
Abstract
We experimentally imposed three different kinds of warming scenarios over 3 years on an alpine meadow community to identify the differential effects of climate warming and extreme climatic events on the abundance and biomass of bryophytes and lichens. Treatments consisted of (a) a constant level of warming with open top chambers (an average temperature increase of 1.87 °C), (b) a yearly stepwise increase of warming (average temperature increases of 1.0; 1.87 and 3.54 °C, consecutively), and (c) a pulse warming, i.e., a single first year pulse event of warming (average temperature increase of 3.54 °C only during the first year). To our knowledge, this is the first climate change study that attempts to distinguish between the effects of constant, stepwise and pulse warming on bryophyte and lichen communities. We hypothesised that pulse warming would have a significant short-term effect compared to the other warming treatments, and that stepwise warming would have a significant mid-term effect compared to the other warming treatments. Acrocarpous bryophytes as a group increased in abundance and biomass to the short-term effect of pulse warming. We found no significant effects of mid-term (third-year) stepwise warming. However, one pleurocarpous bryophyte species, Tomentypnum nitens, generally increased in abundance during the warm year 1997 but decreased in control plots and in response to the stepwise warming treatment. Three years of experimental warming (all treatments as a group) did have a significant impact at the community level, yet changes in abundance did not translate into significant changes in the dominance hierarchies at the functional level (for acrocarpous bryophytes, pleurocarpous bryophytes, Sphagnum or lichens), or in significant changes in other bryophyte or lichen species. The results suggest that bryophytes and lichens, both at the functional group and species level, to a large extent are resistant to the different climate change warming simulations that were applied.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Global climate forecasts include an increase in both the frequency and the magnitude of climatic events; for example, heat waves, drought, heavy rainfall and floods are all predicted to occur more frequently (IPCC 2007). These climate events will likely have different impacts on singular organisms and whole ecosystems (Jentsc and Beierkuhnlein 2008). For bryophytes, heat waves have been shown to trigger the die-off of peat mosses of the genus Sphagnum (Bragazza 2008). Experimentally imposed winter warming was shown to have a negative impact on the net photosynthetic rate of Hylocomium splendens, whereas the lichen Peltigera aphthosa was unaffected by the same experimental treatment (Bjerke et al. 2011).
Polar and alpine ecosystems are assumed to be particularly vulnerable to climate change, as organisms in these environments live at temperatures just above the zero-degree threshold during the very short summer season. This has drawn the attention of the climate change research community that has attempted to assess the impact of climate warming on tundra plants and plant communities in a number of experiments. For logistical reasons, many of these climate change experiments involve passive warming devices. Open-top chambers (OTCs) are one of the most commonly used devices in polar and alpine areas to assess warming effects on terrestrial ecosystems where durable, low-maintenance experimental devices are especially suitable (Alatalo 1998; Arft et al. 1999; Walker et al. 2006). OTCs have been used to simulate above-ground climate change effects on single species of vascular plants (Klanderud 2008; Alatalo and Little 2014), bryophytes (e.g., Molau and Alatalo 1998; Jägerbrand et al. 2003, 2006, 2009), lichens (e.g., Alatalo 1998; Molau and Alatalo 1998; Cornelissen et al. 2004), functional groups (Dormann and Woodin 2002) and entire plant communities (Alatalo 1998). Similarly, OTCs have also been used to simulate climate change effects on subsurface bacteria (Rinnan et al. 2009), fungi (Olsrud et al. 2004) and arthropods (e.g., Bokhorst et al. 2007; Hågvar and Klanderud 2009; Makkonen et al. 2011). However, vascular plants have been the focus of the majority of experimental climate change studies in the Arctic (e.g., Alatalo and Totland 1997; Arft et al. 1999; Dorji et al. 2013).
Although bryophytes and lichens are often overlooked in ecological studies (Jägerbrand et al. 2009), these groups play an important role in Arctic and subarctic vegetation communities, and their relative influence on cover, biomass and nutrient cycling tends to increase at higher latitudes (Longton 1984). Bryophytes in particular have been shown to influence the recruitment of vascular plants (e.g., Soudzilovskaia et al. 2011) and affect permafrost stability (Harden et al. 2006; Romanovsky et al. 2010; Turetsky et al. 2012).
When bryophytes and lichens are studied by climate change experiments, they are typically included as functional types (e.g., Graglia et al. 2001; Van Wijk et al. 2003; Elmendorf et al. 2012) or a limited number of species (e.g., Potter et al. 1995; Jägerbrand et al. 2003, 2012; Bjerke et al. 2011). Only a small number of studies have been conducted on bryophytes and lichens at the species level for whole communities (e.g., Jägerbrand et al. 2006, 2009; Klanderud 2008; Lang et al. 2012).
In most cases, experimental configurations that simulate climate change using OTCs have not allowed for a differentiation between the effects of rare extreme climatic events, a stepwise increase in warming or a stable warming enhancement (but see Bjerke et al. 2011). It is unknown whether the impact of a singular climate event may differ from a general warming trend, such as the effects simulated in most warming experiments, or successively increased warming. Thus, there is a need for studies that can distinguish between the impacts of singular warming events and more general warming. This study aimed to (a) distinguish between the impact of a constant (i.e., standard OTC treatment, a stable increased temperature; Marion et al. 1997), pulse (a 1-year event of high-impact warming simulating an extreme climatic event) and stepwise (a warming that is successively raised among years) temperature change on bryophyte and lichen communities and to (b) examine the extent to which bryophytes and lichens are responsive or resilient to different warming experiments. To better understand the differences in community responses and to evaluate these responses in comparison with previous OTC studies, it was necessary to study the bryophyte and lichen responses at three different hierarchical levels. Thus, we examined the effects of the warming treatments on the community composition, abundance and biomass at both the functional group and individual species level.
The following questions were addressed: (1) are responses to standard OTC warming similar to the responses to stepwise and pulse warming?, (2) are responses to stepwise and pulse warming significantly different?, and (3) do bryophytes and lichens show resilience, or do they respond to the experimental warming treatments? Specifically, we hypothesised that pulse warming would have a significant short-term effect compared to the other warming treatments, and stepwise warming would have a significant mid-term effect compared to the other warming treatments.
Methods
Study area
Fieldwork was conducted in northern Sweden at the Latnjajaure Field Station (LFS) in the valley of Latnjavagge, 68°21′N, 18°29′E, at an elevation of 1,000 m. Since the early spring of 1992, a year-round automatic station has provided a continuous climate data set.
The valley is covered by snow for most of the year, and the climate is characterised by cool summers and relatively mild, snow-rich winters (annual minimum temperature ranging from −27.3 to −21.7 °C). The site has a mean annual temperature of −2.0 to −2.7 °C (data from 1993 to 1999) and is classified as subarctic (Polunin 1951; Alatalo and Molau 1995). The annual precipitation ranges from 605 mm (1996) to 990 mm (1993), and the mean for 1990–1999 was 808 mm. July is the warmest month, with a mean temperature ranging from +5.2 °C (1995) to +8.0 °C (1997). Climate data for the study years 1995–1998 are provided in Table 1.
The vegetation in the valley comprises a wide range of communities, and the environment varies from dry to wet and from acidic to base-rich. Although the geographical environment is subarctic-alpine, the vegetation of the area is representative of the Low Arctic, and Cassiope tetragona, Dryas octopetala and Carex bigelowii are among the dominant species (Molau and Alatalo 1998).
Experimental design
The experiment was performed in a rich meadow community (see Alatalo 1998; Molau and Alatalo 1998 for detailed description) ca. 300-m southeast of the LFS on a gentle northwest-facing slope with an abundant ground water supply. In July 1995, four blocks, each with four 1 × 1 m plots, were marked and numbered. All plots were as similar as possible with respect to the floristic composition and edaphic conditions.
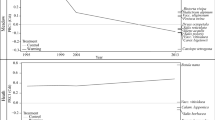
At the end of the 1995 season, the subsequent treatments were distributed within the blocks using a simple lottery. Within each of the four blocks, four different treatments were implemented starting in June 1996. The treatments included (1) control, (2) standard OTC warming, (3) stepwise warming and (4) pulse warming. The OTC experiments (2), configured according to Marion et al. (1997), used hexagonal polycarbonate chambers with a base diameter of 1 m (Marion et al. 1997; Molau 2001; Molau and Alatalo 1998) fixed to the ground from early June 1996 to late August 1998. In the stepwise manipulation (3), an OTC was installed in the plot on 10 cm-high pylons throughout the 1996 season; the OTC was affixed to the ground during the 1997 season, and a polyethylene lid was used throughout the 1998 season, thus increasing the experimental warming each year. In the pulse treatment (4), a closed-top chamber [CTC; a standard OTC with a polyethylene lid as in (3)] was installed in the plot during the 1996 season only and was removed in late August of the same year. Closed top chambers have commonly been used in agricultural research for studies on methane emissions (Knapp and Yavitt 1992), CO2 and H2O fluxes (Dugas et al. 1997; Steduto et al. 2002), photosynthesis and evapotranspiration (Reicosky 1990; Nie et al. 1992). A general overview of the warming treatments is presented in Fig. 1.

Schematic overview of the warming treatments. Each step above the baseline corresponds to ca. 1 °C. Treatments: control (Control), constant warming enhancement with open-top chambers (OTC), a stepwise increase in the magnitude of warming (Step) and a single-summer high-impact warming event (Pulse)
The experiment was specifically developed for this present study, and the plots were not previously studied in any project. The experiment was designed using the BACI (before-after-control-impact) approach, and all plots were mapped in the first year (1995) prior to any manipulation (Underwood 1994). The experiment was ended after the final data collection period in August 1998.
Measurements
All 16 plots were mapped in early August of each year (1995–1998) in the same sequence such that each individual plot was mapped on approximately the same date every year. A 1 × 1 m grid frame (Walker 1996) was used to map the plots; in each of the grid points, the specific identities of the bottom-layer species were noted. A single person, experienced field botanist Vivian Aldén, performed the plot mapping in all 4 years. The bryophyte collection from the point framing in 1995 was determined to the species level by Sven Fransén, Göteborg University. The discovery of the bryophyte Campylopus schimperi (det. S. Fransén) in one of the plots was one of the first records of that species in Sweden; a complete list of the bryophyte and lichen species found in the experimental plots is provided in the species list (Table S1). The nomenclature of Hallingbäck et al. (2006) was used for the bryophytes, and that of Moberg and Holmåsen (1990) was used for the lichens. In the 1 × 1 m control plots, 96 sampling points were consistently used for the bottom layer (four points were sacrificed for orientation screws (5-mm diameter), enabling the proper re-installation of the grid frame each year; Walker 1996). Due to their hexagonal shape, the OTCs reduced the number of points per plot to 87–94. Solifluction at the study site was very low, totalling less than 1 cm of the horizontal distance over the 4 years of the study. The biomass of the various functional groups was estimated using the life form-specific algorithms established for the site (Table 2, after Karlsson 1998; Molau 2010).
The air temperature at the surface of the soil in some of the manipulated plots (always in comparison with parallel control plots) was measured using Tinytag™ temperature loggers; the loggers recorded at 30 min intervals, and mean temperatures were calculated using series comprising 1000–5600 timed readings.
Data analysis
To investigate whether the treatments influenced the abundance and composition of the communities of bryophytes and lichens, we used a multivariate gradient analysis that incorporated environmental (i.e., treatment) variables as constrained factors to optimise the fit of the data. Because it was unknown whether the underlying responses were linear or non-linear, we performed both constrained canonical correspondence analysis (CCA) and redundancy analysis (RDA), and the first three axes explained 29.53 and 49.77 %, respectively, of the cumulative proportion of variance. Thus, we chose to use RDA and performed a full RDA that included the treatments, years and blocks as constrained factors followed by three partial RDA tests to calculate the variance explained by each constraining factor. The significance of the constraining factors was analysed in the full RDA using an ANOVA-like permutation test (Oksanen et al. 2013). Multivariate analysis was performed on non-transformed species data with the default settings. The abundance and biomass were log-transformed prior to the univariate, parametric analyses to meet normality assumptions. ANOVA tests were performed to analyse the significant effects of years, blocks and treatments on the abundance and biomass of functional groups and individual species. On individual species level, the analyses were conducted on the ten most common bryophyte species and the four most common lichen species. Species with less than 100 hits from the point framing from all 4 years were excluded from the species specific analysis, thus the low number of lichen species in the species specific analysis.
To investigate the responses in abundance and biomass for each treatment and during all 4 years of the experiment, we conducted repeated ANOVA tests with the treatment and block as well as their interactions as independent factors and the year as the repeated factor. However, the abundance of the species Anthelia juratzkana, Dicranum elongatum, Polytrichum strictum, Sphaerophorus globosus and Tomentypnum nitens could not be transformed and was therefore tested using the non-parametric Friedman’s test in SPSS. Significant effects were subsequently analysed by generalised linear models for poisson distributed data with a loglinear link function. Effects of year, treatment and their interactions were tested for best fit in the GLM with AIC, Akaike information criterion.
To test for an effect of the first-year pulse on relative changes in the abundance and biomass, we conducted a univariate ANOVA with the treatment as the fixed factor and the block as the random factor. Power analysis was used to compute the effect size and required sample size for abundance, number of species and biomass, since the number of replicates combined with the short-term time-period may have been restraining responses. Power analysis settings were f-test ANOVA repeated measures.
Repeated ANOVA tests were conducted using R version 2.15.2, and multivariate analyses were performed using the R package vegan 2.0-7 (Oksanen et al. 2013). Univariate ANOVA tests, Friedman’s tests and generalised linear models were conducted using SPSS Statistics software (IBM, version 21). Power analysis was conducted in G*Power 3.1.9.2 (Faul et al. 2009).
Results
Effect of treatments on temperature regime
Although the climate differed among the study years, the warming of the air temperature at the surface of the soil provided by the standard OTC remained relatively constant at an average of 1.87 ± 0.25 °C (mean ± SE, n = 7) above the ambient temperature (i.e., the air temperature at the surface of the soil in adjacent control plots). The ventilated OTCs in the first treatment year in the stepwise simulation created a warming of 1.00 ± 0.42 °C (n = 2), and the CTC treatment in the stepwise (year 3) and pulse experiments created an increase of 3.54 ± 0.24 °C (n = 3) above the control plots. The surface temperature of the reference control plots averaged 9.25 ± 0.55 °C during the study seasons. Thus, the experimental warming treatments can be classified into three temperature equivalents of ca. 1 °C each (see Fig. 1): the cumulative sum after the entire experiment was equal for the OTC and stepwise treatments with a total of six units, whereas the pulse treatment received only three units above the control, although in one single season.
Community-level responses
There were only a few species that varied substantially with the RDA axes, while the species composition of the plots in the RDA were more prone to be clustered together, see Fig. 2. The first two axes of the constrained RDA for the bryophyte and lichen communities explained 41.2 % of the variation, and the ANOVA-like permutation test showed that the treatments, years and blocks had a significant influence (Table 3; Fig. 2). The full and partial RDA showed that the treatments (standard OTC, pulse and stepwise) and blocks explained the largest proportion of the inertia (Table 4).

Plot of the constrained redundancy analysis (RDA) of the change in abundance in bryophyte and lichen communities at Latnjajaure Field Station, subarctic Sweden, after 3 years of experimental treatment. RDA 1 and 2 explain 41.2 % of the variation. Constrained variables shown in the plot (blue): OTC constant warming enhancement with open-top chambers, PU a short-term high-impact warming (Pulse), C controls, block B1, B2, B3, B4. Species abbreviations are listed in Table 2. Each row represents one plot. Species and site scores are unscaled (scaling 0) (colour figure online)
Functional group-level responses
As predicted, the pulse warming had a significant effect on relative change of abundance and biomass (univariate ANOVA, p = 0.003), however, this effect was limited to acrocarpous bryophytes, which showed a positive response in terms of the relative change in abundance and biomass in the first year. Pleurocarpous bryophytes, Sphagnum and lichens did not show any significant response to the pulse warming (Fig. 3). The 3 years of constant or stepwise warming did not result in any significant effects on the functional group level of acrocarpous, pleurocarpous bryophytes (including liverworts), Sphagnum or lichens when evaluated by repeated ANOVA tests on abundance and biomass (Figs. 4, 5). Thus, we found no support for the hypothesised third-year effect of stepwise warming on functional groups.

Boxplots of the relative changes (1995–1996) in abundance and biomass at the functional group level with different treatments at Latnjajaure Field Station: Acro acrocarpous bryophytes, Lich lichens, Pleu pleurocarpous bryophytes, Spha Sphagnum spp. Treatments: control (Control), constant warming enhancement with open-top chambers (OTC), a stepwise increase in the magnitude of warming (Step) and a single-summer high-impact warming event (Pulse). Boxplots show the 10th to 90th percentile of the data; n = 4 plots per treatment

Boxplots of the abundance at the functional group level after different treatments and years at Latnjajaure Field Station: a acrocarpous bryophytes, b pleurocarpous bryophytes, c Sphagnum spp. and d Lichens. Treatments: control (Control), constant warming enhancement using open-top chambers (OTC), a stepwise increase in the magnitude of warming (Step) and a single-summer high-impact warming event (Pulse). Boxplots show the 10th–90th percentile of the data; n = 4 plots per treatment

Boxplots of the biomass (dry mass g m−2) after different warming treatments and years at Latnjajaure Field Station: a acrocarpous bryophytes, b pleurocarpous bryophytes, c Sphagnum spp. and d Lichens. Treatments: control (Control), constant warming enhancement using open-top chambers (OTC), a stepwise increase in the magnitude of warming (Step) and a single-summer high-impact warming event (Pulse). Boxplots show the 10th–90th percentile of the data; n = 4 plots per treatment
Individual species-level responses
Of the ten most common bryophyte species recorded in the constant, stepwise and pulse warming plots, we only found significant effects on the abundance of T. nitens (Fig. 6) (treatment and years, Friedman’s test, p = 0.047). The generalised linear model of T. nitens with lowest AIC included year and treatment as single explanatory factors. Results show that the abundance of T. nitens increased in 1997 (parameter estimates: B = 0.26; df = 1, p = 0.03), decreased in control plots (B = −0.68; df = 1, p < 0.0001) and decreased in response to the stepwise warming treatment (B = −1.6; df = 1, p < 0.0001). Other years and treatment were not significant (not presented).

Boxplots of abundance on Tomentypnum nitens after different warming treatments and years at Latnjajaure. Treatments: control (Control), constant warming enhancement using open-top chambers (OTC), a stepwise increase in the magnitude of warming (Step) and a single-summer high-impact warming event (Pulse). Boxplots show the 10th–90th percentile of the data; n = 4 plots per treatment
No significant effects due to constant, stepwise and pulse warming were observed with respect to the four most common lichen species that were analysed. Thus, at the species level, most bryophyte and lichen species exhibited strong resilience to the short-term warming treatments (Figs. S1, S2).
Power analysis
Effect size (f) was small; 0.077, 0.085 and 0.0685, respectively for cover, number of species and biomass. Computed required total sample sizes were 1,544 for abundance, 1,264 for the number of species and 1,952 for biomass. The low effect size indicated that we had low probability to detect significant effects if present.
Discussion
To our knowledge, this is the first climate change study that attempted to distinguish between the effects of constant, stepwise and pulse warming on bryophyte and lichen communities. The 3 years of environmental warming (all treatments as a group) had a significant effect at the community level. However, this effect did not translate into significant changes in the dominance hierarchies at the functional type level (acrocarpous and pleurocarpous bryophytes, Sphagnum and lichens) or at the species level, except for in one species, T. nitens. This finding is in general agreement with modelling results suggesting that northern ecosystems must experience extreme warming before mosses will disappear (Turetsky et al. 2012).
However, a literature review on responses to different perturbations (e.g., fire, warming, nutrient addition and drought) in different ecosystems (e.g., tundra, meadow, forest and peatland) reported that bryophytes declined in 40 % of studies in Arctic ecosystems (Turetsky et al. 2012). Studies on bryophytes and lichens have reported contrasting effects, and bryophytes and lichens have been shown to have highly site-specific responses to warming experiments, with responses varying at the local scale among different vegetation types (e.g., Jägerbrand et al. 2009) or among larger geographical scales (e.g., Lang et al. 2012; Turetsky et al. 2012). The responses to constant warming applied using OTCs have varied, including no effect on the abundance of bryophytes (Chapin et al. 1995; Alatalo 1998; Jägerbrand et al. 2003, 2009, 2012), a negative effect on bryophytes (Press et al. 1998), no effect on lichens (Jägerbrand et al. 2009), a negative effect on lichens (Press et al. 1998; Lang et al. 2012) and positive effects on lichens (Chapin et al. 1995; Alatalo 1998; Biasi et al. 2008; Jägerbrand et al. 2009). It has been hypothesised that these contrasting responses to treatments are dependent on the level of competition between bryophytes and lichens and vascular plants as well as the extent to which they can thrive under competition for light (Alatalo 1998). In sites with existing dense canopies of vascular plants, the bottom layer is dominated by shade-tolerant species, whereas in sites with less-developed vascular plant canopies, the bottom layer is dominated by shade-intolerant species, which are more likely to be negatively affected if the competition for light increases (Alatalo 1998). This hypothesis was supported by a study on the effect of neighbouring vascular plants on the abundance of bryophytes in different vegetation types showing that the relationship between bryophytes and different functional types of vascular plants differed at the local scale among vegetation types (Jägerbrand et al. 2012). Specifically, they found that the abundance of bryophytes was negatively correlated with the abundance of deciduous shrubs, evergreen shrubs and lichens under natural conditions in heath community; however, no such relationship was found in the meadow, suggesting that bryophyte species in the meadow with dense canopy were less sensitive to light competition than the bryophyte species found in the heath community with more sparse canopy of vascular plants (Jägerbrand et al. 2012). While we have not analysed the vascular plant data from this study (the vascular plant canopy cover ranged 63 and 98 %), this study was conducted in a similar meadow as the study by Jägerbrand et al. (2012), but with different perturbations. In our study, the lack of a response in abundance at the functional type and species levels may also have been because most bryophytes and lichens are highly resilient to short-term fluctuations in their environment, and additional time may have been required to observe any effect. In addition, the pulse treatment may have required a higher amplitude (i.e., more “extreme” warming) to have a direct effect, as a die-off response to increased temperature has been reported for Sphagnum (Bragazza 2008). In our study, ventilated OTCs and OTCs increased the temperature with 1.00 and 1.87 °C, respectively, whereas CTCs and pulse treatments increased the temperature by 3.54 °C above average. Thus, the mean temperature increase from Pulse treatment with CTCs was actually higher than the heat wave that caused die-off of Sphagnum during a heat wave over 5 months, where they experience a mean average monthly temperature of 13.5 °C compared to a normal temperature of 11.5 °C (Bragazza 2008). However, the heat wave concurred with a drought spell, causing interactive effects of increased temperature and water stress (Bragazza 2008). The treatments in our experiment may have affected the moisture levels by keeping out precipitation and by increasing evaporation rates following the increased temperatures. Since extreme events of climate change are anticipated to coincide with lower precipitation and higher evaporation (e.g., De Boeck et al. 2010), our treatments may show a rather realistic scenario. Unfortunately, it is difficult to compare our treatments with the few studies available on extreme events due to different methodological problems. For example, Bragazza (2008) used climatic thresholds (i.e., the ratio of precipitation to temperature) as a measure of the detrimental effects of extreme events on Sphagnum. Even so, compared to a climatic threshold, our CTCs (closed open-top chambers) used in the pulse treatment with no precipitation and a mean average temperature increase of 3.54 °C for the summer period must be considered a rather extreme perturbation. Still, we did not notice any sign of die-off and in contrast the only significant responses were increases in the abundance and biomass of acrocarpous bryophytes. A possible explanation for this may be that we did not experimentally impose “draught” that was severe enough as the experimental plots had no water barrier to the surrounding soil.
A study by Klanderud (2008) reported almost the opposite effects: some bryophyte species decreased in abundance with warming perturbations, whereas there were no significant effects at the community level. In our study, there was a significant effect at the community level but on species level, only one species changed significantly in abundance (T. nitens). The differences found between community and species-level responses between our study and Klanderud’s (2008) may depend upon initial site-specific such as, for example, moisture conditions or species composition. As Klanderud’s experiment (2008) was conducted in a dry heath and ours in a meadow, the differences in moisture conditions might have affected the responses of the community by limiting the responses of bryophytes and/or lichens. Moisture availability will significantly affect bryophytes and lichens ability to respond to temperature treatments since they are greatly dependent on water for their photosynthesis and growth.
In our study, T. nitens, a pleurocarpous bryophyte, decreased in control plots and to the stepwise warming treatment but increased generally in 1997, an exceptionally warm year. Still, acrocarpous bryophytes as a group increased in abundance and biomass to the stepwise warming treatment. Even though acrocarpous and pleurocarpous species may have different ecology due to their different growth forms, there is no clear explanation for different responses between acrocarpous bryophytes as a group compared to the responses by T. nitens. However, acrocarpous species are known to possess more advanced water conducting structures compared to the more basic structures found in pleurocarpous species (e.g. Glime 2007) and that could be beneficial when exhibiting drier circumstances, for example as in the stepwise warming treatment.
T. nitens had a non-significant lower growth increase in response to a temperature warming experiment during the summer 1996 at the same site (Jägerbrand 2007). Yet, 4 years of standard OTC warming treatment between 1995 and 1999 did not result in any significant change in the abundance of T. nitens (Jägerbrand et al. 2009). Thus, it is uncertain what responses this species have to different temperature regimes but it is possible it may be able to increase in growth when other environmental factors are beneficial (such as in the year 1997), while studies of relatively small temperature increases, such as OTCs per se fail to induce significant responses.
Confounding responses of bryophytes have previously been shown in longer-term warming experiments comparing responses between two continents (Lang et al. 2012). Lang et al. (2012) found that non-Sphagnum bryophytes showed a negative response to warming at a low-altitude site in Sweden, and the Shannon index, species richness and abundance of lichens, liverworts and non-Sphagnum mosses all significantly decreased in response to warming at Toolik Lake, Alaska. In addition, they found that warming treatments had no significant effect on Sphagnum at the two sites (Lang et al. 2012).
Further, in a meta-analysis of 61 warming experiments across the tundra biome, Elmendorf et al. (2012) found that lichens decreased in almost all experiments included in the study, bryophytes on the other hand were found to have contrasting responses. However, the data were grouped as “mosses” and “lichens”. This can be compared with grouping on the level of “vascular plants”, and gives little predictive information about the responses on functional group level of bryophytes (acrocarpous and pleurocarpous bryophytes and Sphagnum spp.), or species level responses of bryophytes and lichens. Consequently, temperature responses by bryophytes as functional groups or at the species level may show rather divergent patterns, depending on study site, the study species and other abiotic and biotic factors. Additionally, bryophyte populations of Pleurozium schreberi originating from different altitudes have been shown to vary in responses to short-term temperature treatments ex situ depending on site of origin (Jägerbrand et al. 2014). It is unknown how common such site-dependent responses are in other bryophyte species but site-dependent response patterns may explain some of the confounding patterns show by bryophytes to experimental treatments performed at different sites.
Above-average temperatures were recorded in the summers of 1996 and 1997 at the high-alpine site. The mean summer monthly temperature in 1996 was the highest on record in the 20th century (at the time of the study) in the northern Scandes, and the mean August temperature in 1997 was again the highest on record since the onset of meteorological observations in the region (at the time of the study). Together, these two consecutive seasons of unusually warm conditions for plant growth and reproduction represent a rare event, which was unique in the 20th century (Molau 2001). Because treatment responses may be attenuated in warm years (see Molau 2001), this situation may have had a negative influence on the study, diminishing the differences in experimentally induced responses. Thus, the intended warming experiments were also accompanied by a “natural forcing” effect similar to what that was found in a nearby 5 years experiment with nutrient enhancement and warming (Jägerbrand et al. 2009). However, many bryophytes species have been reported to have a temperature optima between +15 and +25 °C, with bryophytes from polar regions having somewhat lower temperature optima, +10 °C to +15 °C (Oechel and Collins 1976; Furness and Grime 1982). Therefore, it is not likely that the unusually warm summers had mean temperatures above the temperature optima reported for bryophytes from polar regions, with only a few days surpassing the temperature optima.
As hypothesised, the pulse warming had a significant effect. However, this effect was limited to acrocarpous bryophytes, which showed a positive response in terms of the relative change in abundance and biomass in the first year. Pleurocarpous bryophytes in general, Sphagnum and lichens did not show any significant response to the pulse warming. We found no support for the hypothesis that stepwise warming would have a significant third-year effect compared to the other warming treatments. One species, T. nitens decreased significantly to stepwise warming and control treatments, but in general, bryophytes and lichens appeared to be more conservative in their responses to experimental warming than many vascular plants. It is also possible that the experimental warming was within the natural span of temperature variation at the site or that the magnitude of the pulse was insufficient to elicit a response. Thus, even more “extreme” warming or longer periods of extreme warming, might be needed to trigger significant effects. The power analysis indicated that the effect size was small and that the number of plots required would have needed to surpass 1200 for the experiment, pointing out the difficulty to conduct field experiments with high power in reality.
Conclusions
The results suggest that bryophytes and lichens, both at the functional group and species level, are to large extent resilient to the various climate change warming simulations that were applied, which included a constant elevated temperature, a single pulse warming event, and a stepwise increase in warming among years. Thus, the results support previous studies showing that bryophytes, to a large extent, are resistant to short-term warming perturbations in simulated climate change experiments (at a constant elevated temperature). While our pulse warming treatment surpassed the mean monthly temperature increase during the heat wave that caused the Sphagnum die-off reported by Bragazza (2008), we found no such drastic effects on bryophytes and lichens. Thus an extreme climatic warming event may need to be accompanied by draught to cause high mortality in bryophytes and lichens. Our results supported the hypothesised pulse effect on acrocarpous bryophytes, increasing their abundance and biomass, but not for other functional groups. Neither the constant OTC warming nor the stepwise warming had any impact on abundance of bryophytes or lichens at the functional group, or on species level (with exception for T. nitens).
References
Alatalo JM (1998) Climate change: impacts on structure and biodiversity of subarctic plant communities. PhD thesis, Göteborg University, Sweden. http://hdl.handle.net/2077/16349
Alatalo JM, Little CJ (2014) Simulated global change: contrasting short and medium term growth and reproductive responses of a common alpine/Arctic cushion plant to experimental warming and nutrient enhancement. SpringerPlus 3:157
Alatalo JM, Molau U (1995) Effect of altitude on the sex ratio in populations of Silene acaulis. Nordic J Bot 15:251–256
Alatalo JM, Totland Ø (1997) Response to simulated climate change in an alpine and sub-arctic pollen-risk strategist, Silene acaulis. Glob Change Biol 3:74–79
Arft AM, Walker MD, Gurevitch J et al (1999) Responses of tundra plants to experimental warming: meta-analysis of the International Tundra Experiment. Ecol Monogr 69:491–511
Biasi C, Meyer H, Rusalimova O et al (2008) Initial effects of experimental warming on carbon exchange rates, plant growth and microbial dynamics of a lichen-rich dwarf shrub tundra in Siberia. Plant Soil 307:191–205
Bjerke JW (2010) Winter climate change: ice encapsulation at mild subfreezing temperatures kills freeze-tolerant lichens. Environ Exp Bot 72:404–408
Bjerke JW, Bokhorst S, Zielke M et al (2011) Contrasting sensitivity to extreme winter warming events of dominant sub-Arctic heathland bryophyte and lichen species. J Ecol 99:1481–1488
Bokhorst S, Huiskes A, Convey P et al (2007) The effect of environmental change on vascular plant and cryptogam communities from the Falkland Islands and the Maritime Antarctic. BMC Ecol 7:1–13
Bokhorst S, Huiskes A, Convey P et al (2011) Microclimate impacts of passive warming methods in Antarctica: implications for climate change studies. Polar Biol 34:1421–1435
Bragazza L (2008) A climatic threshold triggers the die-off of peat mosses during an extreme heat wave. Glob Change Biol 14:2688–2695
Chapin FS III, Shaver GR, Giblin AE et al (1995) Responses of arctic tundra to experimental and observed changes in climate. Ecology 76:694–711
Chapin FS III, Bret-Harte MS, Hobbie SE et al (1996) Plant functional types as predictors of transient responses of arctic vegetation to global change. J Veg Sci 7:347–358
Cornelissen JHC, Callaghan TV, Alatalo JM et al (2004) Global change and arctic ecosystems: is lichen decline a function of increases in vascular plant biomass? J Ecol 89:984–994
De Boeck HJ, Dreesen FE, Janssens IA, Nijs I (2010) Climatic characteristics of heat waves and their simulation in plant experiments. Glob Change Biol 16:1992–2000
Dorji T, Totland Ø, Moe SR et al (2013) Plant functional traits mediate reproductive phenology and success in response to experimental warming and snow addition in Tibet. Glob Change Biol 19:459–472
Dormann CF, Woodin SJ (2002) Climate change in the Arctic: using plant functional types in a meta-analysis of field experiments. Funct Ecol 16:4–17
Dugas W, Reicosky D, Kiniry J (1997) Chamber and micrometeorological measurements of CO2 and H2O fluxes for three C4 grasses. Agric For Meteorol 83:113–133
Elmendorf SC, Henry GHR, Hollister RD et al (2012) Global assessment of experimental climate warming on tundra vegetation: heterogeneity over space and time. Ecol Lett 15:164–175
Faul F, Erdfelder E, Buchner A, Lang A-G (2009) Statistical power analyses using G*Power 3.1: tests for correlation and regression analyses. Behav Res Methods 41:1149–1160
Furness SB, Grime JP (1982) Growth rate and temperature responses in bryophytes: II. A comparative study of species of contrasted ecology. J Ecol 70:525–536. doi:10.2307/2259920
Glime JM (2007) Bryophyte ecology. Volume 1, physiological ecology. Chapter 7–1 water relations. Ebook sponsored by Michigan Technological University and the International Association of Bryologists. Accessed 27 May 2014. http://www.bryoecol.mtu.edu/
Graglia E, Jonasson S, Michelsen A et al (2001) Effects of environmental perturbations on abundance of subarctic plants after three, seven and ten years of treatments. Ecography 24:5–12
Hågvar S, Klanderud K (2009) Effect of simulated environmental change on alpine soil arthropods. Glob Change Biol 15:2972–2980. doi:10.1111/j.1365-2486.2009.01926.x
Hallingbäck T, Hedenäs L, Weibull H (2006) Ny checklista för Sveriges Mossor. Svensk Botanisk Tidskrift 100:96–148
Harden JW, Manies KL, Turetsky MR et al (2006) Effects of wildfire and permafrost on soil organic matter and soil climate in interior Alaska. Glob Change Biol 12:2391–2403
Hofmann H, Urmi E, Bisang I et al (2007) Retrospective assessment of frequency changes in Swiss bryophytes over the last two centuries. Lindbergia 32:18–32
Hollister RD, Webber PJ (2000) Biotic validation of small open-top chambers in a tundra ecosystem. Glob Change Biol 6:835–842
Hudson JMG, Henry GHR (2010) High Arctic plant community resists 15 years of experimental warming. J Ecol 98:1035–1041
IPCC (2007) Climate change 2007: impacts, adaptation and vulnerability: working group II contribution to the fourth assessment report of the IPCC intergovernmental panel on climate change. Cambridge University Press, Cambridge
Jägerbrand AK (2007) Effects of an in situ temperature increase on the short-term growth of arctic-alpine bryophytes. Lindbergia 32:82–87
Jägerbrand AK, Molau U, Alatalo JM (2003) Responses of bryophytes to simulated environmental change at Latnjajaure, northern Sweden. J Bryol 25:163–168
Jägerbrand AK, Lindblad KEM, Björk RG et al (2006) Bryophyte and lichen diversity under simulated environmental change compared with observed variation in unmanipulated alpine tundra. Biodivers Conserv 15:4453–4475
Jägerbrand AK, Alatalo JM, Chrimes D et al (2009) Plant community responses to 5 years of simulated climate change in meadow and heath ecosystems at a subarctic-alpine site. Oecologia 161:601–610
Jägerbrand AK, Kudu G, Alatalo JM et al (2012) Effects of neighbouring vascular plants on the abundance of bryophytes in different vegetation types. Polar Sci 6:200–208
Jägerbrand AK, Alatalo JM, Kudo G (2014) Variation in responses to temperature treatments ex situ of the moss Pleurozium schreberi (Willd. ex Brid.) Mitt. originating from eight altitude sites in Hokkaido, Japan. J Bryol. doi:10.1179/1743282014Y.0000000095
Jentsc A, Beierkuhnlein C (2008) Research frontiers in climate change: effects of extreme meteorological events on ecosystems. CR Geosci 340:621–628
Jentsch A, Kreyling J, Beierkuhnlein C (2007) A new generation of climate-change experiments: events, not trends. Front Ecol Environ 5:365–374
Karlsson U (1998) Evaluation of pointframe cover assessment for estimating biomass in alpine tundra. BSc thesis. Göteborg University, Sweden
Klanderud K (2008) Species-specific responses of an alpine plant community under simulated environmental change. J Veg Sci 193:363–372
Knapp AK, Yavitt JB (1992) Evaluation of a closed-chamber method for estimating methane emissions from aquatic plants. Tellus B 44:63–71
Lang SI, Cornelissen JHC, Shaver GR et al (2012) Arctic warming on two continents has consistent negative effects on lichen diversity and mixed effects on bryophyte diversity. Glob Change Biol 18:1096–1107. doi:10.1111/j.1365-2486.2011.02570.x
Longton RE (1984) The role of bryophytes in terrestrial ecosystems. J Hattori Bot Lab 55:147–163
Makkonen M, Berg MP, van Hal JR et al (2011) Traits explain the responses of a sub-arctic Collembola community to climate manipulation. Soil Biol Biochem 43:377–384. doi:10.1016/j.soilbio.2010.11.004
Marion GM, Henry GHR, Freckman DW et al (1997) Open-top designs for manipulating field temperatures in high-latitude ecosystems. Glob Change Biol 3:20–32
Moberg R, Holmåsen I (1990) Lavar, 3rd edn. Interpublishing, Stockholm
Molau U (2001) Tundra plant responses to experimental and natural temperature changes. Memoirs Natl Inst Polar Res Spec Issue 54:445–466
Molau U (2010) Long-term impacts of observed and induced climate change on tussock tundra near its southern limit in northern Sweden. Plant Ecol Divers 3:29–34
Molau U, Alatalo JM (1998) Responses of subarctic-alpine plant communities to simulated environmental change: biodiversity of bryophytes, lichens and vascular plants. Ambio 27:322–329
Nie D, He H, Mo G et al (1992) Canopy photosynthesis and evapotranspiration of rangeland plants under doubled carbon dioxide in closed-top chambers. Agric For Meteorol 61:205–217
Oechel WC, Collins NJ (1976) Comparative CO2 exchange patterns in mosses from two tundra habitats at Barrow, Alaska. Can J Bot 54:1355–1369. doi:10.1139/b76-148
Oksanen J, Guillaume Blanchet F, Kindt R et al (2013) Community ecology package, package ‘vegan’ version 20-7. URL http://cran.r-project.org, http://vegan.r-forge.r-project.org. Accessed 19 March 2013
Olsrud M, Melillo JM, Christensen TR et al (2004) Response of ericoid mycorrhizal colonization and functioning to global change factors. New Phytol 162:459–469
Pauli H, Gottfried M, Dullinger S et al (2012) Recent plant diversity changes on Europe’s mountain summits. Science 336:353–355
Polunin N (1951) The real Arctic: suggestions for its delimitation, subdivision and characterization. J Ecol 39:308–315
Potter JA, Press MC, Callaghan TV et al (1995) Growth responses of Polytrichum commune and Hylocomium splendens to simulated environmental change in the sub-arctic. New Phytol 131:533–541
Press M, Potter J, Burke M et al (1998) Responses of a subarctic dwarf shrub heath community to simulated environmental change. J Ecol 86:315–327. doi:10.1046/j.1365-2745.1998.00261.x
Reicosky DC (1990) Canopy gas exchange in the field: closed chambers. Remote Sens Rev 5:163–177
Rinnan R, Stark S, Tolvanen A (2009) Responses of vegetation and soil microbial communities to warming and simulated herbivory in a subarctic heath. J Ecol 97:788–800. doi:10.1111/j.1365-2745.2009.01506.x
Romanovsky VE, Drozdov DS, Oberman NG et al (2010) Thermal state of permafrost in Russia. Permafrost Periglac Process 21:136–155
Scherrer D, Körner C (2011) Topographically controlled thermal-habitat differentiation buffers alpine plant diversity against climate warming. J Biogeogr 38:406–416
Smith MD (2011) The ecological role of climate extremes: current understanding and future prospects. J Ecol 99:651–655
Soudzilovskaia NA, Graae BJ, Douma JC et al (2011) How do bryophytes govern generative recruitment of vascular plants? New Phytol 190:1019–1031
Steduto P, Çetinkökü Ö, Albrizio R, Kanber R (2002) Automated closed-system canopy-chamber for continuous field-crop monitoring of CO2 and H2O fluxes. Agric For Meteorol 111:171–186
Turetsky MR, Bond-Lamberty B, Euskirchen E et al (2012) The resilience and functional role of moss in boreal and arctic ecosystems. New Phytol 196:49–67
Underwood AJ (1994) On beyond BACI: sampling designs that might reliably detect environmental disturbances. Ecol Appl 4:3–15
Van Wijk MT, Clemmensen KE, Shaver GR et al (2003) Long-term ecosystem level experiments at Toolik Lake, Alaska, and at Abisko, Northern Sweden: generalizations and differences in ecosystem and plant type responses to global change. Glob Change Biol 10:105–123. doi:10.1046/j.1529-8817.2003.00719.x
Vitt DH, Pakarinen P (1977) The bryophyte vegetation production and organic components of Truelove Lowland. In: Bliss LC (ed) Truelove Lowland, Canada: a high Arctic ecosystem. University of Alberta Press, Edmonton, pp 225–244
Walker MD (1996) Community baseline measurements for ITEX studies. In: Molau U, Miolgaard P (eds) ITEX Man, 2nd edn. Danish Polar Centre, Copenhagen, pp 39–41
Walker MD, Wahren HC, Hollister RD et al (2006) Plant community responses to experimental warming across the Tundra Biome. PNAS 103:1342–1346
Wielgolaski FE, Bliss LC, Svoboda J et al (1981) Primary production of tundra. In: Bliss LC et al (eds) Tundra ecosystems: a comparative analysis. Cambridge University Press, Cambridge, pp 187–226
Acknowledgments
The authors thank two anonymous reviewers and Christian Rixen for their comments that improved the manuscript. We thank the staff of Abisko Scientific Research Station for help and hospitality, Vivian Aldén, Björn Aldén and Olga Khitun for assistance in the field. This study was supported by an NFR grant (B-AA/BU 08424) to UM.
Author information
Authors and Affiliations
Corresponding author
Additional information
This article is part of the special issue Vegetation in cold environments under climate change.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Supplemental Information
Supplemental information for this article (Figs. S1, S2 and Table S1) can be found online at:
Fig. S1
Boxplots of abundance on species level of bryophytes after different warming treatments and years at Latnjajaure. A) Anthelia juratzkana, B) Aulacomnium turgidum, C) Blepharostoma trichophyllum, D) Dicranum elongatum, E) Dicranum groenlandicum, F) Hylocomium splendens, G) Polytrichum strictum, H) Ptilidium ciliare, and I) Sphagnum warnstorfii. Treatments: control (Control), constant warming enhancement using open-top chambers (OTC), a stepwise increase in the magnitude of warming (Step) and a single-summer high-impact warming event (Pulse). Boxplots show the 10th to 90th percentile of the data; n = 4 plots per treatment. Supplementary material 1 (DOCX 2,626 kb)
Fig. S2
Boxplots of the abundance at the species level for lichens after different warming treatments and years at Latnjajaure Field Station. A) Cladina arbuscula, B) Cladonia uncialis, C) Ochrolechia frigida, and D) Sphaerophorus globosus. Treatments: control (Control), constant warming using open-top chambers (OTC), a stepwise increase in the magnitude of warming (Step) and a single-summer high-impact warming event (Pulse). Boxplots show the 10th to 90th percentile of the data; n = 4 plots per treatment. Supplementary material 2 (DOCX 1,173 kb)
Rights and permissions
About this article

Cite this article
Alatalo, J.M., Jägerbrand, A.K. & Molau, U. Climate change and climatic events: community-, functional- and species-level responses of bryophytes and lichens to constant, stepwise, and pulse experimental warming in an alpine tundra. Alp Botany 124, 81–91 (2014). https://doi.org/10.1007/s00035-014-0133-z
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00035-014-0133-z