
Abstract
The aim of the paper is to review critically the notion of autopoiesis as presented by Maturana and Varela. In particular, recognizing that there are difficulties in obtaining a complete and clear picture from the primary literature, an effort is made to present a coherent view—also based on many years of personal contact with Francisco Varela. The paper begins with a few historical notes to highlight the cultural background from which the notion of autopoiesis arose. The basic principles of autopoiesis as a theory of cellular life are then described, emphasizing also what autopoiesis is not: not an abstract theory, not a concept of artificial life, not a theory about the origin of life—but rather a pragmatic blueprint of life based on cellular life. It shown how this view leads to a conceptually clear definition of minimal life and to a logical link with related notions, such as self-organization, emergence, biological autonomy, auto-referentiality, and interactions with the environment. The perturbations brought about by the environment are seen as changes selected and triggered by the inner organization of the living. These selective coupling interactions impart meaning to the minimal life and are thus defined by Maturana and Varela with the arguable term of "cognition". This particular view on the mutual interactions between living organism and environment leads these authors to the notion of "enaction", and to the surprising view that autopoiesis and cognition are two complementary, and in a way equivalent, aspects of life. It is then shown how cognition, so defined, permits us to build a bridge between biology and cognitive science. Autopoiesis also allows one to conceive chemical models of minimal cellular life that can be implemented experimentally. The corresponding work on "chemical autopoiesis" is then reviewed. The surprising impact of autopoiesis in the social sciences ("social autopoiesis") is also briefly discussed. This review also comments on why the theory of autopoiesis had, and still has, a difficult time being accepted into the mainstream of life-science research. Finally, it is pointed out that the new interest in system biology and complexity theories may lead to a reappraisal of autopoiesis and related notions, as outlined also by other authors, such as Tibor Ganti and Stuart Kauffmann.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
The background
The aim of this article is to review the notion of autopoiesis as developed by Maturana and Varela (Varela et al. 1974; Maturana and Varela 1980; Varela and Maturana 1998) and to present it in the context of present-day research in life science. This work is prompted mainly by two observations. The first is that this theory is not always well understood—also due to the shortcomings of the primary literature; the second is that: there is a shift of the "Zeitgeist" of life science towards system biology, which is a view that emphasizes the complexity of the whole system seen as a self-organized unity. In this partly new scenario, the cellular theories, such as autopoiesis, are once again attracting attention, in particular due to the work being developed in the field of experimental cellular models (Szostak et al. 2001; Deamer et al. 2003; Luisi et al. 2002; Pohorille and Deamer 2002).
Autopoiesis deals with the question "what is life?" and attempts to define, beyond the diversity of all living organisms, a common denominator that allows for the discrimination of the living from the non-living. Autopoiesis is not concerned with the origin of life per se, namely with the transition from the non-living to the living; nor is it primarily interested in artificial life. Rather, it is based on the direct observation of how extant cells work, and is concerned with the various processes connected with life, such as the interaction with the environment, evolution, and "cognition".
Autopoiesis is embedded in a particular cultural background and therefore some historical information is necessary at the start. The seeds of the notion of autopoiesis were sown from the first contacts in the 1960s between the young Francisco Varela and his mentor, Humberto Maturana, at the University of Chile in Santiago. Maturana was already famous for his work on visual perception in frogs (Maturana et al. 1960). This was actually the basis for his later work with Varela against representationalism in perception (see also Varela and Maturana 1998).
Varela left Chile in 1968 to work toward a Ph.D. in biology in Harvard, where he had the opportunity to develop his interest in philosophy. European authors such as Husserl, Heidegger and Merleau-Ponty were particularly important for his work. It was also the time when Piaget's famous "Biologie et Connaissance" (Piaget 1969) appeared and was read by Varela with great attention. At Harvard, Varela was exposed to the field of cybernetics and artificial life through his work and friendship with Heinz von Förster. Varela went back to the University of Chile in 1970. There his friendship and collaboration with Maturana deepened. The biological basis of cognition had always been an important item on Maturana's agenda, and during this period he had developed his important theory on the neurophysiology of cognition (Maturana 1970).
The term autopoiesis was used for the first time by the Santiago authors in 1971; by the end of that year Maturana and Varela had prepared a very long manuscript entitled "Autopoiesis: the organization of the living systems". Autopoiesis, from the Greek, means self-producing.
As Varela (2000) recounts, the manuscript was not well received. It was rejected by the most important journals, and colleagues' response was lukewarm.
This was also a difficult time for F. Varela due to the political situation in Chile: President Allende was assassinated and Varela, as one of his supporters, lost his job and had to leave the country in 1973.
Finally a paper on autopoiesis was submitted and later published for the first time in English (Varela et al. 1974).
The notion of autopoiesis was very slow to become recognized. Eventually, some authors begun to see autopoiesis as part of the emergent field of self-organization (Zeleny 1977) and by the mid-1970s international meetings had used the term in their program; books on autopoiesis by other authors appeared (Zeleny 1977; Benseler et al. 1980) and eminent biologists such as Lynn Margulis accepted it as an integral part of the description of the living (Margulis and Sagan 1995). The term "social autopoiesis" was coined (Luhmann 1984; Teubner 1993; Paulson 1988; Mingers 1992, 1995, 1997) and in fact the impact of autopoiesis in the social sciences came as a surprise to the authors. In the 1990s, experimental chemical systems were developed in Zürich based on the theory of autopoiesis (Bachmann et al. 1992; Luisi 1993).
However, it cannot be said that the notion of autopoiesis is now familiar in mainstream science. The term is neither well known or frequently cited, and I will discuss why this is so later on, but we can anticipate that this is partly due to the fact that autopoiesis theory is not centered on DNA, RNA and on replication, and makes only a minimal use of the term "information". Furthermore, the fact that it had been used extensively in the social sciences, and not always in a very rigorous way, gave some people the impression that the concept might be unscientific, even "new-agey".
There are now, however, timely signs that the interest in autopoiesis is growing, and this article would like to offer a contribution to this new development.
Basic autopoiesis
The autopoietic analysis of the living is based on cellular life, the main argument for this being simply that there are no other forms of life on Earth. We all know that even the simplest cells on Earth are extremely complex, encompassing hundreds of genes and other macromolecules. However, beyond this complexity, the question of what a cell really does lends itself to a simple answer. Consider the diagram of Fig. 1, which schematizes the function of a cell. The first thing one observes is the boundary—a semipermeable spherically closed membrane that separates the cell from the medium. Here the term semipermeable means that certain substances (nutrients and some chemicals) are able to penetrate inside, whereas most other chemicals cannot.

Schematization of the action of a cell. One important feature is the boundary, which is created by the internal network of reactions (a boundary of its own making). The network of reactions brings about a large series of transformations; however, under homeostatic conditions all material that disappears is generated again by the internal machinery. Thus, the cell (and by inference the living) can be seen as a factory concerned with self-maintenance
The notion of boundary is, in fact, central in the theory of autopoiesis. Inside the boundary of a cell, many reactions and, correspondingly, many chemical transformations occur. However, despite all these chemical processes, the cell always remain itself, it maintains its own identity. This is so because the cell (under steady state conditions and/or homeostasis), re-generates within its own boundary all those chemicals that are being destroyed or transformed, be they ATP, glucose, amino acids, or proteins. The chain of processes occurring inside the boundary serves essentially the purpose of self-sustainability, or auto-maintenance. Of course, this takes place at the expense of nutrients and energy coming from the medium: the cell is a dissipative, open system. This last point is also made and discussed by Capra, who generally gives a good account of autopoiesis in his latest book (Capra 2002).
From these simple, basic observations, Maturana and Varela (this way of thinking is often referred to as the "Santiago school") arrived at a generalization of living systems based on the autopoietic unit: an autopoietic unit is a system that is capable of self-sustaining owing to an inner network of reactions that re-generate all the system's components (Varela et al. 1974; Varela 1979, 2000; Maturana and Varela 1980; Varela and Maturana 1998).
In other words, an autopoietic system organizes the production of its own components, so that these components are continuously re-generated and the system can therefore maintain the very network process that produces them. The organization of the living, basically, appears then as the constitutive mechanism of the constitution of the identity as a material entity.
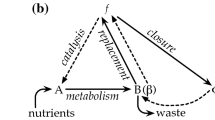
In this way, autopoiesis is capable of capturing the mechanism that generates the essence of the living. The components organize themselves (auto-organization) in a bounded system that produces the components that in turn produce the system, and so on. A graphic representation is given in Fig. 2. Accordingly, the blueprint of life obeys a circular logic- without an identified beginning and/or end. Although the system, as we have already mentioned, is open from the physical point of view, it is one that from an epistemological perspective has a logical operational closure (Varela 1979, 2000). This operational closure characterizes the system as an autonomous identity that can be defined as auto-referential: it produces its own rules of existence and therefore has a particular type of "bio-logical" coherence.

The cyclic logic of cellular life. In the cell, or in the autopoietic unit—which is equivalent—the boundary determines a network of reactions that in turn produces the molecular components that assemble into the organized system that determines the reaction network ... and so on ... with no beginning nor end
It must be re-emphasized that autopoiesis is not an abstract theory of life within the framework of artificial life. This misconception occasionally appears in the literature (Boden 2000), perhaps also due to the fact that Varela and coworkers did occasionally indulge in computer simulation of autopoiesis (Varela et al. 1974; McMullin and Varela 1997). On the other hand, there is in Varela (although not consistently in all primary references) a strong emphasis that the boundary is a physical space. Again, the biological cell is the primary source of inspiration for autopoiesis.
Criteria of autopoiesis and the definition of life.
The most general property of an autopoietic system is the capability of generating its own components via a network process that is internal to the boundary. The boundary of the system must be "of its own making", i.e., also a product of the process of component production. Whether a given system is capable of making its own boundary or not is often the most discriminating criterion for recognizing whether a given system is autopoietic or not (allopoietic). Thus, a virus is not an autopoietic system, as it does not produce its protein coat or the nucleic acids from within its boundary (the host cell does it, and it is living). The question of the criteria of autopoiesis is formalized at length, and not always in a clear way, in the primary literature of autopoiesis. Varela, in his latest book (Varela 2000), has simplified these criteria to three basic ones, which can be expressed in the following way: verifying (1) whether the system has a semipermeable boundary that (2) is produced from within the system and (3) that encompasses reactions that re-generate the components of the system.
Based on this, one is able discriminate the living from the non-living. Consider for example Table 1, which has been presented and discussed earlier by Luisi et al. (1996). All components in the list on the left obey the criteria of autopoiesis, whereas none of components in the list on the right obey them.
It appears from Table 1 then that what we have said about the behavior of a cell is also valid for a mammal or a tree: in each case there is a boundary that is produced from the inside of the living organism, and the internal network of reactions re-generates the organism's components. The living is a factory that makes itself from within. This is the common denominator of the living—regardless of whether we are looking at a microorganism or an elephant. It must be said at this point that in its original form, autopoiesis was limited to cellular life. Varela, for example, was for a long time opposed to generalizing it. In fact, it took quite a while before he could publicly accept (Luisi et al. 1996) that the criteria of autopoiesis could be applied to all higher forms of life, man included.
Once this is accepted, it is clear that the definition of autopoietic unit given above corresponds to a definition of the living: A system can be said to be living when it is defined by a semipermeable chemical boundary which encompasses a reaction network that is capable of self-maintenance by a process of self-generation of the system's components from within. This definition of life has been compared with other possible definitions (see also Luisi 1998).
This definition of life is also, of course, controversial, as particularly in this era of science, life is more popularly defined according to the RNA world-view (Joyce 1994): namely, as a system in which the main ingredients are self-replication and Darwinian evolution in terms of nucleic-acid mechanisms. The difference between the two views is interesting. For example, a colony of bacteria that lives and vegetate without reproducing itself would be alive according to autopoiesis, but not according to the neo-Darwinian view. Also, note—as already emphasized (Luisi 1997, 1998) that the Darwinian view can define life only in terms of a population, whereas autopoiesis also refers to single individuals. In this sense, autopoietic criteria can be applied to laboratory synthetic systems deprived of a genetic heritage; and also—as it does not expressly mention DNA in its basic definitions—to non-coded life.
Maturana and Varela are not the only authors, of course, who emphasize the cellular view for life's basic mechanisms. It is appropriate to recall the work of Morowitz (Morowitz 1992; Morowitz et al. 1996) and, more recently, the importance of metabolism has been emphasized by Boden (2000). One should also mention the work of Tibor Ganti. This Hungarian author presented, in the early 1970s, at about the same time as Maturana and Varela, his view on the "chemoton". Looking at the work of a cell, he sees in it three subsystems, i.e., the cytoplasm, the genetic material, and the cell membrane. For each of these three subsystems, Ganti (1975) develops a model and a set of formal equations; the chemoton being the operative sum of these three subsystems. Also in this case the self-organization of the cellular network is the main factor for cellular life (although Ganti does not use this terminology) as an emergent, distributed property. He is more detailed in the chemical rendering of the cellular life than Varela and Maturana; for example, he develops the notion of autocatalytic cycles and gives biochemical examples (Ganti 1975).
Stuart Kauffmann has introduced over several years (Kauffman 1995) the notion of autocatalytic networks, suggesting that this can very well correspond to the origin of life. Later on (Kauffman 2000) he introduced the notion of "autonomous agent" to describe a life cycle: the novelty here is in the introduction of the thermodynamic notion of work, in the form of a Carnot cycle, within the life cycle of an autonomous agent.
It would be interesting to delve deeper into the comparison between these three views (Maturana and Varela; Ganti, and Kauffman), as these authors appear to share a common view about minimal life as a distributed, emergent property based on an organized network of reactions and/or processes. This comparison (which cannot be carried out here) would also be useful because the three groups of authors—except for a fleeting citation—do not seem to be very well informed about each other's work.
Autopoiesis, emergence, and biological autonomy
Minimal life as defined by autopoiesis is a particular form of self-organization, as visualized in Fig. 2. The relationship between autopoiesis, emergence, and biological autonomy is rather complex (Varela and Maturana 1998; Varela 2000). In order to give a glimpse into this, let us first recall that emergent properties are those novel properties that arise when components (parts) assemble themselves into a higher structural complexity. They are those properties that are not present in the components themselves and arise only when their collective, novel, high-complexity structure is formed (for a general review see Luisi 2002). The autopoietic cycle of Fig. 2 thus suggests an example of emergence, as the new properties of the bounded structure arise only when the components are assembled together. Actually, life itself can be seen as an emergent property, something that the single, non-living components do not display, and that occurs only at the level of the organized, distributed ensemble. The autopoietic organization is characterized by the fact that the product of this organization is the living system itself and that there is no separation between "producer" and "product" (Varela and Maturana 1998).
The emergence of life, as Varela repeats, is a very special novel emergent property: with life, an autopoietic unit acquires the singular property of becoming a biologically autonomous system, namely one that is capable of specifying its own rules of behavior. According to the authors then, autopoiesis is the mechanism that imparts autonomy to the living.
Varela repeats that, from the epistemological point of view, the notion of biological autonomy is equivalent to the notion of auto-referentiality (Varela 1979, 2000) and, in turn, auto-referentiality is connected to the concept of operational closure. This is a process of circular and reflexive interdependency, whose primary effect is its own production. Operational closure must not be viewed as a lack of contact with the environment—we will see in a moment that in the autopoietic perspective such contact is, in fact, central.
From all the above it is apparent that autopoiesis belongs epistemologically to systems theory, according to which it is the organization of the components that characterizes the quality of the system. Thus, the life of a cell is a global property, and cannot be ascribed to any single component.
Actually, the living cell as an autonomous system can be seen as a "self without localization" (Varela and Maturana 1998; Varela 2000). Varela also emphasizes the mutual causality between the local rules of interaction and the global properties of the entire cellular entity (Varela 2000). "Such a reciprocal causality permits a very productive new way of looking at the identification of [the] auto-organization such that the global and the local are correlated to each other...". Autopoiesis is a particular mechanism of auto-organization that permits "the elimination of the traditional opposition between the constituent components and the global properties".
What autopoiesis does not include
The reason why DNA is not mentioned in the autopoietic definition of life lies in the fact that the theory of autopoiesis is primarily concerned with the internal logic (the general "bio-logical" aspects) of minimal life. This should not be understood as the demise of the importance of nucleic acids—which would be absurd—and in fact DNA and RNA are considered as central units in some of the other work of Maturana and Varela (see, for example, Maturana and Varela 1980). However, within the theory of autopoiesis, nucleic acids are only seen as agents that participate in the cell's self-production. Varela and Maturana often emphasize that before one can talk about properties of the living, one has to have the place to put them: the container and the logistics must be there first. Likewise, in describing a car, before talking about the nature of fuel, one has to have a logical scheme describing how a car works and how motion is transmitted from the carburetor to the wheels.
By the same token, the autopoietic definition of life does not mention reproduction. Varela states that to include reproduction in the definition of the living would be ontologically wrong (Varela 2000), as "reproduction is a ...consequence of the existence of individuals. The difficult thing is to create an organism that is capable to self-reproduce with its own boundary. To divide it up in two is easy...". And again (Varela and Maturana 1998), "In order to reproduce something, the unit must first be constituted as a unit, with an organization that defines this unit itself. This is simple common sense logic"; adding that, "A living organism can also exist without being capable of self-reproduction...".
Obviously, this kind of statement is generally arguable and, furthermore, the falling of this statement into a field dominated by the RNA world-view and by the equivalence between DNA and life, may explain why autopoiesis did not gain much ground at the start—and also later on. This problem may have been at least partly avoided, had the authors been less rigid on the matter. In fact, it is not difficult to incorporate nucleic acids and enzymes into the autopoietic scheme of Fig. 1. This was proposed later on (Luisi 1993; Luisi et al. 1996) and the corresponding modification is rather simple, as Fig. 3 shows.

The autopoietic cycle extended to the DNA/RNA/protein world. This is the autopoietic representation of the coded life
The notion that in an autopoietic system reproduction is a consequence of the internal logic of the living, can be visualized in Fig. 4 (Luisi 1993), which represents the various modes of existence of a minimal autopoietic system. The boundary is a semipermeable membrane formed by only one component S. Throughout this membrane can permeate the nutrient A, which is transformed inside the system into S, the component that forms the boundary itself. The system is characterized by two competitive reactions, one that yields S from A, with velocity v gen; and one that breaks down S into the by-product P with velocity v dec. When the two velocities are numerically equal, the system is in homeostasis; when v gen is greater than v dec, the system can grow, and eventually enter a self-reproduction mode. Figure 4 clearly shows that self-reproduction can be seen as a particular kinetic pattern of homeostasis, i.e., a consequence of self-maintenance.

The minimal autopoietic system. This system is characterized by two competitive reactions, one that builds the component of the boundary, and another one that destroys it. According to the relative value of these two velocity constants, the system can be in homeostasis, or grow, or die
Finally, it should be mentioned that the primary literature on autopoiesis, in addition to nucleic acids, reproduction, and evolution, also avoids the use of the term "information". This is due mostly to Francisco Varela's deep concern—which I share—about the misuse of this term in most of the current bio-science literature. In part, the notion of information comes in, in autopoiesis, disguised under the term "cognition". This is also a vague term, and possibly also a considerable source of misunderstanding, as we shall see later on.
The interaction with the environment
Let us consider now the interaction between the autopoietic unit and its environment. As already pointed out, the living cell must be seen as an open system that receives energy and nutrients from the environment. We have here, then, an interesting contradiction between the biological autonomy and at the same time the dependence on the external medium. In fact, the living must operate within this contradiction.
The interaction with the environment, according to autopoiesis, is seen from the internal logic of the living system. In other words, the consequence of the interaction between an autopoietic unit and a given molecule X is not primarily dictated by the properties of the molecule X, but by the way in which this molecule is "seen" by the living organism.
As Varela puts it, "there is no particular nutrient value in sugar, except when the bacterium is crossing the sugar gradient and its metabolism utilizes the molecule so as to permit the continuity of its identity" (Varela 2000). Actually, the compounds that the living organism extracts from the environment to "create its own world" can be seen as something that the organism itself lacks—something that is missing and therefore must be obtained from the outside.
Maturana and Varela, in order to characterize the very particular nature of this interaction, use the term "cognition". They mean by that the following: that the internal structure of the living organism recognizes and utilizes the environment, be it water in the case of the fish and a spider web in the case of the spider. It simply implies the establishment of a specific complementarity between the organism and its environment—a sort of specific recognition, from which the use of the term cognition derives.
Varela (2000) recognizes that the use of this term is problematic, since it has a strong anthropomorphic connotation—one thinks automatically of human cognition. In fact, this has been frequently misunderstood (see, for example, Boden 2000)—particularly if the reader stops at the word without inquiring what the authors really mean by it. Most probably, it would have been better if Maturana and Varela had chosen a different term. On the other hand, Varela, in particular, likes to emphasize the continuity of the process of cognition; namely the notion that life proceeds by a hierarchical ladder of complexity levels. Thus, the cognition level of bacteria is the lowest, but it is still a rudimentary process of cognition (Varela and Maturana 1998; Maturana and Varela 1980). From this level, one can proceed to higher levels of cognition in insects, and then birds, mammals, and so to mankind.
Clearly cognition, so defined, is a fundamental aspect of life. In fact, Maturana and Varela state that there is an equivalence between life and cognition—that there cannot be one without the other. The question "what is cognition?" has been a primary quest in Maturana's and later in Varela's work, together with the question "what is life?". The two authors arrive at the surprising conclusion that the answers to these two questions actually converge. The mechanism of the living and cognition represent two aspects of the same process—the process of life.
At this point we can, however, bring in one element of criticism. In fact, this statement about the equivalence between cognition and life is not always present in Maturana's and Varela's papers on autopoiesis. Also, if cognition is so central to the essence of the living, then—one may argue—cognition should be taken into the definition of autopoiesis from the very start. However, cognition is not given in the list of criteria of autopoiesis; namely, in the criteria to assess whether a system is living or not. The choice of the authors has been to apply the notion of cognition only to systems that have first been found to be autopoietic according to the structural criteria. And perhaps, from a heuristic point of view, it is indeed better to consider the two levels (autopoiesis and cognition) as initially separated.
The cognitive interactions with the environment accompany the organisms during their life and during their biological evolution. Let us repeat the central point about this interaction: the environment has its own structural dynamics and, although independent of the organism, it does not prescribe or determine the changes in it. It induces a reaction in the organism, but the accepted changes are determined by the internal structure of the organism itself. It is the structure of the living system and its previous history of perturbations that determines what reactions the new perturbation will induce. These coupled interactions, accumulated over time, give a particular historical perspective to the autopoietic system: this becomes a historical product, the result of a long series of coupled interactions, the product of evolution. Accordingly, evolution is seen as a result of the maintenance of the internal structure of the autopoietic organism. Since the dynamics of the environment may be erratic, the result, in terms of evolution, is a natural drift, determined primarily by the inner coherence and autonomy of the living organism.
In this sense, Varela and Maturana's view (1998) is close to Kimura's (1985) theory of natural drift and to Jacob's (1994) notion of "bricolage". Evolution does not pursue any particular aim—it simply drifts. The path it chooses is not, however, completely random, but is one of the many that are in harmony with the inner structure of the autopoietic unit (Varela and Maturana 1998). According to this view, then, evolution can be seen as a way in which the internal coherence of a species compensates a given perturbation. When the "fit" is not possible, then the autopoietic structure will be destroyed.
The interaction with the environment also includes the interaction with other autopoietic units, i.e., it encompasses the whole world of mutual interactions among living organisms. In particular, as Varela and Maturana (1998) point out, the interaction between two autopoietic units can give rise to symbiosis, or to meta-cellular systems.
One step further: from biology to cognitive science
According to the view described above, the environment permits the actualization of the living, and the living, throughout its internal structure, picks up those elements from the external environment that will define its own world.
Here is where the thinking of Varela in particular (Varela et al. 1991) becomes closer to certain European philosophers, for example, Merleau-Ponty. Consider the following statement by Merleau-Ponty (1967): "it is the organism itself—according to the proper nature of its receptors, the thresholds of its nerve centers and the movements of the organs—which chooses the stimuli in the physical world to which it will be sensitive. The environment emerges from the world through the actualization or the being of the organism".
This is another important point in the autopoietic world: that the environment brings to life the organism and the organism creates the environment with its own perceptory sensorium. To express this process of mutual calling into existence, Varela and Maturana (1998) and later mostly Varela (1989, 2000) use the word "enaction". This is yet another neologism, and perhaps the simpler word "co-emergence" would also have been appropriate. The confluence expressed before between life and cognition is the best example of this co-emergence. According to Varela (2000), the simultaneous realization of the organic living structure and of cognition are two faces of the same phenomenon, "the phenomenon of life". This can be diagrammatically represented as in Fig. 5.

Pictorial representation of the process of enacting and co-emergence. The living structure and the organisms are here represented as separated domains only for heuristic reasons, in reality they are one
Once this view is accepted, there is no separation between the cognitive act and the organic structure of life—they are one. And if this is so, then the traditional Cartesian division between matter and mind disappears (Varela and Maturana 1998; Varela 2000; Varela et al. 1991). This is also true at the level of human cognition, a level at which the notion of consciousness appears. In this regard Varela and coworkers (Varela et al. 1991; Thompson and Varela 2001) use the term "embodiment" to signify that human consciousness has its counterpart in the organic structure—that there is no consciousness outside the reality of bodily experience. The strong emphasis of Varela on the tight relationship and complementarity between cognition and structure brings him almost naturally into the field of phenomenology, in particular to the philosophy of Husserl (1960). This is where biology and cognitive science merge into each other. Varela in particular (Varela et al. 1991) has carried out pioneer work on this continuum.
Chemical autopoiesis
Let us now go back to more chemical ground. Autopoiesis permits the creation of chemical models of cellular life that can be constructed in the laboratory.
The first structural prerequisite is a compartment with a spherical membrane, and this can be realized by utilizing lipids or other surfactants that make aggregates such as micelles or vesicles. In fact, this has been the research goal of the present writer's group since the early 1990s (Luisi and Varela 1989; Bachmann et al. 1992; Böhler et al. 1993; Walde et al. 1994a; Luisi 1996; Zepik et al. 2001). The basic idea of the work can be understood on the basis of the autopoietic model of Fig. 4. We have seen in this regard that such a model foresees a homeostatic model as well as a self-reproductive one. This second case turned out to be much easier to implement experimentally.
The first idea to build a self-reproductive autopoietic model stems from collaboration with Varela (Luisi and Varela 1989). We decided to utilize reverse micelles for a forced micro-compartmentation of two reagents, which could be up taken in the internal medium of the micelles and react inside the boundary (and not outside) to yield as a product the very surfactant that builds the boundary.
The experimental procedure that was actually applied is slightly different. In fact it was found easier to start from a water-insoluble precursor of the surfactant that binds to the membrane of the compartment, being eventually hydrolyzed there into the active surfactant (Bachmann et al. 1990, 1992; Luisi 1993; Luisi and Varela 1989; Zepik et al. 2001; Böhler et al. 1993). The first experiment was carried out with reverse micelles of caprylic acid (Bachmann et al. 1990); and later on with aqueous micelles of caprylate (Bachmann et al. 1990). This is represented in Fig. 6. Later, this autocatalytic growth scheme was extended to vesicles, in particular to oleate vesicles (Walde et al. 1994a), to vesicles of chiral methyl-dodecanoic acid (Morigaki et al. 1997), and to oleate giant vesicles (Wick et al. 1995). The mechanism is illustraded in Fig. 7.

A self-replicating micellar system. The autocatalytic cycle of self-reproduction of caprylate micelles (Bachmann et al. 1992). EC represents the water-insoluble ethyl caprylate, supernatant to an aqueous alkaline solution. Hydrolysis at the macroscopic interphase yields the first micelles as soon as the critical micelle concentration (CMC) is reached; afterwards the micelles rapidly take up EC, which is being hydrolyzed by physical micellar catalysis, yielding more caprylate and hence more micelles. As more micelles are formed, the more EC is hydrolyzed—a typical autocatalytic process, which brings about a population increase of the micelles. Since this process takes place and is caused by the micelles themselves, one can speak of self-reproduction. By changing the pH to more neutral conditions, a vesicle system is obtained, which works in the same way (see text)

The experimental procedure to obtain self-reproducing liposomes. S-S represents the water-insoluble precursor of the surfactant S, and S_S is hydrolysed on the membrane of the micelles or of the vesicles
These experiments also show the relation between autopoiesis and the autocatalytic mechanisms of self-reproduction. The basis of autopoiesis is self-maintenance and although in principle one could have cellular homeostasis without self-reproducing mechanisms, the most obvious way to attain the components' re-generation from within is via autocatalysis, In this sense, then, the experimental simple models of Figs. 6 and 7 are illustrative of the relation between autocatalytic and autopoietic mechanisms.
One question that rises at this point is whether and to what extent these self-reproducing systems fulfill the criteria of autopoiesis. The first question, whether there is a semipermeable boundary, can be answered positively; and one can also answer positively to the second criterion, namely that the boundary is of its own making. In fact, the membrane itself keeps building the bilayer as the hydrolysis reaction takes place on the membrane (Fig. 7). The positive answer to the third criterion, whether the system encompasses reactions that re-generate the components of the system, is not so straightforward. Reaction takes place on the boundary, which is part of the system, but a rather restricted one: the internal aqueous core is not part of the reaction system, and furthermore we are dealing with only one reaction instead of an internalized reaction network.
In all the experiments exemplified by Figs. 6 and 7, what is being reproduced is the empty shell. This is not suitable for making models of biological cells. It was later attempted (Böhler et al. 1993) to carry out a shell-and-core self-reproduction by using as guest molecules the nucleotide system of Von Kiedrowski (1986, 1989) solubilized in the core of reverse micelles. As the micelles reproduce according to the mechanism of Fig. 6, the hexanucleotide also replicates. In an extension of this work towards more realistic cell models, a system was described, consisting of self-replicating vesicles that contain an enzymatic system. The enzyme, β replicase, replicates RNA inside the vesicles: thus, while the vesicles self-reproduce, RNA self-replicates inside them (Oberholzer et al. 1995). In addition, a similar system in which poly (A)—a model for RNA—was being polymerized simultaneously with the self-reproduction of vesicles has been described (Walde et al. 1994b) and independently by the groups of Deamer and Joyce (Chakrabarti et al.1994).
Based on Fig. 3, a homeostatic mode of existence of a minimal autopoietic system can also be foreseen. This proved to be much more difficult, and could be realized only recently (Zepik et al. 2001). It is based on the oleic acid surfactant system as schematized in Fig. 8. Here, one reaction forms new oleate surfactant from the hydrolysis of the anhydride; and one reaction breaks down oleate via oxidation of the double bond. Depending upon whether the two velocities are equal or not, different pathways for the systems are obtained, namely homeostasis (which corresponds to an autopoietic self-maintenance system), growth and self-reproduction, or decay and death.

The experimental implementation of the autopoietic model of Fig. 3 with two competitive reactions. Here, one reaction forms new oleate surfactant from the hydrolysis of the anhydride, and one reaction destroys oleate via oxidation of the double bond. Depending upon whether the two velocities are equal or not, different pathways for the systems are obtained—homeostasis (which corresponds to an autopoietic self-maintenance system), growth and self-reproduction, or decay and death (Zepik et al. 2001)
Social autopoiesis
The main feature of autopoiesis is the self-maintenance due to a process of self-generation from within. Although this concept came from the analysis of a living cell, it can be metaphorically applied to social systems. Consider, for example, a political party, or a family, whereby the rules that define a party or a family can be seen as a kind of boundary formed by the (social) structure itself. These social structures are open systems: some members leave the structure, new people come in and are transformed into regular members by the rules of the party or of the family. Also, in all these systems, one can recognize certain characteristic features of biology, for example the notion of emergence—the family being an emergent property arising from the organization of individuals, and so on. Figure 9 is a qualitative picture of this situation, which is analogous to Fig. 2 depicted for a cell.

Towards social autopoiesis. The cyclic logic of autopoiesis applied to social systems. See the analogy with Fig. 2. The transfer from biology to social science assumes that the chemical interactions among the cell constituents are substituted by "human relationships" and that the membrane boundary is substituted by the defining rules of the social community
As we have already mentioned, social sciences picked up on this idea and it stirred up a great deal of intellectual excitement. The German sociologist N. Luhmann constructed an entire field based on social autopoiesis (Luhmann 1984), and autopoiesis was also applied to the juridical system (Teubner 1993), to literature (Paulson 1988) and into systemic family therapy, although with some dissenting voices (Varela 1989; Maturana 1987). Zeleny has also contributed to this field (Zeleny 1977, 1997); another author who has significantly developed the idea of social autopoiesis is John Mingers (1995, 1997). See also Capra (2002).
F. Varela remained somewhat skeptical about these extensions of autopoiesis. He says in this regard (Varela 2000): "These ideas are based, in my opinion, on an abuse of language. In autopoiesis, the notion of boundary has a more or less precise meaning. When, however, the net of processes is transformed into one "interaction among people", and the cellular membrane is transformed into the limit of a human group, one falls into an abuse, as I expressly said."
It must be said that Maturana had a more open mind on this topic (compare for example Varela 1989 with Maturana 1987). Despite Francisco Varela's doubts, it appears that, owing to the theory of autopoiesis, a field has been created that did not exist before, with new vistas and perspectives for understanding some aspects of social behavior, which can be taken as an indication of the fertility of autopoiesis.
Concluding remarks
Although the basic principles of autopoiesis are clearly represented in the 1974 paper (Varela et al. 1974), the theory has undergone a significant evolution over time, so that the overview one receives from a superficial reading of all papers and book may be confusing. In fact, especially in later works, certain aspects are emphasized much more than others—often in books more than in specific papers—and it is not easy to get a clear and complete view of autopoiesis on the basis of a single book or publication. These shortcomings of the primary literature have not facilitated the impact of autopoiesis; in fact, they have probably contributed to a misunderstanding of it. Another source of difficulty in autopoiesis is the use of various neologisms, such as cognition, enacting, "creating its own world", embodiment. The acceptance of all these terms and concepts is certainly one stumbling block, both from the scientific and psychological point of view.
In principle, however, autopoiesis can be easily understood without these new terms. Let me repeat the essential points here. According to autopoiesis, the main characteristic of the living system is self-maintenance based on self-reproduction from within the boundary; the interaction with the environment is seen in terms of the internal structure of the living; consequently the internal structure brings about a process of discrimination and selection (creating the environment, and cognition) of the elements of the environment, and the so chosen environment elicits in turn the coming into being of the living organism. This mutual co-emergence of environment and living structure (enacting, or co-emergence) gives rise to the process of life, and simultaneously and equivalently to the process of cognition. Cognition and co-emergence must be seen in a historical context, as the mutual interactions operate over time; thus, the autopoietic structure is a historical product (evolution).
This broad sketch implies as necessary tools self-organization, emergence, biological autonomy, and auto-referentiality. In this way, all the main concepts associated with life are gathered into a coherent picture. This picture also permits the construction of experimental testable models.
Concerning this last point, Gail Fleischaker (Fleischaker 1988) states that all that is alive must be autopoietic. Conversely, if something displays autopoiesis, this something must be living. I believe this is too strong a statement. Most people would rather accept the view that autopoiesis is the necessary, but not the sufficient, condition for life. A clear-cut discrimination between "necessary and sufficient conditions", on the one hand, and "necessary conditions" only, on the other hand, is not easy from the epistemological point of view. However, because of this difficulty, I believe it is safer to remain on the cautious side. For example, self-reproducing micelles and vesicles can be considered as the simplest autopoietic systems, but hardly as living ones. Interestingly, F. Varela did not object to the tagging as "living" of these micellar or vesicular systems (F. Varela, personal communication). His argument was an interesting one: that our notion of life is heavily permeated by a religious bias (the notion of soul), which makes it difficult to freely use the word "life" for simple chemical systems. Once one is liberated from these constraints, the term "life" may acquire a plainer and more usable meaning. More generally, these experimental models form the basis and indicate the direction on which to proceed to realize in the laboratory the minimal living condition.
The acceptance of the notion of cognition is important from another viewpoint, in that it permits us to construct a bridge between biology and cognitive sciences. I maintain that autopoiesis is the only available simple theory that is capable of providing a unitary view of the living, from the molecular level to the realm of human perception.
Despite all these advantages, autopoiesis has not had the impact in mainstream biological science that, according to many, it deserves. Why is this so? One part of the answer has already been mentioned: autopoiesis originated in a time-window (the early 1970s) when the biological science world was completely dominated by a vision of DNA and RNA as the holy grail of life. Alternative views about the mechanism of the living didn't have much chance of being taken seriously in mainstream journals. As mentioned earlier, however, this situation also reflects the intellectual choice of the authors, and partly perhaps their rigidity, as it would have been relatively easy to make autopoiesis compatible and complementary to the DNA/RNA world.
One can read in the modern literature of the life sciences some signals indicating that the notion of life is moving away from the simplistic equation nucleic acids = life. This in turn is accompanied by a revival of a systems theory of life that emphasizes collective properties such as self-assembly, self-organization, and emergence. In this new, perhaps more philosophically mature "Zeitgeist", autopoiesis could re-emerge as a very useful conceptual and experimental framework.
References
Bachmann PA, Walde P, Luisi PL, Lang J (1990) Self-replicating reverse micelles and chemical autopoiesis. J Am Chem Soc 112:8200–8201
Bachmann, PA, Luisi PL, Lang J (1992) Autocatalytic self-replicating micelles as models for prebiotic structures. Nature 357:57–59
Benseler F, Hejl P, Kock W (1980) Autopoiesis, communication and society: the theory of autopoietic systems in the social sciences. Campus, Frankfurt
Boden M (2000) Autopoiesis and life. Cognitive Sci Q 1:115–143
Böhler C, Bannwarth W, Luisi PL (1993) Self-replication of oligonucleotides in reverse micelles. Helv Chim Acta 76:2313–2320
Capra F (2002) The hidden connections: a science for sustainable living. HarperCollins, London
Chakrabarti AC, Breaker RR, Joyce GF, Deamer DW (1994) Production of RNA by a polymerase protein encapsulated within phospholipid vesicles. J Mol Evol 39:555–559
Deamer DW, Dworkin J, Sandford SA, Bernstein MP, Allamandola LJ (2003) The first cell membranes. Science
Fleischaker G (1988) Autopoiesis: the status of its system logic. BioSystems 22:37–49
Ganti T (1975) Organization of chemical reactions into dividing and metabolizing units: the chemotons. BioSystems 7:15–21
Husserl E (1960) Cartesian meditations: an introduction to phenomenology. Cairns D (trans). Kluwer, Dordrecht
Jacob F (1994) The possible and the actual. University of Washington Press, Seattle
Joyce GF (1994) Foreword. In: Deamer DW, Fleishaker GR (eds) Origins of life: the central concepts. John and Bartlett, Boston, pp xi–xii
Kauffman S (1995) At home in the Universe. Oxford University Press, New York
Kauffman S (2000) Investigations. Oxford University Press, New York
Kimura M (1985) The neutral theory of molecular evolution. New Sci 107:1464
Luhmann K (1984) Soziale Systeme. Suhrkamp, Frankfurt
Luisi PL (1993) Defining the transition to life: self-replicating bounded structures and chemical autopoiesis. In: Stein W, Varela F (eds) Thinking about biology. Addison-Wesley, New York
Luisi PL (1996) Self-reproduction of micelles and vesicles: models for the mechanisms of life from the perspective of compartmented chemistry. Adv Chem Phys 92:425–438
Luisi PL (1997) Self-reproduction of chemical structures and the question of the transition to life. In: Cosmovici CB, Bowyer S, Westhimer D (eds) Astronomical and biochemical origins and the search for life in the universe. Editrice Compositori, Bologna, pp 461–468
Luisi PL (1998) About various definitions of life. Origins Life Evol Biosphere 28:613–622
Luisi PL (2002) Emergence in chemistry: chemistry as the embodiment of emergence. Found Chem 4(3):183–200
Luisi PL, Varela F (1989) Self-replicating micelles: a chemical version of minimal autopoietic systems. Origins Life Evol Biosphere 19:633–643
Luisi PL, Lazcano A, Varela F (1996) What is life? Defining life and the transition to life. In: Rizzotti M (ed) Defining life: the central problem in theoretical biology. Edizione Università di Padova, Padua, pp146–167
Luisi PL, Oberholzer T, Lazcano A (2002) The notion of a DNA minimal cell: a general discourse and some guidelines for an experimental approach. Helv Chim Acta 85:1759–1777
Margulis L, Sagan D (1995) What is life. Simon and Schuster, New York
Maturana H (1970) Neurophysiology of cognition. In: Garvin P (ed) Cognition: a multiple view. Spartan, New York, pp 3–23
Maturana H (1987) The biological foundations of consciousness. Conference Workbook: texts in cybernetics. American Society for Cybernetics Conference, Felton, Calif.
Maturana H, Varela F (1980) Autopoiesis and cognition: the realization of the living. Reidel, Boston
Maturana H, Lettvin J, McCulloch W, Pitts W (1960) Life and cognition. Gen Physiol 43:129–175
McMullin B, Varela F (1997) Rediscovering computational autopoiesis. In: Husbands P, Harvey J (eds) Proceedings of the fourth European conference on artificial life. MIT Press, Cambridge, Mass.
Merleau-Ponty M (1967) The structure of behaviour. Beacon, Boston
Mingers J (1992) The problems of social autopoiesis. Int J Gen Syst 21:229–236
Mingers J (1995) Self-producing systems: implications and applications of autopoiesis. Plenum, New York
Mingers J (1997) A critical evaluation of Maturana's constructivist family therapy. Syst Practice 10(2):137–151
Morigaki K, Dellavalle S, Walde P, Colonna S, Luisi PL (1997) Autopoietic self-reproduction of chiral fatty acid vesicles. J Am Chem Soc 119:292–301
Morowitz H (1992) The beginning of cellular life. Yale University Press, New Haven, Conn.
Morowitz H, Peterson E, Chang S (1996) The synthesis of glutamic-acid in the absence of enzymes: implications for biogenesis. Origins Life Evol Biosphere 4:395–399
Oberholzer T, Wick R, Luisi PL, Biebricher CK (1995) Enzymatic RNA replication in self-reproducing vesicles: an approach to a minimal cell. Biochem Biophys Res Commun 207(1):250–257
Paulson W (1988) The noise as culture. Cornell University Press, Ithaca
Piaget J (1969) Essai sur les relations entre les régulations organiques et les processus cognitifs. In: Biologie et Connaissance. Gallimard, Paris
Pohorille A, Deamer DW (2002) Artificial cells: prospects for biotechnology. Trends Biotechnol 20:123–128
Szostak, JW, Bartell D, Luisi PL (2001) Synthesizing life. Nature 409:387–390
Teubner G (1993) Laws as an autopoietic system. Blackwell, Oxford
Thompson E, Varela F (2001) Radical embodiment: neural dynamics and consciousness. Trends Cognitive Sci 5:418–425
Varela F (1979) Principles of biological autonomy. North Holland/Elsevier, New York
Varela F (1989) Reflections on the circulation of concepts between a biology of cognition and systemic family therapy. Family Process 28:15–24
Varela F (2000) El fenómeno de la vida. Dolmen Esayo, Santiago, Chile
Varela F, Maturana H (1998) The tree of knowledge (rev edn). Shambala, Boston
Varela, F, Maturana H, Uribe R (1974) Autopoiesis: the organization of living systems, its characterization and a model. BioSystems 5:187–195
Varela F, Thompson E, Rosch E (1991) The embodied mind: cognitive science and human experience. MIT Press, Cambridge, Mass.
Von Kiedrowski G (1986) A self-replicating hexadeoxynucleotide. Angew Chem 98:932–934
Von Kiedrowski G (1989) Angew Chem 101:1259
Walde P, Wick R, Fresta M, Mangone A, Luisi PL (1994a) Autopoietic self-reproduction of fatty acid vesicles. J Am Chem Soc 116:11649–11654
Walde P, Goto A, Monnard PA, Wessicken M, Luisi PL (1994b) Oparin's reactions revisited: enzymatic synthesis of poly(adenylic acid) in micelles and self-reproducing vesicles. J Am Chem Soc 116:7541–7544
Wick R, Walde P, Luisi PL (1995) Autocatalytic self-reproduction of giant vesicles. J Am Chem Soc117:1435–1436
Zeleny M (1977) Self-organization of living systems formal model of autopoiesis. Int J Gen Syst 4:13–28
Zeleny M (ed) (1997) Autopoiesis: a theory of the living organization, North Holland, New York
Zepik HH, Bloechliger E, Luisi PL (2001) A chemical model of homeostasis. Angew Chem 40:199–202
Acknowledgements
The author expresses his thanks to Amy Cohen-Varela, who has edited the manuscript and contributed important critical suggestions.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Luisi, P.L. Autopoiesis: a review and a reappraisal. Naturwissenschaften 90, 49–59 (2003). https://doi.org/10.1007/s00114-002-0389-9
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00114-002-0389-9