
Abstract
Lago de Tota, the largest lake in Colombia, is the primary source of water for 250,000 people and a focus of regional economic activity in agriculture, aquaculture, and tourism. Recently, agencies and stakeholders report a shift from the naturally oligotrophic state toward eutrophy. However, the relative contributions of different inputs, including agricultural runoff, aquaculture and municipal wastewaters are unknown, hampering efforts to mitigate nutrient loading. We examined spatial and temporal variation in the trophic state of the lake over one year, as well as stable C and N isotopic profiles of aquatic producers and consumers in relation to two main potential sources (fertilizer and trout feed). We found that Lago de Tota is moderately eutrophic (average chlorophyll-a: 6.4 µg/L, TN: 1.5 mg/L and TP: 0.06 mg/L) with a 32% reduction of transparency over the last 15 years. δ15N and δ13C of aquatic organisms and surface sediments were enriched relative to prehistoric sediments, indicating that human sources dominate the C and N cycles of the lake. δ15N of macrophytes (15.7‰), particulate organic matter (12.5‰), and invertebrates (20.2‰) were enriched relative to trout food (4.6‰), but similar to chicken manure (13.7‰), suggesting that farming in the watershed may be a more important source of N than aquaculture. Our results indicate that Lago de Tota is on a trajectory toward eutrophication with potentially severe consequences for water resources in a rapidly developing mountain region.
Similar content being viewed by others


Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Eutrophication is a consequence of both lake aging (Lindeman 1942) and human activities that increase nutrient loading. Cultural eutrophication remains a pervasive global threat to water security, economic development and the delivery of ecosystem services from freshwater environments (Carpenter et al. 1998; Smith and Schindler 2009). High elevation lakes in many regions may be less vulnerable to the effects of anthropogenic eutrophication because cold temperatures and steep slopes make their watersheds unsuitable for many agricultural and industrial activities that generate nutrient runoff. Mountain lakes may receive a larger proportion of nutrients from atmospheric transport in dissolved and particulate forms (Sickman et al. 2003). However, in the tropics, human settlement and farming can extend to high elevations, leading to potential eutrophication in mountain lakes at elevations that are less impacted by watershed development at temperate latitudes (Catalan and Donato Rondón 2016). Transitions to eutrophy may be difficult to reverse due to hysteresis driven by elevated internal phosphorus loading from anoxic sediments (Carpenter et al. 2015), and the tendency for polymixis to make regenerated P readily available (Lewis 1987). Anticipating and preventing such transitions may therefore be more effective than restoring water quality once lakes enter a degraded state (JØrgensen et al. 2013).
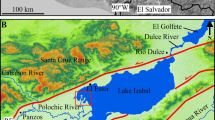
Lago de Tota (Boyacá, Colombia, Fig. 1) is the largest freshwater lake in Colombia with a surface area of 5620 hectares and 1920 million m3 of volume, located at an elevation of 3015 m in the eastern slope of the northern end of the Andes mountain range. Lago de Tota is of tectonic-glacial origin (Rangel and Aguirre 1983), and its watershed, which reaches to 3700 m of elevation, is heavily influenced by past glaciation. Tota Lake is the primary source of water for the city of Sogamoso (population: 113,000) as well as a number of smaller towns and villages Agriculture in the watershed consists primarily of green onion farming that has expanded in recent decades. A number of large aquaculture pens for rainbow trout cultivation are also present within the lake, which is an attraction for regional tourism. The lake has experienced a continuous descending trend in its seasonal water levels associated with sustained extractions for irrigation, industry, and urban supply and climate forcing (Cañon and Valdes 2011). A 1983 report determined that the oligotrophic state of Lago de Tota was at risk due to agricultural and aquacultural activities in the watershed and the lake (CAR 1983). Despite its size and importance, no consistent sampling program is in place for monitoring water quality in the lake.

A map showing the location of Lago de Tota and our sampling sites, and a photo of the lake (photo credit: Marika Schulhof)
Anecdotal evidence suggests a trend toward eutrophication in Lago de Tota. Beds of macrophytes, particularly Egeria densa, have reportedly expanded, creating barriers to navigation in the littoral zone. Water transparency has also reportedly declined. Figure 2 shows a comparison of average Secchi disc transparency and the density of algal cells from samples recorded in two sampling campaigns conducted between 2002 and 2005 and 2012–2015. The results of the second survey are reported in Muñoz-López et al. (2017), while the earlier survey is unpublished data of N.A. Between the two time periods, algal density increased more than five-fold and water transparency declined from 10 to 7 m (Fig. 2). The limited available information suggests a path toward increasingly eutrophic state in Lago de Tota.

a Comparison of algal density (organisms/ml, where organisms consist of single cells as well as colonies) and b Secchi disk transparency in samples collected in two time periods (2002–2005 and 2012–2015) in Lago de Tota. Abundance was estimated by chamber sedimentation method and observation under an inverted microscope (Utermöhl 1958) as described in Muñoz-López (2017). The difference between the two time periods is significant by two-tailed t test (both variables P < 0.0001)
Three potential anthropogenic sources of nutrients may contribute to the eutrophication of Lago de Tota. First, untreated waste water originates from the town of Aquitania (population 15,000) through the outlet of the La Mugre stream on the eastern shore of the lake. Second, the dominant land use in the watershed consists of extensive onion farms fertilized by gallinaza, a fertilizer consisting mainly of chicken manure. Third, twelve aquaculture pens for cultivating rainbow trout (Oncorhynchus mykiss, Walbaum 1792) are present in the lake, concentrated mainly at the northern end of the larger basin (Lago Grande). Lack of information about the contributions of these three sources to declining water quality in the lake hampers the implementation of strategies to remediate the loading of nutrients and reduce the risk of a transition to a eutrophic state with degraded water quality.
We examined temporal and spatial variation in the trophic state and limnological conditions of Lago de Tota over one year. We also compared stable isotopic ratios for C and N of particulate organic matter (POM, consisting mainly of phytoplankton), littoral amphipods (Hyalella sp.), macrophytes and superficial sediments with two putative sources of eutrophication: the chicken manure used to fertilize onion farm fields throughout the water shed, and the feed provided to rainbow trout in aquaculture pens situated in the surface waters of the lake. Our goals were to determine (1) the current trophic state of Lago de Tota and how it varies spatially among sites and the two main basins of the lake (Lago Chico and Lago Grande) and throughout the year, and (2) how the isotopic signatures of contemporary organisms compare to two of the main anthropogenic sources of nutrients that have been implicated as potential drivers of eutrophication. Despite its tremendous importance to the regional and national economy, little research has been conducted on the Lago de Tota ecosystem. Our study represents a first attempt at understanding the causes of ongoing changes in the lake and its watershed.
Materials and methods
We sampled ten stations in Lago de Tota eight times between 9th of September, 2014 and 3rd of September, 2015. Samples were collected approximately monthly until 25th of November, 2014, and again after 13th of July, 2015. The first sampling period occurred during the period of relatively high water levels, and the second during lower levels. Sampling stations were distributed throughout the two main basins of the lake and in order to cover gradients of proximity to the main population center (Aquitania, a town of around 15,000 on the eastern shore of “Lago Chico”, the smaller basin of the lake) and the zone with the greatest concentration of trout aquaculture activity (the northern end of “Lago Grande”, the larger basin). At each station, we recorded water column depth and Secchi depth transparency. Samples of a common benthic amphipod (Hyalella sp.) were collected from several locations with a sweep net from the littoral zone on the last sampling date, and surface sediments were collected with a grab sampler. Samples of the fertilizer used on onion crops and the feed used in trout farms were acquired from farmers.
We collected water samples for nutrient, isotope and chlorophyll analysis from the depth of Secchi transparency (between 5 and 7 m) using a Van Dorn sampler. Nutrient samples were fixed in the laboratory with concentrated sulfuric acid (approximately 12.5 µL for sample to reduce pH to 2) and refrigerated at 4 °C until analysis (APHA 1976). These were analyzed through flow injection analysis following persulfate digestion using a QuikChem® autoanalyzer equipped with TN and TP modules (Lachat instruments USA, 2013). Chlorophyll-a samples were filtered in the field using Advantec Glass Fiber that were kept in the dark at − 70 °C (APHA 1976). Chlorophyll was extracted from the filter in the lab with ketone and the extracts were analyzed through ultraviolet–visible spectroscopy using Jeffrey and Humphrey’s (1975) trichromatic equation.
A sample of Particulate Organic Matter (POM) was filtered into a pre-combusted Advantec (Vernon Hills, IL, USA) 0.47 µm Glass Fiber filters for stable C and N analysis. Samples of macrophytes, particulate organic matter (POM), littoral amphipods (Hyallela sp.), sediments, trout food and agriculture fertilizer were dried in an oven at 50–60 °C for 12 h, then ground, weighed to target weight for each material, and encapsulated in tin (Sn) capsules for Carbon (δ13C) and Nitrogen (δ15N) analysis at the U.C. Davis Stable Isotope Facility. Isotope samples were analyzed using an Elementar Vario EL Cube or Micro Cube elemental analyzer and PDZ Europa ANCA-GSL elemental analyzer interfaced to a PDZ Europa 20–20 isotope ratio mass spectrometer.
Stable isotope mixing models
In order to estimate the proportional contributions of chicken manure and trout feed to POM, macrophytes and invertebrates, we used a Bayesian mixing model based on δ13C and δ15N implemented in the R package “simmr” (Parnell 2016). Mixing models estimate consumer diets and the incorporation of different sources of nutrients into biological materials based on measured isotopic ratios. Bayesian model frameworks incorporate variability in both source and consumer isotopic ratios, and enrichment fractions, in estimating proportional contributions to properly account for all sources of error and uncertainty. The models require assumptions about means and variation in C and N isotopic fractionation between consumers and their sources which are based on values reported in the literature (Phillips et al. 2014). The two sources in our model were chicken manure and trout feed. For POM and macrophytes, we assumed no fractionation upon uptake (Moore et al. 2014), and the observed average and standard deviation of the measured δ13C and δ15N values of the two sources. For invertebrates, we assumed an average trophic enrichment for 15N of 3.4‰ (standard deviation = 1.0) and 0.4‰ for 13C (standard deviation = 1.3), following Post (2002). Estimates of proportional contributions of the two sources were based on 1000 Monte Carlo Markov Chain simulations.
The trophic state of Lago de Tota was determined by the Trophic State Index (TSI) of Carlson (1977), modified for the tropics by Toledo et al. (1983) based on Chlorophyll-a, Secchi disc transparency and total phosphorus concentrations. The criteria for trophic state assignments were: TSI > 44 = mesotrophic condition; TSI > 54 = eutrophic condition.
Results
During the period of our study, the daily average TN concentration of Lago de Tota was 1.5 mg/L (range 0.8 to 2.0 mg/L), TP averaged 0.06 mg/L (0.02–0.09 mg/L) and the average chlorophyll-a was 6.4 µg/L (3.4–9.9 µg/L). The N/P ratio by weight ranged among dates 18.1 to 35.7 among sampling dates (averaged across sites on each date), with an average value of 25.3.
The concentrations of chlorophyll and nutrients varied in space and time (Fig. 3). Chlorophyll-a concentration was greater in the first sampling period in 2014 than in 2015, and often higher in Lago Chico (the smaller eastern basin) than Lago Grande. The highest chlorophyll concentrations were often observed near the outflow of the La Mugre stream near the town of Aquitania on the eastern shore. The lowest nutrient concentrations were observed in 2014. Higher TN concentrations were often found in Lago Chico, while TP was often greater in Lago Grande, the larger western basin of the lake (Fig. 3). The trophic index of the lake varied between mesotrophic and eutrophic states during the duration of the study (Fig. 4). The Secchi disc transparency indicated that the lake was persistently eutrophic at all sites, while chlorophyll-a and TP most often fell within the mesotrophic range (Fig. 4).

Maps of Total Nitrogen (TN), Total Phosphorous (TP) and chlorophyll-a concentration in the surface waters of Lago de Tota at ten stations on eight sampling dates. The size of each circle is proportional to the concentration, and raw values are provided in the Supplementary Information. The point on the bottom right of each row indicates the size of the point corresponding to the concentration shown in mg/L (for TN and TP) and µg/L (for chlorophyll-a)

Trophic status indices based on chlorophyll-a, total phosphorus and Secchi disc transparency for the ten sample sites in Lago de Tota. The horizontal lines at 44 and 54 indicate the boundaries between oligotrophic, mesotrophic and eutrophic states
Both C and N isotopes were most enriched in littoral amphipods and macrophytes, followed by POM and the lowest in superficial sediments (Fig. 5). Chicken manure had isotopic ratios very similar to those of POM and macrophytes, with similar δ13C to trout feed but more enriched δ15N values. Trout feed and farm soils showed similar δ13C to both POM and macrophytes, but were more depleted δ15N by around 7–10‰. Amphipods had more enriched δ13C and δ15N values than either of the sources, or any of the other materials analyzed. Sediments from a paleo-limnological study by Cardozo et al. (2014) (dated between 4000 and 2000 YBP, and labeled “Ancient sediments” in Fig. 5) were much more depleted in heavy isotopes of both C and N, with δ15N values centered around 2 and δ13C around − 30. The C:N atomic ratio of the surface sediments (10.1, SD = 2.2) were similar to those of the ancient sediments (11.7, SD = 0.9) sampled by Cardozo et al. (2014). The C:N ratios of the other materials were 8.9, SD = 2.0 (macrophytes), 9.1, SD = 2.0 (POM), 3.9, SD = 0.5 (amphipods), and 11.2, SD = 1.8 (soil).

Carbon and Nitrogen stable isotopes of organisms (invertebrates, macrophytes and POM consisting mainly of phytoplankton), superficial and ancient sediments, along with two potential anhropogenic sources (chicken manure and trout feed). Each type of material is averaged over multiple samples collected through time over the course of our survey. The arrows show the difference in position between POM and the two potential sources, and the ancient sediments as an indicator of the pre-settlement isotopic ratios of the lake. Ancient settlement ratios were taken from sediment cores collected by Cardozo et al. (2014) dated between 2000 and 4000 YBP
The stable C and N isotopic signatures of different organismal groups also varied through time. Figure 6 shows the δ13C and δ15N profiles for each sampling date, with POM, chicken manure and trout feed highlighted in color. The δ15N of POM was more similar to chicken manure than trout feed during most of the sample period, but became more depleted on certain dates (e.g., September 24, 2014).

Stable carbon and nitrogen isotopes signatures through time. The orange “+” symbol indicates the mean of POM samples, and the chicken manure and trout feed are shown in blue and green, respectively. The symbols indicate the other points indicate the type of material as follows: POM (+), littoral amphipods (squares), macrophytes (triangles), superficial aquatic sediments (×) and soil (diamonds)
The Bayesian stable isotope mixing model estimated that chicken manure contributed 87.2% of POM mass (SD = 3.5%), 98.3% (SD = 1.1%) of macrophyte biomass, and 97% (SD = 2.1%) of invertebrate biomass (Fig. 7).

Results of mixing models showing the posterior distributions of estimated percent contributions of chicken manure and trout feed to POM (a) and the tissues of macrophytes (b) and littoral invertebrates (c) based on Monte Carlo Markov Chain simulations. The analyses were performed using the simmr package in R (Parnell 2016)
Discussion
Our study shows several indications that nutrient inputs have contributed to recent eutrophication of Lago de Tota, and that the lake may therefore be at risk for transition to a persistent eutrophic lake with a large increase in algal abundance and macrophyte bed expansion in recent decades. First, deep mountain lakes of the tropics such as Tota are typically oligotrophic (Löffler 1962; Lewis 1987; Catalan and Donato Rondón 2016). However our research indicated that chlorophyll and nutrient levels of Lago de Tota were in the mesotrophic to eutrophic range, which is evident in the increase of the phytoplankton biomass expressed as biovolume (Muñoz-López et al. 2017). Second, carbon and nitrogen stable isotopic signatures of phytoplankton, macrophytes and invertebrates were all enriched relative to values for aquatic ecosystems with low levels of anthropogenic nutrient impacts, as well as pre-settlement sediments from Lago de Tota. In addition, δ15N of aquatic organisms were also elevated compared to the feed used in trout aquaculture in the lake, but similar to chicken manure applied to green onion crops in the watershed. These results indicate that onion agriculture in the watershed may contribute the bulk of nutrients supporting elevated productivity in the Lake.
Nitrogen isotopic ratios have a long history as indicators of eutrophication and runoff in aquatic ecosystems (Cabana and Rasmussen 1996; Fry 2006), and the δ15N of organisms in Lago de Tota provide strong indication of major human influence. For instance, Cabana and Rasmussen (1996) showed that herbivorous zooplankton consumers have δ15N ratios around 2–5 in lakes with the lowest human population density in their watersheds, while in the most impacted lakes have invertebrate ratios are around 10–13. Littoral amphipods from Lago de Tota have δ15N around 20, indicating high anthropogenic contributions. A survey of species-specific δ15N signatures of pelagic zooplankton from Lago de Tota showed a range from 16.0 for Ceriodaphnia to 20.2 for Boeckella (Nidia Gil, unpublished data). Thus, the δ15N of the Tota Lake pelagic and benthic invertebrates is on par with some of the most eutrophic lakes globally. The amphipods in our data set also show highly enriched δ13C signatures, indicating that they may rely on unknown aquatic or terrestrial carbon sources. In addition, our δ15N ratios of POM (12.5) and macrophytes (15.7) are similar to or slightly higher than values observed in aquatic systems with the greatest contributions wastewater to the dissolved inorganic nitrogen (DIN) pool (Fig. 5 in Cole et al. 2005).
Carbon is less often used as an indicator of eutrophication than nitrogen; however, a variety of evidence suggests that δ13C signatures are also reliable indicators of eutrophication in aquatic ecosystems. Oczkowski et al. (2014) show that δ13C increased with coastal phytoplankton productivity in experiments and observations (see also Voss and Struck 1997). The enriched C isotopic ratios of producers and consumers observed in Lago de Tota also indicate elevated productivity due to human influence, and are heavier than those found in ancient sediments from before extensive agricultural development in the watershed (Cardozo et al. 2014).
Carbon and nitrogen isotopic signatures also provide some insight into the contributions of two putative sources of nitrogen supporting eutrophication in Lago de Tota: onion crops and trout aquaculture. We found that fertilizers used in onion crops (chicken manure) and the food provided to trout in aquaculture enclosures had similar δ13C signatures between − 22 and − 24; however, chicken fertilizer was more enriched in heavy nitrogen, with a mean δ15N of 13.7, compared with trout feed with a ratio of 4.6. Bayesian mixing models estimated a dominant role for terrestrial agriculture relative to in situ trout aquaculture in supporting growth of algae, aquatic plants and invertebrates. Although these ratios and the mixing models suggest a greater contribution of terrestrial agriculture than aquaculture as a source of N to organisms in the lake, a number of outstanding questions remain before the importance of different sources can be confidently inferred.
First, the roles of urban waste waters in fertilizing Lago de Tota remain unclear. The largest town in the watershed is Aquitania on the eastern shore, with a population of 15,000. Agriculture is the dominant land use in the watershed, with extensive green onion crops extending to the shores of the lake. Wastewater typically contains nitrogen with δ15N of 6 or greater (Moore et al. 2014), which is similar to the signature of trout feed and lower than that of chicken manure. The spatial pattern of POM and nutrients suggest that the town of Aquitania may be a contributor of nutrients, with elevated levels occurring periodically in the eastern basin of the lake near the outlet of the La Mugre stream. For instance, nitrogen concentrations were substantially elevated at the site closest to Aquitania on September 9 and October 31, 2014, and August 13, 2015 (Fig. 3) compared to other sampling stations in the lake. Urban wastewaters may therefore play some role in eutrophication of the lake, but cannot be included in the mixing models because their isotopic signatures are unknown.
Second, stimulation of microbial processes such as denitrification within the lake may elevate δ15N relative to primary sources. Dentrification in low oxygen conditions produces substantial enrichment in δ15N (Altabet et al. 1995), therefore the signatures of organisms in Lago de Tota may reflect processing of nitrogen within the lake that leads to enrichment with 15N rather than external sources. We found that the surface waters of lake were generally saturated with oxygen; however, denitrification in the sediments may affect the N isotopic signatures of POM or macrophytes. Fractionation on uptake may also enrich the nitrogen isotopes of phytoplankton or macrophytes (Fry 2006). In addition, Lago de Tota may be more sensitive to internal phosphorus loading due to its long renewal time of 30 years (Cañón and Rodríguez 2002). Phosphorus recycling from anoxic sediments may maintain high productivity even if external inputs are limited (Carpenter et al. 1999). The observed daily average TN:TP ratio by weight among all of our samples ranged from 18.1 to 35.7 with a mean of 25.3. Phosphorus limitation of phytoplankton growth typically occurs at TN:TP ratios above ~ 22 by mass (Guildford and Hecky 2000), indicating that primary productivity of Lago de Tota is most likely phosphorus limited. Thus, although the nitrogen in producers may be of primarily terrestrial origin, the major sources of P to the lake remains remain to be determined.
Finally, the contributions of agriculture and aquaculture to the nutrient budget of Lago de Tota may vary spatially and temporally. For instance, while the δ15N of POM was on average closer to that of chicken manure, during some sampling periods the ratio became more depleted and more similar to that of trout feed. For instance, on September 24, 2014 the δ15N of POM was closer to that of trout feed, suggesting that aquaculture may play a significant role during certain periods of the year. Sustained spatial and temporal sampling is needed to construct a detailed budget of nutrients throughout the watershed in order to identify and understand the causes of changes occurring in Lago de Tota.
The composition of the phytoplankton community also indicates that Lago de Tota is entering a eutrophic state. A recent survey by Muñoz-López et al. (2017) of Lago de Tota over a year found that limnological conditions were related to nutrients dynamics, particularly Total Nitrogen, which is influenced by seasonal level water variation. Second, they surveyed the functional traits of phytoplankton community and showed that the dominant morphological functional groups are typical of enriched systems, For example flagellated algae and diatoms contributed more biovolume in the smaller basin “Lago Chico” where trophic state indicators like TP were generally greater. These groups are often associated with high TP concentration in eutrophic lakes of Colombia (Duque and Donato-Rondón 1992) and other regions (Kruk et al. 2010). In addition, Muñoz-López et al. (2017) found that photic zone depth was the strongest environmental predictor of the functional traits of the Lago de Tota phytoplankton community over an annual cycle, indicating that changes in lake trophic status has a large influence on the traits of pelagic primary producers.
High-altitude lakes like Lago de Tota are typically oligotrophic with cold temperatures and low nutrients concentrations (Catalan and Donato Rondón 2016). Allochthonous organic matter and atmospheric deposition represent the principal sources of carbon and nitrogen (Auguet et al. 2011; Catalan and Donato Rondón 2016). Eutrophication of alpine lakes has mainly been studied in the context of atmospheric deposition of dissolved and particulate nutrients in temperate regions (Sickman et al. 2003); however, in the tropics, high elevation systems may be at risk from watershed sources as well. For instance, Lake Titicaca in Bolivia has undergone eutrophication as a result of expansion of mining activities and accompanying urban population growth (Archundia et al. 2017). Our results indicate that Lago de Tota is presently in a mesotrophic to eutrophic state, and may be at risk of transition to a similar degraded state.
Despite its regional and national importance as a source of water resources and focus of economic activities in agriculture, aquaculture and tourism, almost no information is available about the status of Lago de Tota or its trajectory through time. Our study is the first to show human disruption of the nutrient cycles of the lake and its watershed, and our data raise concern about the risk of potential catastrophic loss of water resources in this mountain region. Studies of potential nutrient retention mechanisms and wastewater treatment to control runoff from farms and towns in the watershed are badly needed to insure that Lago de Tota does not undergo transition to a degraded eutrophic state. Such a loss of water resources could be severely economically and socially costly in a developing region that is vulnerable to water and food insecurity.
References
Altabet MA, Francois R, Murray DW, Prell WL (1995) Climate-related variations in denitrification in the arabian sea from sediment n-15/n-14 ratios. Nature 373:506–509
APHA (1976) Standard methods for the examination of water and wastewater. In. D.C, American Public Health Association Washington
Archundia D, Duwig C, Spadini L, Uzu G, Guedron S, Morel MC, Cortez R, Ramos OR, Chincheros J, Martins JMF (2017) How Uncontrolled Urban Expansion Increases the Contamination of the Titicaca Lake Basin (El Alto, La Paz, Bolivia). Water Air and Soil Pollution, 228
Cabana G, Rasmussen JB (1996) Comparison of aquatic food chains using nitrogen isotopes. Proc Natl Acad Sci USA 93:10844–10847
Cañon J, Valdes J (2011) Assessing the influence of global climate and anthropogenic activities on the water balance of an Andean Lake. J Water Resour Protect 3:883–891
Cañón J, Rodríguez C (2002) Análisis isotópico en el lago Tota. XV Seminario Nacional de Hidráulica e Hidrología. In: Medellin, Colombia
CAR (1983) Informe sobre el Lago de Tota Consultado de Richard Vollenweider. In: Corporación Autónoma Regional CAR y Organización Panamericana de la Salud. Bogotá, Colombia Bogota, Colombia, p 26 pp
Cardozo AYV, Gomes DF, da Silva EM, Duque SRE, Rangel JOC, Sifeddine A, Turcq B, Albuquerque ALS (2014) Holocene paleolimnological reconstruction of a high altitude Colombian tropical lake. Palaeogeogr Palaeoclimatol Palaeoecol 415:127–136
Carlson RE (1977) Trophic state index for lakes. Limnol Oceanogr 22:361–369
Carpenter SR, Caraco NF, Correll DL, Howarth RW, Sharpley AN, Smith VH (1998) Nonpoint pollution of surface waters with phosphorus and nitrogen. Ecol Appl 8:559–568
Carpenter SR, Ludwig D, Brock WA (1999) Management of eutrophication for lakes subject to potentially irreversible change. Ecol Appl 9:751–771
Carpenter SR, Brock WA, Folke C, van Nes EH, Scheffer M (2015) Allowing variance may enlarge the safe operating space for exploited ecosystems. Proc Natl Acad Sci USA 112:14384–14389
Catalan J, Donato Rondón JC (2016) Perspectives for an integrated understanding of tropical and temperate high-mountain lakes. J Limnol 75:215–234
Cole ML, Kroeger KD, McClelland JW, Valiela I (2005) Macrophytes as indicators of land-derived wastewater: application of a delta N-15 method in aquatic systems. Water Resources Research 41
Duque SR, Donato-Rondón J (1992) Biología y Ecología del fitoplancton de las aguas dulces en Colombia. Cuadernos Divulgativos 35:1–21
Fry B (2006) Stable isotope ecology. Springer, New York
Guildford SJ, Hecky RE (2000) Total nitrogen, total phosphorus, and nutrient limitation in lakes and oceans: is there a common relationship? Limnol Oceanogr 45:1213–1223
Jeffrey SW, Humphrey GF (1975) New spectrophotometric equations for determining chlorophylls A, B, C1 and C2 in higher-plants, algae and natural phytoplankton. Biochemie Physiologie Der Pflanzen 167:191–194
Jørgensen S, Tundisi J, Tundisi M (2013) Handbook of inland aquatic ecosystem management. CRC Press, Boca Raton
Kruk C, Huszar VLM, Peeters E, Bonilla S, Costa L, Lurling M, Reynolds CS, Scheffer M (2010) A morphological classification capturing functional variation in phytoplankton. Freshw Biol 55:614–627
Lewis WM (1987) Tropical limnology. Annu Rev Ecol Syst 18:159–184
Lindeman R (1942) The trophic-dynamic aspect of ecology. Ecology 23:399–418
Löffler H (1962) The limnology of tropical high-mountain lakes. Internationale Vereinigung fuer Theoretische Angewandte Limnologie Verhandlungen 15:176–193
Moore JW, Lambert TD, Heady WN, Honig SE, Osterback AMK, Phillis CC, Quiros AL, Retford NA, Herbst DB (2014) Anthropogenic land-use signals propagate through stream food webs in a California, USA, watershed. Limnologica 46:124–130
Muñoz-López CL, Aranguren-Riano NJ, Duque SR (2017) Functional morphology of phytoplankton in a tropical high mountain lake: Tota Lake (Boyaca-Colombia). Revista De Biologia Tropical 65:669–683
Oczkowski A, Markham E, Hanson A, Wigand C (2014) Carbon stable isotopes as indicators of coastal eutrophication. Ecol Appl 24:457–466
Parnell A (2016) Package ‘simmr’: a stable isotope mixing model. In
Phillips DL, Inger R, Bearhop S, Jackson AL, Moore JW, Parnell AC, Semmens BX, Ward EJ (2014) Best practices for use of stable isotope mixing models in food-web studies. Can J Zool 92:823–835
Post DM (2002) Using stable isotopes to estimate trophic position: Models, methods, and assumptions. Ecology 83:703–718
Rangel O, Aguirre J (1983) Comunidades acuáticas altoandinas—I. Vegetación sumergida y de ribera en el lago de Tota. Boyacá Colombia Caldasia 65:719–742
Sickman JO, Melack JM, Clow DW (2003) Evidence for nutrient enrichment of high-elevation lakes in the Sierra Nevada, California. Limnol Oceanogr 48:1885–1892
Smith VH, Schindler DW (2009) Eutrophication science: where do we go from here? Trends Ecol Evol 24:201–207
Toledo APd, Talarico M, García E (1983) A aplicacao de modelos simplificados para avaliacao do processo de eutrofizacao em lagos e reservatorios tropicais. In. CETESB Sao Paulo
Utermöhl H (1958) Zur Vervollkommnung der quantitative Phytoplankton-Methodik. Mitteilungen des Internationalen Limnologie 9:1–38
Voss M, Struck U (1997) Stable nitrogen and carbon isotopes as indicator of eutrophication of the Oder river (Baltic sea). Mar Chem 59:35–49
Acknowledgements
The Corpoboyacá and Fulbright Colombia provided funding for our project. We thank Alejandra Jiménez and Nidia Gil for helpful assistance in field.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
Aranguren-Riaño, N.J., Shurin, J.B., Pedroza-Ramos, A. et al. Sources of nutrients behind recent eutrophication of Lago de Tota, a high mountain Andean lake. Aquat Sci 80, 39 (2018). https://doi.org/10.1007/s00027-018-0588-x
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s00027-018-0588-x