–114°) for III and IV, and 77.0° (range 68°–94°) for I–IV. The average step and stride lengths of the trackway are 44 cm (range 41–46 cm) and 87 cm (range 86–88 cm), respectively. Pace angulation values are high (175°).
Because there are no fossil footprints that bear any close resemblance to Shandongornipes, we erect the new ichnofamily diagnosed aboveFootnote 1. The initial description (Li et al. 2005) interpreted the trackway as if it were an anisodactyl form tending toward heterodactyly and, thus, interpreted digit II as IV. Our detailed analysis and description indicate a zygodactyl, paraxonic form in which the cranially oriented digits II and III are bisected by the parasagittal plane of the trackway axis, and lateral digit IV is easily distinguished by its short pads compared to those of digit II. Most claw traces are widely separated from their associated distal phalangeal pads by distances of up to 1.0 cm, suggesting that the claws were strongly curved such that the main body of each was elevated; therefore, only its plantarily directed claw tip penetrated the substrate.
Shandongornipes is unique among fossil bird trackways in showing almost perfect and highly consistent preservation of all digital pad and claw traces in four out of five consecutive footprints. This may be because, in contrast to most bird tracks of the shorebird variety, its digits were wider, more robust, and thus, more easily registered clear impressions.
The relative lengths of the digit impressions (III longer than IV; both longer than II) are remarkably similar to those of tracks made by the greater roadrunner (Cuculidae: Neomorphinae: Geococcyx californianus), which are also very similar in track size (typical foot lengths 7–9 cm) and step length (reaching 37–48 cm; Elbroch and Marks 2001; Figs. 2f,h and 3e,f). As direct relationship is unlikely—Geococcyx is only known from the Late Pleistocene through Recent and only from North America (Brodkorb 1971; Carpenter and Mead 2003)—it is a remarkable example of convergence in only distantly related and temporally widely separated avian clades.

a–e Polarities in the symmetry and configuration of non-avian theropod and avian pedal digits. All tracks shown are right feet. a, b Polarity between caudalization of median (II and III) hypex (black arrowhead) and lateral (III and IV) hypex (white arrowhead) in the Cretaceous non-avian theropod tracks aSiamopodus (Lockley et al. 2006c) and bSaurexallopus (Lockley et al. 2004). c The nearly symmetrical anisodactyl condition exhibited by Ardea (after Elbroch and Marks 2001). d The heterodactyl condition seen only in trogons (track is hypothetical because no trogon footprints have been described). e Zygodactyly exhibited by Geococcyx (after Elbroch and Marks 2001). Note the strong respective senses of counterclockwise and clockwise rotation that differentiate d and e. f–k Examples of track morphologies made by zygodactyl birds from various extant neoavian clades. All tracks are of left feet shown to same scale (5 cm). fGeococcyx (after Elbroch and Marks 2001). g Parrot (after specimen in CU at Denver collections). hColaptes (after Murie 1954). iAthene (after Jaeger 1948). jPicoides (after Jaeger 1948). kBubo (after Stuart and Stuart 2000)
Shandongornipes is associated with abundant trackways of bipedal non-avian theropod dinosaurs ranging in foot length from about 6.0–30.0 cm, suggesting that its track maker occupied a niche in which carnivorous cursorial bipeds were common and where cursorial ability would have an obvious advantage. Based on the formula:
where v = velocity, g = acceleration due to gravity, SL = stride length, and h = hip height (estimated as 4 × footprint length; Alexander 1976), we derived a speed estimate of 2.23 m/s, or 8.03 km/h, for the Shandongornipes trackway. This likely represents an accelerated pace: The large SL values are typical of higher speed gaits; the alternative explanation, a particularly long hip height for such a short foot length, would be highly unusual and unparsimonious. This supports the conclusion that the track maker was a relatively fast-moving cursorial biped, with cursorial ability, similar to Geococcyx.
Discussion
Shandongornipes tracks were most parsimoniously made by a bird rather than a non-avian theropod. Most prominently, among Mesozoic tetrapods, bipedality is only known in the Dinosauria, particularly in some ornithischians and in theropods. Of these, anisodactyly is seen only within Aves; it is not present in Archaeopteryx (Mayr et al. 2005, 2007), although it is present in some other non-pygostylians such as Jeholornis (Zhou and Zhang 2003b) and Dalianraptor (Gao and Liu 2005). The small overall track size, slenderness of the digit impressions, and wide divarication angles between the outer digits are also characteristic of (though not unique to) bird footprints. Small bird tracks generally exhibit only indistinct pads, however, and, as above, Shandongornipes tracks are unique in this respect.
Shandongornipes tracks were originally assumed to pertain to a shorebird-like taxon (Li et al. 2005), but the tracks are unique in possessing impressions of not only the plesiomorphically reversed hallux but an additional reversed or highly divergent digit IV as well. Among extant cursorial birds, zygodactyly is best known in the greater roadrunner (Engels 1938), but evolved convergently to varying degrees in numerous arboreal, exclusively neoavian taxa, including parrots (Psittaciformes; Fig. 3g), woodpeckers (Picidae; Fig. 3h), and owls (Strigiformes; Fig. 3i; see also Supplementary table 1 for a list of zygodactyl neoavians and their fossil records). Geococcyx itself plesiomorphically retains zygodactyly from its cuculid ancestors. Shandongornipes thus could have been made by a Cretaceous bird occupying a Geococcyx-like niche. Given the predominant association of zygodactyly with arboreality among neornitheans (Fig. 3f–i; see also Supplementary table 1), a cursorial owl-, parrot-, or woodpecker-like Shandongornipes track maker with unusually long steps appears unlikely. The holotype trackway forms a component of a larger ichnofauna that includes ornithopod, deinonychosaur, and other non-avian theropod tracks (Fig. 1b). Deinonychosaur tracks in particular are extremely rare in the palichnological record. The Tianjialou Formation thus appears to preserve an unusual faunal record, possibly a paleoenvironment typically frequented during the Early Cretaceous by both deinonychosaurs and large, cursorial ground birds that is not represented at most other track-bearing sites.
Two pedal morphologies could potentially produce tracks with two reversed digits: zygodactyly (digit IV reversed) and heterodactyly (digit II reversed), both of which are perching/grasping adaptations that presumably evolved from a plesiomorphic anisodactyl morphology (Bock and Miller 1959). Indeed, heterodactyl and zygodactyl foot structures are only distinguishable via the morphology of the tarsometatarsus. Two important factors argue in favor of a zygodactyl track maker for Shandongornipes. First, heterodactyly is rare, known exclusively in the Trogoniformes, for which direct fossil evidence of a heterodactyl pes dates back to the Middle Eocene (Mayr 2005), although a trogon is represented by isolated cranial material from the Latest Paleocene–Earliest Eocene of Denmark (Kristoffersen 2002), and a heterodactyl tarsometatarsus of uncertain affiliations is known from the Lower Eocene London Clay (Mayr 1999). Extant trogons are arboreal and have small, comparatively weak legs that are poorly suited to a terrestrial lifestyle (Collar 2001). Obligate and facultative zygodactyly, in contrast, are relatively common in extant birds, having evolved convergently numerous times (Gill 1995; Fig. 3f–i; see also Supplementary table 1). Second, the digital pad formula in Shandongornipes is well preserved and suggests a 2-3-3 arthral formula for the plantar portions of digits II, III, and IV, respectively, with proximodistally long phalangeal pads in digit II and short pads in digit IV. This conforms closely to the most common avian pattern (2-3-4), although it is known that digital pads can be both arthral and mesarthral and, thus, do not necessarily reflect the underlying phalangeal formulae in footprints (Olsen et al. 1998). Thus, unparsimonious assumptions are required to assume that Shandongornipes represents a heterodactyl foot.
The holotype trackway was initially interpreted in reverse (Li et al. 2005), as if it were an exaggerated anisodactyl track tending toward heterodactyly. However, Shandongornipes prints exhibit zygodactyly, with a tendency toward ectropodactyly. The latter, a trunk-climbing adaptation that typifies most modern woodpeckers (Picidae; Bock and Miller 1959), is characterized both by rotation of digit IV to a position roughly 90° to digits II and III and decreased retroversion of digit I. Shandongornipes prints retain fully reversed halluces, however, and thus cannot be considered truly ectropodactyl; they also lack any indication of the conjoined bases of digits II and III that are common among picids.
Developmental derivation of the zygodactyl from the anisodactyl condition requires digit IV to rotate caudolaterally, increasing the hypex between digits III and IV and decreasing its counterpart between I and IV. Conversely, heterodactyly derives from caudomedial rotation of digit II, increasing the II and III and reducing the I and II hypexes. In either case, the parasagittal orientation of digits I and III remain relatively fixed. In the former, the lateral hypex (III and IV) migrates caudally relative to the medial hypex (II and III), and vice versa in the latter case. Both hypex orientation polarities exist in non-avian theropod tracks (Fig. 3; Lockley 1998), but tendency toward ectropodactyly/zygodactyly (i.e., caudalization of hypex III and IV) is far more common than toward heterodactyly, foreshadowing the situation in extant birds.
Shandongornipes prints demonstrate variability in the degree of zygodactyly (Fig. 2b–e). The most pronounced retroversion of digit IV is in tracks 3 and 5 (left footprints), whereas left track 1 and especially right track 4 exhibit more subtly “ectropodactyl” configurations. This may reflect facultative, rather than obligate, zygodactyly in the track maker. Shandongornipes prints do not exhibit the extreme degree of zygodactyly seen in many extant bird feet, but the degree of zygodactyly varies markedly in the latter, as well (Fig. 3f–i). Variations in the rotation, digit divarication, or step lengths (limping) between consecutive left and right footprints in trackway sequences are known elsewhere in the track record (Lockley et al. 1994) and do not necessarily indicate an unusual or pathological condition in the Shandongornipes track maker.
Zygodactyl (and heterodactyl) footprints are completely unknown in the fossil record. Of the neornithean taxa that are characterized in part by zygodactyl feet, the fossil records of some extend back to the Early or Middle Eocene; others extend to the Paleocene (see also Supplementary table 1), rendering skeletal evidence of zygodactyly a maximum of 65–56 million years old (Gradstein et al. 2004). Shandongornipes, therefore, effectively doubles the known temporal range of this morphology and represents an avian morphology and paleoecology unknown from any body fossil and, hitherto, unknown before the Cenozoic. The Shandongornipes track maker appears to have been a cursorial bird with a long step, functionally convergent with the extant roadrunner Geococcyx (Fig. 2f). The phylogenetic relationships of the Shandongornipes track maker cannot be established on the basis of tracks alone, but the restriction of zygodactyl neoavian clades to the Cenozoic render a neornithean track maker highly unlikely. Shandongornipes, thus, represents a striking example of evolutionary convergence of an Early Cretaceous bird with the locomotory habits and possibly the ecology of the modern roadrunner.
Notes
Unlike phylogenetic systematics but like other parataxonomic systems, ichnotaxonomy, by convention, does not parallel evolutionary phylogeny, utilizing instead artificial groupings that represent particular morphologies and/or behavioral modes of, phylogenetically, often disparate organisms—widely disparate organisms can produce similar ichnites that indicate similar ecologies and/or behaviors. Thus, an ichnofamily is not a Linnean entity per se, and ichnofamilies can be monotaxic because they are intended only to indicate a high degree of difference from other ichnological morphologies and/or behaviors. Given the multiple occurrences of zygodactyly in birds and, indeed, some other tetrapods, other fossil zygodactyl tracks should eventually be found and also reside in this ichnofamily. See Bertling et al. (2006) and Lockley (2007) for further details.
References
Alexander RM (1976) Estimates of the speeds of dinosaurs. Nature 261:129–130
Bertling M, Braddy SJ, Bromley RG, Demathieu GR, Genise J, Mikuláš R, Nielsen JK, Nielsen KSS, Rindsberg AK, Schlirf M, Uchman A (2006) Names for trace fossils: a uniform approach. Lethaia 39:265–286
Bock WJ, Miller WD (1959) The scansorial foot of the woodpeckers, with comments on the evolution of perching and climbing feet in birds. Am Mus Novit 1931:1–45
Brodkorb P (1971) Catalogue of fossil birds: part 4 (Columbiformes through Piciformes). Bull Fla State Mus Biol Sci 15:163–266
Carpenter MC, Mead JI (2003) Late Pleistocene roadrunner (Geococcyx) from Kartchner Caverns State Park, southeastern Arizona. Southwest Nat 48:402–410
Chiappe LM (1993) Enantiornithine (Aves) tarsometatarsi from the Cretaceous Lecho Formation of northwestern Argentina. Am Mus Novit 3083:1–27
Chiappe LM (2002a) Basal bird phylogeny: problems and solutions. In: Chiappe LM, Witmer LM (eds) Mesozoic birds: above the heads of dinosaurs. University of California Press, Berkeley, pp 448–472
Chiappe LM (2002b) Osteology of the flightless Patagopteryx deferrariisi from the Late Cretaceous of Patagonia (Argentina). In: Chiappe LM, Witmer LM (eds) Mesozoic birds: above the heads of dinosaurs. University of California Press, Berkeley, pp 281–316
Chiappe LM (2007) Glorified dinosaurs: the origin and early evolution of birds. Wiley-Liss, Hoboken
Chiappe LM, Ji S, Ji Q, Norell MA (1999) Anatomy and systematics of the Confuciusornithidae (Theropoda: Aves) from the Late Mesozoic of northeastern China. Bull Am Mus Nat Hist 242:1–89
Clarke JA, Tambussi CP, Noriega JI, Erickson GM, Ketcham RA (2005) Definitive fossil evidence for the extant avian radiation in the Cretaceous. Nature 433:305–309
Collar NJ (2001) Family Trogonidae (Trogons). In: Del Hoyo J, Elliott A, Sargatal J (eds). Handbook of the birds of the world, vol 6. Lynx Edicions, Barcelona, pp 80–127
Dyke GJ, van Tuinen M (2004) The evolutionary radiation of modern birds (Neornithes): reconciling molecules, morphology and the fossil record. Zool J Linn Soc 141:153–177
Elbroch M, Marks E (2001) Bird tracks & sign: a guide to North American species. Stackpole books, Mechanicsburg
Engels WL (1938) Cursorial adaptations in birds. Limb proportions in the skeleton of Geococcyx. J Morphol 63:207–217
Feduccia A (1995) Explosive evolution in Tertiary birds and mammals. Science 267:637–638
Gao C-L, Liu J-Y (2005) A new avian taxon from Lower Cretaceous Jiufotang formation of western Liaoning. Global Geology 24:313–318
Gill FB (1995) Ornithology, 2nd edn. Freeman, New York
Gradstein FM, Ogg JG, Smith A (2004) A geologic time scale 2004. Cambridge University Press, Cambridge
Harris JD, Lamanna MC, You H-L, Ji S-A, Ji Q (2006) A second enantiornithean (Aves: Ornithothoraces) wing from the Early Cretaceous Xiagou Formation near Changma, Gansu Province, People’s Republic of China. Can J Earth Sci 43:547–554
Hou L, Martin LD, Zhou Z, Feduccia A (1996) Early adaptive radiation of birds: evidence from fossils from northeastern China. Science 274:1164–1167
Jaeger E (1948) Tracks and trailcraft. Macmillan, New York
Kim JY, Kim SH, Kim KS, Lockley M (2006) The oldest record of webbed bird and pterosaur tracks from South Korea (Cretaceous Haman Formation, Changseon and Sinsu islands): more evidence of high avian diversity in East Asia. Cretac Res 27:56–69
Kristoffersen AV (2002) An Early Paleocene Trogon (Aves: Trogoniformes) from the Fur Formation, Denmark. J Vertebr Paleontol 22:661–666
Kurochkin EN, Dyke GJ, Karhu AA (2002) A new presbyornithid bird (Aves, Anseriformes) from the Late Cretaceous of Southern Mongolia. Am Mus Novit 3386:1–11
Li R, Lockley MG, Liu M (2005) A new ichnotaxon of fossil bird track from the Early Cretaceous Tianjialou Formation (Barremian–Albian), Shandong Province, China. Chin Sci Bull 50:1149–1154
Lockley M (1998) Philosophical perspectives on theropod track morphology: blending qualities and quantities in the science of ichnology. Gaia 15:279–300
Lockley MG (2007) A tale of two ichnologies: the different goals and potentials of invertebrate and vertebrate (tetrapod) ichnotaxonomy and how they relate to ichnofacies analysis. Ichnos 14:39–57
Lockley MG, Yang SY, Matsukawa M, Fleming F, Lim SK (1992) The track record of Mesozoic birds: evidence and implications. Philos Trans R Soc Lond B 336:113–134
Lockley MG, Hunt AP, Moratalla JJ, Matsukawa M (1994) Limping dinosaurs? Trackway evidence for abnormal gaits. Ichnos 3:193–202
Lockley MG, Nadon G, Currie PJ (2004) A diverse dinosaur-bird footprint assemblage from the Lance Formation, Upper Cretaceous, eastern Wyoming: implications for ichnotaxonomy. Ichnos 11:229–249
Lockley M, Matsukawa M, Ohira H, Li J, Wright J, White D, Chen P (2006a) Bird tracks from Liaoning province, China: new insights into avian evolution during the Jurassic–Cretaceous transition. Cretac Res 27:33–43
Lockley MG, Houck K, Yang S-Y, Matsukawa M, Lim S-K (2006b) Dinosaur-dominated footprint assemblages from the Cretaceous Jindong Formation, Hallyo Haesang National Park area, Goseong County, South Korea: evidence and implications. Cretac Res 27:70–101
Lockley MG, Matsukawa M, Sato Y, Polahan M, Daorerk V (2006c) A distinctive new theropod dinosaur track from the Cretaceous of Thailand: implications for theropod track diversity. Cretac Res 27:139–145
Marsh OC (1880) Odontornithes: a monograph on the extinct toothed birds of North America. Prof Pap Eng Dept US Army 18:1–201
Mayr G (1999) A new trogon from the Middle Oligocene of Céreste, France. Auk 116:427–434
Mayr G (2005) New trogons from the Early Tertiary of Germany. Ibis 147:512–518
Mayr G, Pohl B, Peters DS (2005) A well-preserved Archaeopteryx specimen with theropod features. Science 310:1483–1486
Mayr G, Pohl B, Hartman S, Peters DS (2007) The tenth skeletal specimen of Archaeopteryx. Zool J Linn Soc 149:97–116
Murie OJ (1954) A field guide to animal tracks. Houghton-Mifflin, Boston
Olsen PE, Smith JB, McDonald NG (1998) Type material of the type species of the classic theropod footprint genera Eubrontes, Anchisauripus, and Grallator (Early Jurassic, Hartford and Deerfield basins, Connecticut and Massachusetts, U.S.A.). J Vertebr Paleontol 18:586–601
Sereno PC, Rao C, Li J (2002) Sinornis santensis (Aves: Enantiornithes) from the Early Cretaceous of northeastern China. In: Chiappe LM, Witmer LM (eds) Mesozoic birds: above the heads of dinosaurs. University of California Press, Berkeley, pp 184–208
Sibley CG, Ahlquist JE (1990) Phylogeny and classification of birds: A study in molecular evolution. Yale University Press, New Haven
Stuart C, Stuart T (2000) A field guide to the tracks and signs of southern and eastern African wildlife. Struik/New Holland Publishers, Cape Town
You H-L, Lamanna MC, Harris JD, Chiappe LM, O’Connor J, Ji S-A, Lü J-C, Yuan C-X, Li D-Q, Zhang X, Lacovara KJ, Dodson P, Ji Q (2006) A nearly modern amphibious bird from the Early Cretaceous of northwestern China. Science 312:1640–1643
Zhang Z, Hou L, Hasegawa Y, O’Connor J, Martin LD, Chiappe LM (2006) The first Mesozoic heterodactyl bird from China. Acta Geol Sin 80:631–635 (engl ed)
Zhou Z (2004) The origin and early evolution of birds: discoveries, disputes, and perspectives from fossil evidence. Naturwissenschaften 91:455–471
Zhou Z, Zhang F (2003a) Anatomy of the primitive bird Sapeornis chaoyangensis from the Early Cretaceous of Liaoning, China. Can J Earth Sci 40:731–747
Zhou Z, Zhang F (2003b) Jeholornis compared to Archaeopteryx, with a new understanding of the earliest avian evolution. Naturwissenschaften 90:220–225
Zhou Z-H, Zhang F-C (2004) Mesozoic birds of China: an introduction and review. Acta Zool Sin 50:913–920
Zhou Z, Zhang F (2006) Discovery of an ornithurine bird and its implication for Early Cretaceous avian radiation. Proc Natl Acad Sci USA 102:18998–19002
Acknowledgments
Earlier drafts of this manuscript were greatly improved by discussions with and comments and reviews by L. Chiappe, M. Lamanna, G. Mayr, A. R. C. Milner, J. O’Connor, H.-L. You, and an anonymous reviewer. This study was supported financially, in part, by National Natural Science Foundation of China grant 40572011 to RL. Additional support was provided by the University and Society Collaboration of the Japanese Ministry of Education, Science, Sports and Culture grant-in-aid 1183303, 1999–2001 to M. M. and by the CU at Denver Dinosaur Tracks Museum to ML. The authors declare that all material was collected, examined, and replicated in full compliance with the laws of their countries.
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by G. Mayr
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
About this article
Cite this article
Lockley, M.G., Li, R., Harris, J.D. et al. Earliest zygodactyl bird feet: evidence from Early Cretaceous roadrunner-like tracks. Naturwissenschaften 94, 657–665 (2007). https://doi.org/10.1007/s00114-007-0239-x
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00114-007-0239-x