
Abstract
Transcription factor 21 (TCF21) is a member of the basic helix-loop-helix (bHLH) transcription factors that mediate cell fate and differentiation by orchestrating temporal and spatial gene expression during the development of various organs. It plays a crucial role in a wide spectrum of biological processes, including organogenesis, epithelial-mesenchymal transition (EMT), cell cycle, autophagy, proliferation, differentiation, specification, maturation, and survival of cells, as well as invasion and metastasis of cancer cells. Controlled expression and activity of TCF21 provide a balanced transcriptional program that guarantees appropriate growth and maturation during embryogenesis and organ development. Its dysregulation is closely correlated with a variety of diseases, including cancer. Its function is mainly regulated by non-coding RNAs (ncRNAs), post-translational modifications (PTMs), and protein–protein interactions. However, the exact mechanisms of TCF21 dysregulation in disease progression are still elusive. This review summarizes the regulatory mechanisms of TCF21 expression and activity and highlights its critical role in health and disease. This information may contribute to the development of better diagnostics and treatments for cancer and other developmental diseases.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Basic helix-loop-helix (bHLH) transcription factors are crucial determinants that regulate lineage- and developmental-specific gene networks [1]. They are characterized by specific protein structures, such as the basic and HLH domains. The basic domain mediates their direct binding with DNA, whereas the HLH domain is involved in dimerization [2]. Currently, more than 240 bHLH family members have been identified in species ranging from yeast to human [3]. They are grouped into seven distinct classes (I to VII) according to their expression patterns, dimerization selectivity, and DNA-binding specificity [1]. bHLH proteins act as important regulators in a wide spectrum of biological processes through controlling the expression of many genes involved in proliferation [4], apoptosis [5], differentiation [6], autophagy [7], cell cycle [8], cell growth [9], development [10], angiogenesis [11], hematopoiesis [12], morphogenesis [13], and tumorigenesis [14]. Therefore, they play crucial roles in human health and disease, particularly cancer.
Transcription factor 21 (TCF21), also known as capsulin, epicardin, or Pod1, is a member of the class II bHLH transcription factor superfamily and is widely expressed in multiple tissues and organs [15,16,17]. It was first cloned in 1998 from various tissues and organs by five different laboratories simultaneously [15,16,17,18,19]. TCF21 has been shown to play crucial roles in organogenesis and the regulation of numerous cellular functions [20,21,22,23,24,25,26,27]. Loss of TCF21 results in abnormalities in multiple organs, as well as neonatal lethality [28, 29]. Its expression and activity are regulated by different mechanisms, such as non-coding RNAs (ncRNAs) [30], post-translational modifications (PTMs) [31], and protein–protein interactions [32]. A large number of studies have demonstrated that dysregulation of TCF21 has been correlated with many diseases, including cancer [23, 33,34,35,36]. This review focuses on the exact regulation mechanisms of TCF21, and on recent findings regarding its vital functions in health and cancer, presents important advances in the potential use of TCF21 as a biomarker in cancer therapy, and discusses future directions of research aimed at developing applications of TCF21 as a target for cancer treatment.
Overview: structure and functions of TCF21
Structure of TCF21
The TCF21 gene is located in the human chromosome 6q23-q24, and encodes an estimated 20 kDa functional TCF21 protein composed of 179 amino acids. Its structure is highly conserved across species. The bHLH domain of TCF21 mainly mediates its dimerization and direct interaction with DNA. It also contains an N-terminal interface domain (amino acids 2 to 100), which mediates its interaction with mitogen-activated protein kinase 1 (MEK1) [37]. Both the bHLH and the C-terminal domains (amino acids 80 to 179) of TCF21 are required to mediate the interaction between TCF21 and HeLa E-box-binding protein (HEB) [38]. Moreover, the N-terminal portions (amino acids 1 to 77) are required for the interaction of TCF21 with androgen receptor (AR) [39] (Fig. 1).

Schematic diagram of the TCF21 structure. Letters within the bar represent structural domains. The well-known proteins that interact with TCF21 are shown above the lines in the corresponding domains. Only representatives of TCF21-interacting proteins are shown. bHLH, basic helix-loop-helix DNA–binding domain; AR, androgen receptor; HEB, HeLa E-box-binding protein; MEK, mitogen-activated protein kinase
Functions of TCF21
TCF21 is a key bHLH transcription factor that regulates cell fate and differentiation during the development of various tissues and organs, such as muscle [40,41,42], gonads [43], testes [24, 39, 44], and spleen [45]. It is also strongly associated with organogenesis [28]. Besides, TCF21 mediates multiple physiological and pathological processes by targeting signaling pathways or inducing the expression of target genes involved in angiogenesis [46], endometriosis [36], epithelial–mesenchymal transition (EMT) [22], cell cycle [47], and autophagy [34]. It is also involved in the regulation of proinflammatory environment and active extracellular matrix (ECM) remodeling of visceral and subcutaneous fat pads [48]. In addition, TCF21 plays an important role in sex determination and differentiation [27, 49]. The multiple functions of TCF21 in biological processes indicate that its dysregulation is closely correlated with a number of diseases, particularly cancer. Therefore, further studies are required to explore its physiological functions and clinical applications.
Molecular mechanisms of TCF21 regulation
The expression and activity of TCF21 are regulated by distinct mechanisms, such as DNA methylation, ncRNAs, and PTMs. Here, we summarize the main modes of TCF21 regulation in physiological and pathological conditions.
Regulation of TCF21 at the transcription level
DNA methylation plays important roles in regulating numerous biological processes, such as development, aging, and tumorigenesis [50]. Aberrant DNA methylation is closely correlated with the occurrence and development of malignant tumors. Hypermethylation inhibits the expression of genes at the transcriptional level [51], whereas hypomethylation mainly influences chromosome stability and improves aneuploidy [52]. TCF21 promoter hypermethylation has been found in a variety of cancers, including head and neck [53, 54], lung [55,56,57], colon [58], kidney [59,60,61], breast [55], gastric [62], and urological [63] cancer. Correspondingly, low expression of TCF21 is observed in these malignant tumors. In addition to solid tumor tissues, aberrant TCF21 methylation has also been detected in sputum and urine specimens from patients with cancer [55, 59]. Gooskens et al. demonstrated that hypermethylation of TCF21 promoter silences its expression, whereas TCF21 overexpression suppresses clonogenic proliferation and migration of renal tumor cells [60]. Moreover, TCF21 hypermethylation is significantly associated with lymph node invasion in colon cancer patients [58]. These studies suggest that the detection of TCF21 methylation status has the potential to be used in clinical applications.
p53 is a well-studied transcription factor that acts as a tumor suppressor in malignant diseases, and its gene is known to be frequently mutated in human cancers [64,65,66]. Li et al. showed that TCF21 is a direct target gene of p53. Its expression in p53-positive cells is significantly increased compared with that in p53-negative cells. They also showed that TCF21 promoter contains four potential p53-binding motifs. p53 binds to the TCF21 promoter and promotes its expression to respond to hypoxic stress [37]. Sex-determining region Y (SRY) is a key regulator of male sex determination, acting as a master switch that regulates the testis determination pathway [67]. Bhandari et al. revealed that overexpression of SRY increases the transcription activity of the TCF21 promoter. They also found that SRY directly binds to the SRY/SOX9 response elements in the TCF21 promoter and activate its expression during fetal rat testis development. A mutation of SRY/SOX9 response elements leads to the loss of SRY actions on TCF21 promoter [44]. In addition, analysis of DNA sequence showed that the TCF21 promoter also possesses other transcription factor binding motifs, such as SOX9 and GATA4, indicating the potential regulation of TCF21 expression by other transcription factors [44].
Post-transcriptional regulation of TCF21 by non-coding RNAs
Non-coding RNAs (ncRNAs) are crucial factors that play an important role in regulating gene expression and chromatin structure [68]. They are divided into different classes according to their specific biogenesis and function, including long non-coding RNA (lncRNA), piwi-interacting RNA (piRNA), microRNA (miRNA), small interfering RNA (siRNA), small nuclear RNA (snRNA), and circular RNA (circRNA) [50, 69]. In recent years, growing studies show that ncRNAs contribute to the post-transcriptional regulation of TCF21 (Table 1).
MicroRNAs are small non-coding RNAs that regulate the expression of genes involved in many crucial physiological and pathological processes [74,75,76]. They target the 3′-untranslated region (3′UTR) of gene messenger RNAs (mRNAs) to repress these genes translation [77]. TCF21 is a direct target of miR-224 and miR-146a in coronary heart disease. miR-224 binds to the TCF21 3′UTR C variant, and inhibits TCF21 expression at the transcription level in human coronary artery smooth muscle cells [78]. miR-146a directly binds to TCF21 promoter. However, rs2910164 in miR-146a disrupts its binding to TCF21 promoter and increases the risk of coronary heart disease in an Iranian population [79]. TCF21 is also regulated by several miRNAs in cancer, including miR-205, miR-92a, miR-3648, miR21, and miR-1228. miR-205 promotes invasion of ovarian cancer cells by directly targeting TCF21. Consistent with this, miR-205 expression is negatively correlated with TCF21 expression in patients with ovarian cancer [70]. TCF21 is also a potential target of miR-92a in osteosarcoma cells. Overexpression of miR-92a promotes proliferation, invasion, growth, and metastasis of osteosarcoma cells by inhibiting TCF21 expression [73]. Moreover, miR-3648 promotes invasion and metastasis of human bladder cancer by negatively regulating TCF21 [80]. In renal cancer, miR-21 overexpression promotes invasion of renal cell carcinoma Caki-1 cells by directly targeting TCF21 [72]. Similar to miR-205, a negative correlation is observed between expression of miR-1228 and TCF21 in lung cancer samples. miR-1228 directly interacts with TCF21 mRNA and inhibits TCF21 expression in lung cancer cells [71].
Circular RNAs are a class of endogenous non-coding RNAs characterized by a stable cyclic structure [81]. Growing evidence shows that circRNAs play a crucial role in human health and diseases, particularly in cancer [82,83,84]. It is well defined that circRNAs regulate gene expression by sponging miRNAs [85,86,87]. TCF21 is an indirect target of circRNAs. For example, overexpression of hsa_circ_100395 inhibits proliferation, migration, and invasion of lung cancer cells through upregulation of TCF21 by sponging miR-1228, indicating a novel regulatory loop of hsa_circ_100395/miR-1228/TCF21 axis in lung carcinogenesis [71].
Long non-coding RNAs are a class of important regulators more than 200 nucleotides in length, which participate in a number of cellular functions and disease processes [88, 89]. They act as endogenous miRNA sponges to regulate gene expression involved in carcinogenesis [90,91,92]. They can also alter epigenetic signatures through interactions with chromatin remodeling enzymes [93,94,95]. TCF21 can be regulated by lncRNAs. For instance, lncRNA TCF21 antisense RNA inducing demethylation (TARID) promotes TCF21 expression through inducing promoter demethylation. This process is mediated by growth arrest and DNA damage-inducible protein 45 alpha (GADD45A), a crucial regulator of DNA demethylation. TARID interacts with GADD45A and then recruits thymine-DNA glycosylase to promote demethylation of the TCF21 promoter, leading to upregulation of TCF21 [30]. TCF21 is also a target of lncRNA LINC00163 in lung cancer cells. LINC00163 overexpression significantly inhibits proliferation, migration, and invasion of lung cancer cells in vitro and impairs tumor propagation in vivo through upregulation of TCF21 [96]. These studies reveal a complex network of ncRNAs in TCF21 regulation, which are involved in TCF21-related disease progression, including cancer. Further studies are required to fully understand the relation of ncRNAs to TCF21 for TCF21-based treatment strategies.
Post-translational modifications contribute to TCF21 regulation
PTMs are dynamic reversible processes that play crucial roles in regulating protein functions, such as protein stability, subcellular translocation, DNA-binding affinity, protein folding, and interaction with other proteins [50]. Common PTMs in mammalian include phosphorylation, sumoylation, ubiquitination, methylation, succinylation, and acetylation [77, 97, 98].
Growing evidence suggests that TCF21 is regulated by PTMs, including phosphorylation, ubiquitination, and sumoylation (Fig. 2). Phosphorylation is a well-studied PTM that plays an important role in regulating a variety of cellular functions, such as cell growth, proliferation, apoptosis, and cell cycle [99, 100]. A large number of transcription factors, such as forkhead box protein K2 (FOXK2), forkhead box protein O3a (FOXO3a), and p53, have been shown to be regulated by phosphorylation [101,102,103]. TCF21 is a target of phosphorylation. Mass spectrometry analysis revealed that three phosphorylation sites (serine residues S37, S48, and S67) exist in TCF21 amino acid sequence (Fig. 1). These phosphorylation sites are localized in the N-terminal region of TCF21 structure and are evolutionarily conserved across species, indicating the functional role of TCF21 phosphorylation in the specification and maturation of proepicardial cells [31]. Sumoylation is a crucial post-translational modification involved in regulation of protein functions, such as protein subcellular translocation, protein half-life, and interaction between proteins [104]. Our previous study demonstrated that TCF21 can be sumoylated by small ubiquitin-like modifier 1 (SUMO1) at lysine residue 24, and SUMO-specific protease 1 (SENP1) mediates its desumoylation. The study further showed that sumoylation can extend the half-life of TCF21 but cannot change its subcellular localization. It also found that TCF21 sumoylation enhances its inhibition on the transcriptional activity of estrogen receptor-α (ERα) by recruiting histone deacetylases 1/2 (HDAC1/2), leading to the suppression of ERα-positive breast cancer cell growth [105]. PTMs have been reported to interact with each other, thus exploring the PTMs network of TCF21 is crucial to understanding the mechanisms of TCF21 regulation.

Regulation of TCF21 by post-translational modifications (PTMs). TCF21 is sumoylated by the small ubiquitin-like modifier 1 (SUMO1). Sumoylation of TCF21 inhibits the transcriptional activity of estrogen receptor-α (ERα) by recruiting histone deacetylases 1/2 (HDAC1/2). SUMO-specific protease 1 (SENP1) mediates the desumoylation of TCF21. Mutation of TCF21sumoylation site enhances the ubiquitination of TCF21, leading to its degradation in a ubiquitin/proteasome-dependent manner
Implication of TCF21 in disease development
The role of TCF21 in non-neoplastic diseases
TCF21 is shown to be associated with several cardiovascular diseases, such as coronary artery disease (CAD), ventricular septal defects, and hypertension. It has been reported that TCF21 inhibits differentiation of human coronary artery smooth muscle cells and decreases CAD risk [106]. It specifically binds to the conserved sequence CAGCTG in target genes and regulates a network of genes associated with CAD [107]. TCF21 also mediates the regulation of disease-related growth factor and embryonic signaling pathways on CAD risk [35]. In addition, TCF21 rs12190287 is reported to increase the susceptibility to CAD in an Iranian population [79]. rs12190287 has also been found to increase the susceptibility to hypertension [108] and ventricular septal defects [109].
TCF21 is also linked with the development of endometriosis and proteinuric renal disease. Endometriosis affects 10–15% of women of reproductive age [110]. TCF21 expression is found to be significantly increased in endometriotic stromal cells compared with that in endometrial stromal cells. Knockdown of TCF21 in endometriotic xenografts results in the abrogation of ectopic lesion growth in mice [36], indicating the potential of TCF21 as a target for endometriosis treatment. Podocytes are central components of glomerular filtration barrier [111]. Defects of podocyte differentiation cause glomerular injury, leading to proteinuria and glomerulosclerosis [112]. TCF21 is shown to highly express in podocytes. Knockdown of TCF21 in podocytes leads to glomerular abnormality with a decreased number of endothelial and mesangial cells. Moreover, 40% of 5-week-old podocyte-specific TCF21 knockout mice develop massive proteinuria and similar lesions with focal segmental glomerulosclerosis [33].
The role of TCF21 in carcinogenesis
It is well-known that TCF21 plays an important role in regulating cellular functions, including proliferation, differentiation, epithelial–mesenchymal transition, invasion, metastasis, cell cycle, autophagy, and survival. Thus, its dysregulation is closely related to a number of cancers (Table 2). A deeper understanding of TCF21 mechanisms in carcinogenesis will contribute to better diagnosis and treatment of cancer.
TCF21 targets signaling pathways in cancer
Growing evidence shows that TCF21 is involved in the regulation of signaling pathways in cancer. Understanding its mechanism in signaling pathway regulation may provide us new insights into cancer progression. PI3K/AKT signaling pathway modulates a variety of cellular functions during cancer progression, such as cell proliferation, metastasis, and drug resistance [123]. It has been reported that overexpression of TCF21 significantly downregulates the expression of PI3K and p-AKT in human colorectal cancer cell lines [118]. Another study showed that knockdown of TCF21 increases p-AKT expression, whereas overexpression of TCF21 decreases p-AKT expression in gastric cancer cell lines. In gastric cells treated with LY294002 (AKT inhibitor), knockdown of TCF21 failed to reduce CDDP-induced apoptosis [20]. In addition, overexpression of TCF21 is shown to significantly decrease the expression of p-PI3K and p-AKT in cholangiocarcinoma cells. Administration of 740 Y-P (PI3K activator) significantly recovered p-PI3K, and p-Akt levels suppressed by TCF21 in cholangiocarcinoma cells [46]. These studies suggest that TCF21 may act as a tumor suppressor by targeting PI3K/AKT signaling pathway.
Mitogen-activated protein kinases/extracellular signal-regulated kinase (MAPK/ERK) signaling pathway plays a crucial role in regulating fundamental cellular processes, such as cell proliferation, differentiation, migration, senescence, and apoptosis. Dysregulation of MAPK/ERK cascade is closely associated with many aspects of carcinogenesis [124, 125]. In cholangiocarcinoma cells, overexpression of TCF21 is shown to significantly downregulate the expression of p-ERK1/2, without affecting the total protein expression [46]. In another study, overexpression of TCF21 significantly reduces the expression of p-ERK1/2 in uterine corpus endometrial carcinoma cells. They also found that TCF21 directly interacts with MEK1 through its N-terminal interface domain (2–100) and inhibits the kinase activity of MEK1 by blocking the key functional domain of MEK1, leading to downregulation of p-ERK1/2 level [37]. These studies indicate that TCF21 suppresses cancer progression by inhibiting MAPK Pathway.
Estrogen receptor signaling pathway mediates estrogen-stimulated proliferation, migration, and survival of target cells. This pathway is the target of hormone therapy for ER-positive breast cancer [126]. In previous study, we found that TCF21 suppresses the growth of ERα-positive breast cancer cells by targeting estrogen receptor signaling pathway. TCF21 directly interacts with ERα, and inhibits its transcriptional activity in a HDACs-dependent manner [105]. Taken together, these studies suggest that TCF21 may be a core factor in the network of signaling regulation in cancer progression.
TCF21 regulates proliferation, apoptosis, and cell cycle
The main characteristics of cancer cells are their persistent proliferation and evasion of apoptosis [127]. A growing number of studies show that TCF21 plays an important role in carcinogenesis as a tumor suppressor (Fig. 3). Consistent with this, the expression of TCF21 has been shown to be significantly decreased in tumor tissues compared with that in adjacent normal tissues [22, 61, 113]. It has been reported that the anti-tumoral functions of TCF21 are closely related to inhibiting proliferation and promoting apoptosis in various types of cancer, such as colorectal [58], gastric [20], lung [34], breast [113], and liver cancer [128], as well as esophageal squamous cell carcinoma [22] and uterine corpus endometrial carcinoma [37].

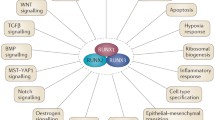
The role of TCF21 in cancer progression. TCF21 mainly acts as a tumor suppressor by inhibiting proliferation, cell cycle, epithelial–mesenchymal transition (EMT), invasion, metastasis, autophagy, and angiogenesis, and by inducing apoptosis in various types of cancer
The anti-tumoral characteristics of TCF21 may be partly due to its negative regulation on cell cycle progression. Cyclin D1 is a crucial intracellular mediator of extracellular signals that control proliferation, and is involved in regulation of the cell cycle and pathological process [129]. In our previous work, we found that TCF21 significantly inhibits the transcriptional activity of cyclin D1 in ERα-positive breast cancer cell lines MCF-7 and ZR-75-30, and that overexpression of TCF21 results in cell cycle arrest of MCF-7. Furthermore, TCF21 directly interacts with ERα and inhibits its transcriptional activity, leading to the suppression of MCF-7 cell proliferation [105]. Cyclin E1 is another key regulator in cell cycle progression due to its role in inducing S phase entry [130]. Franca et al. showed that TCF21 overexpression significantly increases cyclin E1 expression through targeting SHP in hepatocarcinoma cell line HepG2 [117]. Moreover, TCF21 overexpression inhibits the transition of gastric cancer cells from G1 to S phase [20]. These findings indicate the crucial regulatory role of TCF21 in cell cycle balance.
In addition, TCF21 represses proliferation of uterine endothelium tumor cells through inhibiting MAPK signaling pathway [37]. Molecular studies on colorectal and gastric cancer cells revealed that TCF21 overexpression inhibits proliferation of tumor cells by repressing PI3K/AKT signaling pathway [20, 118]. These studies indicate that TCF21 exerts its anti-tumoral effect in different cancers through diverse mechanisms. Further studies are needed to investigate the exact mechanism of TCF21 in carcinogenesis, which may provide new insights for the TCF21-based therapeutics strategy in cancer.
TCF21 in tumor angiogenesis
Abnormal angiogenesis is shown to be responsible for multiple pathological processes, including carcinogenesis [131, 132]. Tumor angiogenesis contributes to tumor growth by supplying oxygen and nutrients to cancer cells, while its suppression results in tumor stasis [133]. It has been reported that tumors lacking of TCF21 in nude mice are more vascular than tumors that expressed wild-type TCF21, indicating the potential inhibitory role of TCF21 on tumor angiogenesis [53]. Vascular endothelial growth factor (VEGF) and platelet-derived growth factor (PDGF) are crucial pro-angiogenic factors that maintain the angiogenic phenotype of vessel cells, which contribute to carcinogenesis [134]. Duan et al. demonstrated that overexpression of TCF21 significantly inhibits angiogenesis in cholangiocarcinoma xenograft tumors, and targets the expression and secretion of VEGF and PDGF in vitro and in vivo. The suppression of VEGF and PDGF production by TCF21, in turn, reduces proliferation, invasiveness, and tube formation of endothelial cells. The study also showed that TCF21 exerts its anti-angiogenic influence by targeting PI3K/AKT and ERK1/2 signaling pathways in cholangiocarcinoma [46]. Despite the progress that has been made in investigating the influence of TCF21 on tumor angiogenesis, further studies are needed to elucidate its exact mechanism, which leads to the development of novel TCF21-based anti-angiogenesis therapeutics.
TCF21 and autophagy
Growing studies suggest that autophagy is closely associated with the development of many diseases, including cancer [135]. Autophagy is a double-edged sword in cancer progression. Normally, it can induce cell death by activating apoptosis signaling pathways. However, an increasing number of studies show that autophagy also promotes cancer cell survival during treatment [136]. For instance, a correlation between TCF21 expression and autophagy is observed in lung cancer progression. TCF21 knockdown may promote global DNA methylation of non-small cell lung cancer cells, leading to increased autophagy. This process contributes to lung cancer progression. In turn, autophagy inhibition by 3-methyladenine (3-MA), an autophagy inhibitor, significantly promotes TCF21 promoter methylation, leading to upregulation of TCF21 expression and enhanced apoptosis [34]. Research on the role of TCF21 in regulating autophagy is currently limited. Further studies are thus required to elucidate its exact mechanisms.
The role of TCF21 in EMT
EMT plays a crucial role in many physiological processes, such as embryonic development, mesoderm formation, and tissue repair. Aberrant activation of EMT contributes pathologically to a number of diseases, including cancer [50, 137, 138]. During the EMT process, the expression of epithelial markers is decreased, whereas the expression of mesenchymal markers is increased [50].
TCF21 has been shown to correlate with EMT in a number of cancer types. For instance, TCF21 expression is essential in maintaining the epithelial phenotype of cells. Overexpression of TCF21 results in increased expression of epithelial markers WNT4 and CDH1, and decreased expression of mesenchymal markers SNAI1 and vimentin in lung cancer cell line A549 [53]. Moreover, high expression of WNT4 is also observed in TCF21-positive lung cancer cells [55], indicating that WNT4 may be one of the targets of TCF21. Cancer stem cells (CSCs) are generated during EMT progress, which are recognized as the main cause of therapeutic resistance, metastasis, and recurrence of cancer [115, 139]. Upregulation of TCF21 induced by curcumin is shown to inhibit the emergence of CSCs in lung cancer. In breast cancer cell line MDA-MB-231, ectopic TCF21 expression significantly decreases SNAI1 expression, and inhibits the EMT [113]. Furthermore, TCF21 overexpression in colorectal cancer cells significantly downregulates the expression of mesenchymal marker vimentin and upregulates the expression of epithelial marker E-cadherin [58], indicating its inhibitory role in the EMT of colorectal cancer. However, overexpression of TCF21 in renal cancer cells significantly upregulates the expression of epithelial marker E-cadherin at both the mRNA and the protein levels, but does not affect the mRNA level of vimentin and SNAI1 [60]. Taken together, these studies confirm the involvement of TCF21 in regulating EMT progress by controlling the expression of EMT-related genes such as WNT4, CDH1, vimentin, SNAI1, and E-cadherin.
TCF21 controls invasion and metastasis
Invasion and metastasis are the most life-threatening aspects of cancer progression [140]. Metastasis is a multistep pathological process characterized by the transfer of cancer cells from one primary site to other organs or tissues, while invasion is the first step toward metastasis and one of the major features of cancer [141, 142]. TCF21 has been identified as a key regulator of invasion and metastasis in a variety of cancer types. For instance, TCF21 expression or its SNPs have been shown to be closely associated with metastasis in some cancers, such as breast cancer [113], osteosarcoma [48], and hepatocellular carcinoma [143]. Overexpression of TCF21 is reported to inhibit invasion and metastasis in a number of cancers, including esophageal squamous cell carcinoma [22], uterine corpus endometrial carcinoma [37], melanoma [23], as well as ovarian [70], bladder [80], gastric [20], lung [34], and colorectal cancer [58, 118]. Chen et al. demonstrated that overexpression of TCF21 significantly upregulates Kiss-1 level and reverses EMT-related proteins (E-cadherin, N-cadherin, snail, twist, and vimentin), leading to inhibition of invasion and metastasis in esophageal squamous cell carcinoma [22]. Similarly, TCF21 also suppresses invasion and metastasis in bladder cancer [80] and metastatic melanoma by targeting Kiss-1 [23]. In addition, TCF21 overexpression inhibits migration and invasion of colorectal cancer cells through the inhibition of PI3K/AKT signaling and matrix metalloproteinase 2 (MMP2) and MMP9 expression [118]. TCF21 also represses invasion of ovarian cancer cells by downregulation of MMP2 and MMP10 [70]. Although progress has been made in exploring the regulatory mechanisms of invasion and metastasis, many aspects still need to be clarified.
TCF21 has been shown to play a crucial role in mediating the response of cancer cells to oxidative stress and hypoxia [37, 144]. Thioredoxin reductase 1 (TXNRD1) is an oxidoreductase that maintains reactive oxygen species and superoxide concentration. It has been reported that oxidative stress induced by H2O2 increases miR526b/miR655 expression in breast cancer cell line MCF-7. Upregulation of miR526b/miR655 increases TXNRD1 expression by targeting TCF21 [144]. Moreover, TCF21 is a hypoxia-driven p53 target and responds to hypoxia stress by inhibiting MAPK signaling pathway in uterine corpus endometrial carcinoma [37].
TCF21 as a biomarker and therapeutic target in cancer
Although, in recent years, many advances in the strategies of cancer diagnosis and treatment have been made, the incidence and mortality of cancer are still increasing, causing tremendous pain and heavy financial burdens to patients and their families. A large number of cancer patients still diagnosed in advanced stages due to the difficulty of observing early clinical symptoms. The identification of valuable biomarkers is very important for early screening and prognostic judgments of cancer treatments. A growing number of studies indicate that TCF21 is a promising candidate biomarker for the diagnosis, prognosis, and treatment of many types of cancer. For instance, the methylation pattern of the TCF21 promoter offers a sensitive diagnostic biomarker for several types of cancer, including colorectal [58], gastric [54], and lung [56, 57] cancer, as well as neck squamous cell carcinoma [54], metastatic melanoma [23], and urological cancers [59, 63]. In addition, low expression of TCF21 has been shown to correlate with poor prognosis in colorectal cancer [118], lung adenocarcinoma [116], clear cell renal cell carcinoma [119], hepatocellular carcinoma [143], esophageal squamous cell carcinoma [22], and lung cancer [115]. However, low expression of TCF21 is identified as an independent biomarker for favorable overall disease-specific survival in patients with head neck squamous cell carcinoma [120]. The opposite results may suggest that the prognostic values of TCF21 expression are diverse in different types of cancer. The clinical significance of TCF21 as a candidate biomarker has been validated by many small-scale studies. However, large-scale studies are still required to confirm possible clinical applications of TCF21 as a biomarker during cancer treatment.
Owing to its essential role in cancer progression, TCF21 has the potential to be an efficient therapeutic target. TCF21 overexpression has been reported to inhibit proliferation, migration, invasion, and metastasis in a variety of cancers [20, 58, 70, 143], which is indicative of its therapeutic value. Ectopic TCF21 expression also suppresses the EMT in lung, breast, renal, and colorectal cancer [53, 58, 60, 113]. Therefore, screening of natural products for drug discovery and synthesis of chemotherapeutic drugs targeting TCF21 may provide valuable and efficient therapeutic strategies for cancer treatment. In addition, as TCF21 has been shown to be a direct target of some miRNAs, circRNAs, and lncRNAs [30, 70, 71], screening or synthesis of novel drugs targeting these ncRNAs may also contribute to valuable therapeutic strategies. Although available data confirm the potential clinical applications of TCF21, further studies are needed to elucidate its mechanisms and effects in chemotherapy sensitivity and resistance.
Conclusions and perspectives
TCF21 is recognized as a key regulator in many crucial biological processes, including cell proliferation, differentiation, survival, cell cycle, EMT, invasion, metastasis, autophagy, specification, and maturation of cells as well as organogenesis. This may depend on its following characteristics: Firstly, TCF21 acts as a transcription factor to regulate the expression of its target genes involved in these progresses. Secondly, TCF21 targets crucial signaling pathways, such as PI3K/AKT, MAPK, and miRNAs. It may also cross talk with other signaling pathways. Thirdly, it regulates cellular functions by cooperating with other regulators, such as JUN, SMAD3, and HDAC1/2. Therefore, a controllable regulation of TCF21 expression and activity provides a balanced transcriptional network that guarantees proper growth and maturation during embryogenesis and organ development. Conversely, the dysregulation of TCF21 function contributes to the occurrence of a number of diseases, including cancer. Owing to its central role in transcriptional regulation, it has great potential as a therapeutic target for a number of cancers. For instance, ERα is a well-known efficient target of endocrine therapy in breast cancer. Our previous work demonstrated that TCF21 significantly inhibits the transcriptional activity of ERα in an HDAC-dependent manner in breast cancer [105], indicating its essential role in determining the sensitivity or resistance of endocrine treatment. As TCF21 is an effective tumor suppressor, its regulation by agonists both in normal and disease conditions may be useful in both preventing and treating a wide variety of cancers. Therefore, the identification of specific efficient TCF21 agonists could be a promising strategy for cancer treatment. In addition, as ncRNAs have been shown to be important regulators of TCF21, targeting the ncRNA-TCF21 axis may provide a new strategy for cancer treatment. Considering the complexity of the TCF21 network and its cross talk with other transcription factors, continued in-depth investigation of its role in health and in cancer is vital for providing novel TCF21-based therapeutic strategies for cancer patients. Nevertheless, targeting TCF21 can be a double-edged sword because of its key role in the development of normal tissues and organs, which may also affect carcinogenesis. The current challenge is to regulate TCF21 expression and activity in particular pathological conditions, which will need exact information on the regulation and activity of TCF21 in health and in cancer.
Abbreviations
- TCF21:
-
transcription factor 21
- bHLH,:
-
basic helix–loop–helix
- EMT,:
-
epithelial–mesenchymal transition
- ncRNAs,:
-
non-coding RNAs; PTMs, post-translational modifications
- MEK,:
-
mitogen-activated protein kinase 1
- SRY:
-
sex-determining region Y
- HEB:
-
HeLa E-box-binding protein
- AR:
-
androgen receptor
- ECM:
-
extracellular matrix
- IL6:
-
interleukin 6
- Scx:
-
scleraxis
- SF1:
-
steroidogenic factor 1
- ChIP:
-
chromatin immunoprecipitation
- lncRNA:
-
long non-coding RNA
- piRNA:
-
piwi-interacting RNA
- miRNA:
-
microRNA
- mRNAs:
-
messenger RNAs
- siRNA:
-
small interfering RNA
- snRNA:
-
small nuclear RNA
- circRNA:
-
circular RNA
- 3′UTR:
-
3′-untranslated region
- TARID:
-
TCF21 antisense RNA-inducing demethylation
- GADD45A:
-
growth arrest and DNA damage-inducible protein 45 alpha
- FOXK2:
-
forkhead box protein K2
- FOXO3a:
-
forkhead box protein O3a
- SNPs:
-
single-nucleotide polymorphisms
- SUMO1:
-
small ubiquitin-like modifier 1
- SENP1:
-
SUMO-specific protease 1
- ERα:
-
estrogen receptor-α
- HDAC1/2:
-
histone deacetylases 1/2
- CAD:
-
coronary artery disease
- AHR:
-
aryl hydrocarbon receptor
- AP-1:
-
activated protein-1
- ERβ:
-
estrogen receptor beta
- USF2:
-
upstream stimulatory factor 2
- MAPK:
-
mitogen-activated protein kinase
- ERK:
-
extracellular signal-regulated kinase
- MMP2:
-
matrix metalloproteinase 2
- VEGF:
-
vascular endothelial growth factor
- PDGF:
-
platelet-derived growth factor
- 3-MA:
-
3-methyladenine
References
Murre C (2019) Helix–loop–helix proteins and the advent of cellular diversity: 30 years of discovery. Genes Dev 33:6–25
Dennis DJ, Han S, Schuurmans C (2019) bHLH transcription factors in neural development, disease, and reprogramming. Brain Res 1705:48–65
Jiang X, Yang Z (2018) Multiple biological functions of transcription factor 21 in the development of various cancers. OncoTargets Ther 11:3533–3539
Zhang X, Liu R, Zhao N, Ji S, Hao J (2019) Sohlh2 inhibits breast cancer cell proliferation by suppressing Wnt/β-atenin signaling pathway. Mol Carcinog 58(6):1008–1018
Liu Q, Wu Y, Yoshizawa T, Yan X, Morohashi S, Seino H, Kato Y, Kijima H (2016) Basic helix-loop-helix transcription factor DEC2 functions as an anti-apoptotic factor during paclitaxel-induced apoptosis in human prostate cancer cells. Int J Mol Med 38(6):1727–1733
Kokabu S, Nakatomi C, Matsubara T, Ono Y, Addison WN, Lowery JW, Urata M, Hudnall AM, Hitomi S, Nakatomi M (2017) The transcriptional co-repressor TLE3 regulates myogenic differentiation by repressing the activity of the MyoD transcription factor. J Biol Chem 292(31):jbc.M116.774570
Martina JA, Yong C, Marjan G, Rosa P (2012) MTORC1 functions as a transcriptional regulator of autophagy by preventing nuclear transport of TFEB. Autophagy 8(6):903–914
Gallo C, Fragliasso V, Donati B, Torricelli F, Tameni A, Piana S, Ciarrocchi A (2018) The bHLH transcription factor DEC1 promotes thyroid cancer aggressiveness by the interplay with NOTCH1. Cell Death Dis 9(9):871
Jiang G, Huang C, Liao X, Li J, Huang C (2019) The RING domain in the anti-apoptotic protein XIAP stabilizes c-Myc protein and preserves anchorage-independent growth of bladder cancer cells. J Biol Chem 294(15):jbc.RA118.005621
Jin K, Xiang M (2019) Transcription factor Ptf1a in development, diseases and reprogramming. Cell Mol Life Sci : CMLS 76(5):921–940
Tanaka A, Itoh FK, Takezawa T, Kurihara H, Itoh S, Kato M (2010) Inhibition of endothelial cell activation by bHLH protein E2-2 and its impairment of angiogenesis. Blood 115(20):4138–4147
Gasiewicz TA, Singh KP, Casado FL (2010) The aryl hydrocarbon receptor has an important role in the regulation of hematopoiesis: implications for benzene-induced hematopoietic toxicity. Chem Biol Interact 184(1):246–251
de la Parra J, Cuartero MI, Perez-Ruiz A, Garcia-Culebras A, Martin R (2018) AhR deletion promotes aberrant morphogenesis and synaptic activity of adult-generated granule neurons and impairs hippocampus-dependent memory. eNeuro 5(4):ENEURO.0370-17.2018
Mikheev AM, Mikheeva SA, Severs LJ, Funk CC, Huang L, Mcfaline-Figueroa JL, Schwensen J, Trapnell C, Price ND, Wong S (2018) Targeting TWIST1 through loss of function inhibits tumorigenicity of human glioblastoma. Mol Oncol 12(7):1188–1202
Lu J, Richardson JA, Olson EN (1998) Capsulin: a novel bHLH transcription factor expressed in epicardial progenitors and mesenchyme of visceral organs. Mech Dev 73(1):23–32
Hidai H, Bardales R, Goodwin R, Quertermous T, Quertermous EE (1998) Cloning of capsulin, a basic helix-loop-helix factor expressed in progenitor cells of the pericardium and the coronary arteries. Mech Dev 73(1):33–43
Robb L, Mifsud L, Hartley L, Biben C, Copeland NG, Gilbert DJ, Jenkins NA, Harvey RP (2010) epicardin: a novel basic helix-loop-helix transcription factor gene expressed in epicardium, branchial arch myoblasts, and mesenchyme of developing lung, gut, kidney, and gonads. Dev Dyn 213(1):105–113
Familari M, Giraud AS (1998) Identification of the rat homologue of the mouse capsulin gene by cDNA representational difference analysis. Gene 222(2):245–248
Quaggin SE, Heuvel GBV, Igarashi P (1998) Pod-1, a mesoderm-specific basic-helix-loop-helix protein expressed in mesenchymal and glomerular epithelial cells in the developing kidney. Mech Dev 71(1-2):37–48
Yang Z, Jiang X, Li D, Dong Q, Zhao H, Jiang X (2019) TCF21 inhibits proliferation and chemoresistance through the AKT pathway in human gastric cancer. Gene 682:42–49
Zhang X, Cheng B, Liu C, Du Z, Zhang H, Wang N, Wu M, Li Y, Cao Z, Li H (2019) A novel regulator of preadipocyte differentiation, transcription factor TCF21, functions partially through promoting LPL expression. Front Physiol 10:458
Chen Y, Zhang C, Chen J, Zhang B, Zhang H, Yang X, Liu J, Wu Q (2018) Expression of transcription factor 21 (TCF21) and upregulation its level inhibits invasion and metastasis in esophageal squamous cell carcinoma. Med Sci Monit 24:4128–4136
Arab K, Smith LT, Gast A, Weichenhan D, Huang PH, Claus R, Hielscher T, Espinosa AV, Ringel MD, Morrison CD (2011) Epigenetic deregulation of TCF21 inhibits metastasis suppressor KISS1 in metastatic melanoma. Carcinogenesis 32(10):1467–1473
Zhang H, Na W, Zhang HL, Wang N, Du ZQ, Wang SZ, Wang ZP, Zhang Z, Li H (2017) TCF21 is related to testis growth and development in broiler chickens. Gen Select Evol Gse 49(1):25
Ide S, Finer G, Maezawa Y, Onay T, Souma T, Scott R, Ide K, Akimoto Y, Li C, Ye M (2018) Transcription factor 21 is required for branching morphogenesis and regulates the Gdnf-axis in kidney development. J Am Soc Nephrol 29(12):2795–2808
Cui S, Schwartz L, Quaggin SE (2010) Pod1 is required in stromal cells for glomerulogenesis. Dev Dyn 226(3):512–522
Cui S, Ross A, Stallings N, Parker KL, Capel B, Quaggin SE (2004) Disrupted gonadogenesis and male-to-female sex reversal in Pod1 knockout mice. Development 131(16):4095–4105
Lu J, Chang P, Richardson JA, Gan L, Weiler H, Olson EN (2000) The basic helix-loop-helix transcription factor capsulin controls spleen organogenesis. Proc Natl Acad Sci U S A 97(17):9525–9530
Quaggin SE, Schwartz L, Cui S, Igarashi P, Deimling J, Post M, Rossant J (1999) The basic-helix-loop-helix protein pod1 is critically important for kidney and lung organogenesis. Development 126(24):5771–5783
Arab K, Park YJ, Lindroth A, Schäfer A, Oakes C, Weichenhan D, Lukanova A, Lundin E, Risch A, Meister M (2014) Long noncoding RNA TARID directs demethylation and activation of the tumor suppressor TCF21 via GADD45A. Mol Cell 55(4):604–614
Panna T, Miteva YV, Kuchenbrod LM, Cristea IM, Conlon FL (2013) Tcf21 regulates the specification and maturation of proepicardial cells. Development 140(11):2409–2421
Zhao Q, Wirka R, Nguyen T, Nagao M, Cheng P, Miller CL, Kim JB, Pjanic M, Quertermous T (2019) TCF21 and AP-1 interact through epigenetic modifications to regulate coronary artery disease gene expression. Genome medicine 11(1):23
Maezawa Y, Onay T, Scott RP, Keir LS, Dimke H, Li C, Eremina V, Maezawa Y, Jeansson M, Shan J (2014) Loss of the podocyte-expressed transcription factor Tcf21/Pod1 results in podocyte differentiation defects and FSGS. J Am Soc Nephrol Jasn 25(11):2459–2470
Chen B, Zeng C, Ye Y, Wu D, Mu Z, Liu J, Xie Y, Wu H (2017) Promoter methylation of TCF21 may repress autophagy in the progression of lung cancer. J Cell Commun Sign 19(Suppl 17):1–10
Miller CL, D Ryan A, Kundu RK, Azad R, Nürnberg ST, Roxanne D, Karen C, Leeper NJ, Chung-Hsing C, I-Shou C (2013) Disease-related growth factor and embryonic signaling pathways modulate an enhancer of TCF21 expression at the 6q23.2 coronary heart disease locus. PLoS Genet 9(7):e1003652
Wu PL, Zhou Y, Zeng C, Li X, Dong ZT, Zhou YF, Bulun SE, Xue Q (2018) Transcription factor 21 regulates expression of ERβ and SF-1 via upstream stimulatory factor-2 in endometriotic tissues. Biochim Biophys Acta 1861(8):706–717
Li Q, Lei Y, Du W (2018) A novel target of p53, TCF21, can respond to hypoxia by MAPK pathway inactivation in uterine corpus endometrial carcinoma. DNA Cell Biol 37(5):473–480
Miyagishi M, Nakajima T, Fukamizu A (2000) Molecular characterization of mesoderm-restricted basic helix-loop-helix protein, POD-1/capsulin. Int J Mol Med 5(1):27–31
Cheol Yi H, Eun-Yeung G, Kabsun K, Ho SJ, Hyun-Mi K, Hyun Joo L, Hueng-Sik C, Keesook L (2005) Modulation of the expression and transactivation of androgen receptor by the basic helix-loop-helix transcription factor Pod-1 through recruitment of histone deacetylase 1. Mol Endocrinol 19(9):2245–2257
Natalia M, Cross JW, Christine S, Annette K, Kevin T, Rigby PWJ, Carvajal JJ (2012) Musculin and TCF21 coordinate the maintenance of myogenic regulatory factor expression levels during mouse craniofacial development. Development 139(5):958–967
Noriko F, Kimie O, Takayuki K, Masataka N (2003) Basic helix-loop-helix transcription factor epicardin/capsulin/Pod-1 suppresses differentiation by negative regulation of transcription. J Biol Chem 278(9):7486–7493
Jian-Rong L, Rhonda BD, April H, Priscilla C, Renee V, Hai W, Lin G, Shelton JM, Richardson JA, Olson EN (2002) Control of facial muscle development by MyoR and capsulin. Science 298(5602):2378–2381
Tamura M, Kanno Y, Chuma S, Saito T, Nakatsuji N (2001) Pod-1/Capsulin shows a sex- and stage-dependent expression pattern in the mouse gonad development and represses expression of Ad4BP/SF-1. Mech Dev 102(1):135–144
Bhandari RK, Sadlerriggleman I, Clement TM, Skinner MK (2011) basic helix-loop-helix transcription factor TCF21 is a downstreamtarget of the male sex determining gene SRY. PLoS One 6(5):e19935
Andrea B, Elisabetta F, Valentina S, Kelvin M, Susan Q, Francesco B, Cleary ML, Licia S (2005) A Pbx1-dependent genetic and transcriptional network regulates spleen ontogeny. Development 132(13):3113–3126
Duan HX, Li BW, Zhuang X, Wang LT, Cao Q, Tan LH, Qu GF, Xiao S (2019) TCF21 inhibits tumor-associated angiogenesis and suppresses the growth of cholangiocarcinoma by targeting PI3K/Akt and ERK signaling. Am J Physiol Gastrointest Liver Physiol 316(6):G763–G773
França MM, Ferraz-De-Souza B, Santos MG, Lerario AM, Latronico AC, Kuick RD, Hammer GD, Lotfi CFP (2013) POD-1 binding to the E-box sequence inhibits SF-1 and StAR expression in human adrenocortical tumor cells. Mol Cell Endocrinol 371(1-2):140–147
Akama T, Chun TH (2018) Transcription factor 21 (Tcf21) promotes proinflammatory interleukin 6 expression and extracellular matrix remodeling in visceral adipose stem cells. J Biol Chem 293(17):6603–6610
Bhandari RK, Schinke EN, Haque MM, Ingrid SR, Skinner MK (2012) SRY induced TCF21 genome-wide targets and cascade of bHLH factors during Sertoli cell differentiation and male sex determination in rats. Biol Reprod 87(6):131
Liu Y, Ding W, Ge H, Ponnusamy M, Wang Q, Hao X, Wu W, Zhang Y, Yu W, Ao X (2019) FOXK transcription factors: regulation and critical role in cancer. Cancer Lett 458:1–12
Xiong L, Wu F, Wu Q, Xu L, Cheung OK, Kang W, Mok MT, Szeto LLM, Lun CY, Lung RW, Zhang J, Yu KH, Lee SD, Huang G, Wang CM, Liu J, Yu Z, Yu DY, Chou JL, Huang WH, Feng B, Cheung YS, Lai PB, Tan P, Wong N, Chan MW, Huang TH, Yip KY, Cheng AS, To KF (2019) Aberrant enhancer hypomethylation contributes to hepatic carcinogenesis through global transcriptional reprogramming. Nat Commun 10(1):335
Francois G, Graeme JH, Amir E, Laurie JG, Jessica D, Gray JW, Heinrich L, Rudolf J (2003) Induction of tumors in mice by genomic hypomethylation. Science 300(5618):489–492
Smith LT, Mauting L, Brena RM, Lang JC, Schuller DE, Otterson GA, Morrison CD, Smiraglia DJ, Christoph P (2006) Epigenetic regulation of the tumor suppressor gene TCF21 on 6q23-q24 in lung and head and neck cancer. Proc Natl Acad Sci U S A 103(4):982–987
Daniel W, Christian S, Katrin SD, Claudia R (2013) Protein expression and promoter methylation of the candidate biomarker TCF21 in head and neck squamous cell carcinoma. Cell Oncol 36(3):213–224
Shivapurkar N, Stastny V, Xie Y, Prinsen C, Frenkel E, Czerniak B, Thunnissen FB, Minna JD, Gazdar AF (2008) Differential methylation of a short CpG-rich sequence within exon 1 of TCF21 gene: a promising cancer biomarker assay. Cancer Epidemiol Biomark Prev 17(4):995–1000
Shi YX, Wang Y, Li X, Zhang W, Zhou HH, Yin JY, Liu ZQ (2017) Genome-wide DNA methylation profiling reveals novel epigenetic signatures in squamous cell lung cancer. BMC Genomics 18(1):901
Richards KL, Baili Z, Menghong S, Wenli D, Jennifer C, Bachinski LL, Wilson CD, Baggerly KA, Guosheng Y, D Neil H (2011) Methylation of the candidate biomarker TCF21 is very frequent across a spectrum of early-stage nonsmall cell lung cancers. Cancer 117(3):606–617
Dai Y, Duan H, Duan C, Zhou R, He Y, Tu Q, Shen L (2016) Down-regulation of TCF21 by hypermethylation induces cell proliferation, migration and invasion in colorectal cancer. Biochem Biophys Res Commun 469(3):430–436
Xin J, Rong X, Lin S, Xin M, Cai W, Jin Z, Fu C, Zhen G, Lai J, Yue L (2016) Clinical potential ofTCF21methylation in the diagnosis of renal cell carcinoma. Oncol Lett 12(2):1265–1270
Gooskens SL, Klasson TD, Gremmels H, Logister I, Pieters R, Perlman EJ, Giles RH, Mm HE (2018) TCF21 hypermethylation regulates renal tumor cell clonogenic proliferation and migration. Mol Oncol 12(2):166–179
Gooskens SL, Gadd S, Auvil JMG, Gerhard DS, Khan J, Patidar R, Meerzaman D, Chen QR, Hsu CH, Yan C (2015) TCF21 hypermethylation in genetically quiescent clear cell sarcoma of the kidney. Oncotarget 6(18):15828–15841
Yang Z, Li DM, Xie Q, Dai DQ (2015) Protein expression and promoter methylation of the candidate biomarker TCF21 in gastric cancer. J Cancer Res Clin Oncol 141(2):211–220
Costa VL, Henrique R, Danielsen SA, Eknaes M, Patricio P, Morais A, Oliveira J, Lothe RA, Teixeira MR, Lind GE (2011) TCF21 and PCDH17 methylation: an innovative panel of biomarkers for a simultaneous detection of urological cancers. Epigenetics 6(9):1120–1130
Mrakovcic M, Kleinheinz J, Frohlich LF (2019) p53 at the crossroads between different types of HDAC inhibitor-mediated cancer cell death. Int J Mol Sci 20(10):2415
Ji L, Liu C, Yuan Y, Gao H, Tang ZX, Yang Z, Liu ZT, Jiang GH (2019) Key roles of Rho GTPases, YAP, and Mutant P53 in anti-neoplastic effects of statins. Fundam Clin Pharmacol 34(1):4–10
Yi Y, Zhang W, Yi J, Xiao ZX (2019) Role of p53 family proteins in metformin anti-cancer activities. J Cancer 10(11):2434–2442
She ZY, Yang WX (2017) Sry and SoxE genes: how they participate in mammalian sex determination and gonadal development? Semin Cell Dev Biol 63:13–22
Robles V, Valcarce DG, Riesco MF (2019) Non-coding RNA regulation in reproduction: their potential use as biomarkers. Non-coding RNA Res 4(2):54–62
Zhang W, Xu W, Feng Y, Zhou X (2019) Non-coding RNA involvement in the pathogenesis of diabetic cardiomyopathy. J Cell Mol Med 23(9):5859–5867
Wei J, Zhang L, Li J, Zhu S, Tai M, Mason CW, Chapman JA, Reynolds EA, Weiner CP, Zhou HH (2017) MicroRNA-205 promotes cell invasion by repressing TCF21 in human ovarian cancer. J Ovar Res 10(1):33
Chen D, Ma W, Ke Z, Xie F (2018) CircRNA hsa_circ_100395 regulates miR-1228/TCF21 pathway to inhibit lung cancer progression. Cell Cycle 17(16):2080–2090
Zhang H, Guo Y, Shang C, Song YS, Wu B (2012) miR-21 downregulated TCF21 to Inhibit KISS1 in renal cancer. Urology 80(6):1298–1302
Cao S, Jiang L, Shen L, Xiong Z (2019) Role of microRNA-92a in metastasis of osteosarcoma cells in vivo and in vitro by inhibiting expression of TCF21 with the transmission of bone marrow derived mesenchymal stem cells. Cancer Cell Int 19(1):31
Angelucci F, Cechova K, Valis M, Kuca K, Zhang B, Hort J (2019) MicroRNAs in Alzheimer’s disease: diagnostic markers or therapeutic agents? Front Pharmacol 10:665
Butz H, Patocs A (2019) MicroRNAs in endocrine tumors. Ejifcc 30(2):146–164
Cui J, Zhou B, Ross SA, Zempleni J (2017) Nutrition, microRNAs, and human health. Adv Nutr 8(1):105–112
Ying L, Xiang A, Wei D, Ponnusamy M, Wei W, Hao X, Yu W, Wang Y, Li P, Wang J (2018) Critical role of FOXO3a in carcinogenesis. Mol Cancer 17(1):104
Miller CL, Ulrike H, Roxanne D, Leeper NJ, Kundu RK, Bhagat P, Assimes TL, Kaiser FJ, Ljubica P, Ulf H (2014) Coronary heart disease-associated variation in TCF21 disrupts a miR-224 binding site and miRNA-mediated regulation. PLoS Genet 10(3):e1004263
Bastami M, Ghaderian SM, Omrani MD, Mirfakhraie R, Vakili H, Alipour PS, Nariman-Saleh-Fam Z, Masotti A (2016) MiRNA-related polymorphisms in miR-146a and TCF21 are associated with increased susceptibility to coronary artery disease in an Iranian population. Gen Test Mol Biomarkers 20(5):241–248
Sun W, Li S, Yu Y, Jin H, Xie Q, Hua X, Wang S, Tian Z, Zhang H, Jiang G, Huang C, Huang H (2019) MicroRNA-3648 is upregulated to suppress TCF21, resulting in promotion of invasion and metastasis of human bladder cancer. Mol Ther Nucleic Acids 16:519–530
Pamudurti NR, Bartok O, Jens M, Ashwalfluss R, Stottmeister C, Ruhe L, Hanan M, Wyler E, Perezhernandez D, Ramberger E (2017) Translation of CircRNAs. Mol Cell 66(1):9–21
Li M, Ding W, Tariq MA, Chang W, Zhang X, Xu W, Hou L, Wang Y, Wang J (2018) A circular transcript of ncx1 gene mediates ischemic myocardial injury by targeting miR-133a-3p. Theranostics 8(21):5855–5869
Zhang HD, Jiang LH, Sun DW, Hou JC, Ji ZL (2018) CircRNA: a novel type of biomarker for cancer. Breast Cancer 25(1):1–7
Haque S, Harries LW (2017) Circular RNAs (circRNAs) in health and disease. Genes (Basel) 8(12):353
Wang M, Yu F, Li P (2018) Circular RNAs: characteristics, function and clinical significance in hepatocellular carcinoma. Cancers 10(8):258
Xiong S, Peng H, Ding X, Wang X, Wang L, Wu C, Wang S, Xu H, Liu Y (2019) Circular RNA expression profiling and the potential role of hsa_circ_0089172 in Hashimoto’s thyroiditis via sponging miR125a-3p. Mol Ther Nucleic Acids 17:38–48
Zhang L, Song X, Chen X, Wang Q, Zheng X, Wu C, Jiang J (2019) Circular RNA CircCACTIN promotes gastric cancer progression by sponging MiR-331-3p and regulating TGFBR1 expression. Int J Biol Sci 15(5):1091–1103
Ignarski M, Islam R, Muller RU (2019) Long non-coding RNAs in kidney disease. Int J Mol Sci 20(13):3276
Xu J, Bai J, Zhang X, Lv Y, Gong Y, Liu L, Zhao H, Yu F, Ping Y, Zhang G, Lan Y, Xiao Y, Li X (2017) A comprehensive overview of lncRNA annotation resources. Brief Bioinform 18(2):236–249
Wu XS, Wang F, Li HF, Hu YP, Jiang L, Zhang F, Li ML, Wang XA, Jin YP, Zhang YJ (2017) LncRNA-PAGBC acts as a microRNA sponge and promotes gallbladder tumorigenesis. EMBO Rep 18(10):1837–1853
Bayoumi AS, Sayed A, Broskova Z, Teoh JP, Wilson J, Su H, Tang YL, Kim IM (2016) Crosstalk between long noncoding RNAs and microRNAs in health and disease. Int J Mol Sci 17(3):356
Abbastabar M, Sarfi M, Golestani A, Khalili E (2018) lncRNA involvement in hepatocellular carcinoma metastasis and prognosis. EXCLI J 17:900–913
Li W, Notani D, Qi M, Tanasa B, Nunez E, Chen AY, Merkurjev D, Jie Z, Ohgi K, Song X (2013) Functional importance of eRNAs for estrogen-dependent transcriptional activation events. Nature 498(7455):516–520
Guohui W, Yunhua L, Cecil H, Xinna Z, Xiongbin L (2014) Noncoding RNAs in DNA repair and genome integrity. Antioxid Redox Signal 20(4):655–677
Rinn JL, Chang HY (2012) Genome regulation by long noncoding RNAs. Annu Rev Biochem 81:145–166
Guo X, Wei Y, Wang Z, Liu W, Yang Y, Yu X, He J (2018) LncRNA LINC00163 upregulation suppresses lung cancer development though transcriptionally increasing TCF21 expression. Am J Cancer Res 8(12):2494–2506
Liu Y, Ao X, Jia Z, Bai XY, Xu Z, Hu G, Jiang X, Chen M, Wu H (2015) FOXK2 transcription factor suppresses ERα-positive breast cancer cell growth through down-regulating the stability of ERα. Sci Rep 5(4):8796
Yan K, Wang K, Li P (2019) The role of post-translational modifications in cardiac hypertrophy. J Cell Mol Med 23(6):3795–3807
Krentz NAJ, Hoof DV, Li Z, Watanabe A, Tang M, Nian C, German MS, Lynn FC (2017) Phosphorylation of NEUROG3 links endocrine differentiation to the cell cycle in pancreatic progenitors. Dev Cell 41(2):129–142
Singh V, Ram M, Kumar R, Prasad R, Roy BK, Singh KK (2017) Phosphorylation: implications in cancer. Protein J 36(1):1–6
Anett M, Zongling J, Child ES, Eberhard K, Mann DJ, Sharrocks AD (2010) Cell cycle-dependent regulation of the forkhead transcription factor FOXK2 by CDK·cyclin complexes. J Biol Chem 285(46):35728–35739
Lee EW, Oh W, Song HP, Kim WK (2017) Phosphorylation of p53 at threonine 155 is required for Jab1-mediated nuclear export of p53. BMB Rep 50(7):373–378
Moller AB, Vendelbo MH, Schjerling P, Couppe C, Moller N, Kjaer M, Hansen M, Jessen N (2019) Immobilization decreases FOXO3a phosphorylation and increases autophagy-related gene and protein expression in human skeletal muscle. Front Physiol 10:736
Yang Y, He Y, Wang X, Liang Z, He G, Zhang P, Zhu H, Xu N, Liang S (2017) Protein SUMOylation modification and its associations with disease. Open Biol 7(10):170167
Xiang A, Li S, Xu Z, Yang Y, Chen M, Jiang X, Wu H (2016) Sumoylation of TCF21 downregulates the transcriptional activity of estrogen receptor-alpha. Oncotarget 7(18):26220–26234
Iyer D, Zhao Q, Wirka R, Naravane A, Nguyen T, Liu B, Nagao M, Cheng P, Miller CL, Kim JB (2017) Coronary artery disease genes SMAD3 and TCF21 promote opposing interactive genetic programs that regulate smooth muscle cell differentiation and disease risk. PLoS Genet 14(10):e1007681
Olga S, Yuqi Z, Sylvia N, Clint M, Milos P, Castano VG, Kim JB, Salfati EL, Kundaje AB, Gill B (2015) Characterization of TCF21 downstream target regions identifies a transcriptional network linking multiple independent coronary artery disease loci. PLoS Genet 11(5):e1005202
Fujimaki T, Oguri M, Horibe H, Kato K, Matsuoka R, Abe S, Tokoro F, Arai M, Noda T, Watanabe S (2015) Association of a transcription factor 21 gene polymorphism with hypertension. Biomedical Reports 3(1):118–122
Yang L, Gao X, Luo H, Huang Q, Su D, Tan X, Lu C (2017) TCF21 rs12190287 Polymorphisms are associated with ventricular septal defects in a Chinese population. Gen Test Mol Biomarkers 21(5):312–315
Mehedintu C, Plotogea MN, Ionescu S, Antonovici M (2014) Endometriosis still a challenge. J Med Life 7(3):349–357
Xiao Z, Rodriguez PQ, He L, Betsholtz C, Tryggvason K, Patrakka J (2013) Wtip- and gadd45a-interacting protein dendrin is not crucial for the development or maintenance of the glomerular filtration barrier. PLoS One 8(12):e83133
Nagata M (2016) Podocyte injury and its consequences. Kidney Int 89(6):1221–1230
Wang J, Gao X, Wang M, Zhang J (2015) Clinicopathological significance and biological role of TCF21 mRNA in breast cancer. Tumor Biol 36(11):1–5
Gao X, Yang J, Wang M, Zhang J (2016) TCF21 genetic polymorphisms and breast cancer risk in Chinese women. Oncotarget 7(34):55757–55764
Wu H, Zhou J, Zeng C, Wu D, Mu Z, Chen B, Xie Y, Ye Y, Liu J (2016) Curcumin increases exosomal TCF21 thus suppressing exosome-induced lung cancer. Oncotarget 7(52):87081–87090
Xiao J, Liu A, Lu X, Chen X, Li W, He S, He B, Chen Q (2017) Prognostic significance of TCF21 mRNA expression in patients with lung adenocarcinoma. Sci Rep 7(1):2027
Franca MM, Ferrazdesouza B, Lerario AM, Lotfi CFP (2015) POD-1/TCF21 reduces SHP expression, affecting LRH-1 regulation and cell cycle balance in adrenocortical and hepatocarcinoma tumor cells. Biomed Res Int 2015(17):1–9
Dai Y, Duan H, Duan C, Zhu H, Zhou R, Pei H, Shen L (2017) TCF21 functions as a tumor suppressor in colorectal cancer through inactivation of PI3K/AKT signaling. Oncotargets & Therapy 10:1603–1611
Ye YW, Jiang ZM, Li WH, Li ZS, Han YH, Sun L, Wang Y, Xie J, Liu YC, Zhao J (2012) Down-regulation of TCF21 is associated with poor survival in clear cell renal cell carcinoma. Neoplasma 59(6):599–605
Vyskocil E, Pammer J, Altorjai G, Grasl MC, Parzefall T, Haymerle G, Janik S, Perisanidis C, Erovic BM (2019) Dysregulation of ß-catenin, WISP1 and TCF21 predicts disease-specific survival and primary response against radio(chemo)therapy in patients with locally advanced squamous cell carcinomas of the head and neck. Clin Otolaryngol (Blackwell) 44(3):263–272
Passaia BDS, Dias MH, Kremer JL, Antonini SRR, Almeida MQD, Lotfi CFP (2018) TCF21/POD-1, a transcritional regulator of SF-1/NR5A1, as a potential prognosis marker in adult and pediatric adrenocortical tumors. Front Endocrinol 9:38–52
França MM, Lerario AM, Fragoso MCBV, Lotfi CFP, França MM, Lerario AM, Fragoso MCBV, Lotfi CFP (2017) New evidences on the regulation of SF-1 expression by POD1/TCF21in adrenocortical tumor cells. Clinics 72(6):391–394
Hoxhaj G, Manning BD (2020) The PI3K-AKT network at the interface of oncogenic signalling and cancer metabolism. Nat Rev Cancer 20(2):74–88
Sun Y, Liu WZ, Liu T, Feng X, Yang N, Zhou HF (2015) Signaling pathway of MAPK/ERK in cell proliferation, differentiation, migration, senescence and apoptosis. J Recept Signal Transduct Res 35(6):600–604
Peng WX, Huang JG, Yang L, Gong AH, Mo YY (2017) Linc-RoR promotes MAPK/ERK signaling and confers estrogen-independent growth of breast cancer. Mol Cancer 16(1):161
Iida M, Tsuboi K, Niwa T, Ishida T, Hayashi SI (2019) Compensatory role of insulin-like growth factor 1 receptor in estrogen receptor signaling pathway and possible therapeutic target for hormone therapy-resistant breast cancer. Breast Cancer 26(3):272–281
Evan GI, Vousden KH (2001) Proliferation, cell cycle and apoptosis in cancer. Nature 411(6835):342–348
Tan J, Zhang G, Liu R, Zhou M, Li Z, Wu Z (2015) Over-expression of transcription factor 21 inhibits the proliferation and migration and promotes apoptosis of SMMC-7721 cells. Chin J Cell Mol Immunol 31(7):884–888
John RR, Malathi N, Ravindran C, Anandan S (2017) Mini review: multifaceted role played by cyclin D1 in tumor behavior. Indian J Dent Res 28(2):187–192
Kanska J, Zakhour M, Taylor-Harding B, Karlan BY, Wiedemeyer WR (2016) Cyclin E as a potential therapeutic target in high grade serous ovarian cancer. Gynecol Oncol 143(1):152–158
Zanotelli MR, Reinhart-King CA (2018) Mechanical forces in tumor angiogenesis. Adv Exp Med Biol 1092:91–112
Chen X, Man G, Liu Y, Wu F, Huang J, Li TC, Wang CC (2017) Physiological and pathological angiogenesis in endometrium at the time of embryo implantation. Am J Reprod Immunol 78(Suppl l):e12693
Hongu T, Yamauchi Y, Funakoshi Y, Katagiri N, Ohbayashi N, Kanaho Y (2016) Pathological functions of the small GTPase Arf6 in cancer progression: tumor angiogenesis and metastasis. Small Gtpases 7(2):47–53
Lin B, Song X, Yang D, Bai D, Yao Y, Lu N (2018) Anlotinib inhibits angiogenesis via suppressing the activation of VEGFR2, PDGFRβ and FGFR1. Gene 654:77–86
Beth L, Guido K (2008) Autophagy in the pathogenesis of disease. Cell 132(1):27–42
Guo JY, White E (2017) Autophagy, metabolism, and cancer. Cold Spring Harb Symp Quant Biol 21(22):5037–5046
Samy L, Jian X, Rik D (2014) Molecular mechanisms of epithelial-mesenchymal transition. Nat Rev Mol Cell Biol 15(3):178–196
Gonzalez DM, Damian M (2014) Signaling mechanisms of the epithelial-mesenchymal transition. Sci Signal 7(344):re8
Liu Y (2019) Targeting the non-canonical AKT-FOXO3a axis: a potential therapeutic strategy for oral squamous cell carcinoma. EBioMedicine 49:6–8
Stracke ML, Liotta LA (1992) Multi-step cascade of tumor cell metastasis. Vivo 6(4):309–316
Steeg PS (2016) Targeting metastasis. Nat Rev Cancer 16(4):201–218
Gerashchenko TS, Novikov NM, Krakhmal NV, Zolotaryova SY, Zavyalova MV, Cherdyntseva NV, Denisov EV (2019) Markers of cancer cell invasion: are they good enough? J Clin Med 8(8):1092
Lu W, Yang C, Du P, Zhang JL, Zhang JC (2019) Expression tendency and prognostic value of TCF21 in hepatocellular carcinoma. Artificial Cell Nanomed Biotechnol 47(1):1466–1470
Shin B, Feser R, Nault B, Hunter S, Maiti S, Ugwuagbo KC, Majumder M (2019) miR526b and miR655 induce oxidative stress in breast cancer. Int J Mol Sci 20(16):4039
Funding
This work was supported by the National Natural Science Foundation of China (81702785, 81802822), Natural Science Foundation of Shandong Province (ZR2017PH013, ZR2018BH017), and Qingdao Postdoctoral Application Research Funded Project (2016067).
Author information
Authors and Affiliations
Contributions
WD, YZ, and DD collected the related paper. XA and YL drafted and wrote the manuscript. YL designed and revised the manuscript. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no conflict of interest.
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
About this article

Cite this article
Ao, X., Ding, W., Zhang, Y. et al. TCF21: a critical transcription factor in health and cancer. J Mol Med 98, 1055–1068 (2020). https://doi.org/10.1007/s00109-020-01934-7
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00109-020-01934-7