
Abstract
We examined the influence of land-use, habitat, and water quality on the spatial distribution of aquatic macroinvertebrates in two human-dominated catchments in the Swiss Plateau (Gürbe, Mönchaltorfer Aa). Land-use in the Gürbe catchment was dominated by agriculture, whereas urban land-use was more common in the Mönchaltorfer Aa. Study sites in each catchment were characterized using measures of local habitat conditions, water quality parameters including water temperature, and organic matter resources. A strong longitudinal gradient in temperature, conductivity and nitrogen was evident among sites in the Gürbe catchment, although sites on a main tributary had a strong agricultural signature and deviated from this pattern. Percentage agricultural land-use in the Gürbe was strongly correlated with algal biomass and the water quality PCA axes associated with conductivity, nitrogen (axis-1) and temperature (axis-3). Spatial grouping of sites by water quality was less evident in the Mönchaltorfer Aa, except for a strong signal by wastewater treatment plant effluents and partial differences between upper and lower basin sites. Percentage forest and agricultural land-use in the Mönchaltorfer Aa were correlated with water quality PCA axis-2, being associated with phosphorus and temperature. Macroinvertebrate densities, taxonomic richness, and axis-1 from a non-metric multidimensional scaling analysis (NMDS) of taxonomic composition were significantly correlated with water quality PCA axis-1 in the Gürbe catchment. Here, macroinvertebrate densities and NMDS axis-1 scores based on taxon relative abundances and densities were correlated with land-use features. Spatial distances between sites also were related to site differences in macroinvertebrates, reflecting the strong longitudinal environmental gradient in the Gürbe. Taxonomic differences between water quality PCA site groups were less pronounced in the Mönchaltorfer Aa, although differences were significant for trichopterans, ephemeropterans, chironomids, gastropods and coleopterans. Here, NMDS axis-1 based on taxon relative abundances and densities was correlated with forest land-use. Spatial distances between sites were not evident in macroinvertebrate site differences, reflecting the less pronounced spatial and longitudinal patterns in environmental attributes in this catchment. Our results support the hypothesis that spatial distributions of macroinvertebrates are related to spatial relationships among environmental attributes like land-use, habitat, and water quality in human-dominated catchments that depend on river network complexity, a habitat-filtering template in line with ecological niche theory.
Similar content being viewed by others
Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Most rivers are longitudinally-connected networks in the landscape that display a hierarchical spatial-structuring at multiple scales (Vannote et al. 1980; Allan et al. 1997). For example, coarse-scale river properties are dictated by regional patterns in geology and climate that influence flow and temperature regimes along with background characteristics in water quality (Frissell et al. 1986; Amoros and Bornette 2002). More fine-scaled patterns occur at local scales related to spatial linkages with the surrounding landscape and inputs from upstream sources (Richards et al. 1997). Even finer-scaled properties are reflected in meso-habitats associated with differences in substrate and flow characteristics within a particular stream reach (e.g. Sponseller et al. 2001). This spatial hierarchy forms the so-called habitat template for flora and fauna inhabiting rivers and the inherent functioning of river ecosystems (sensu Southwood 1988). For instance, spatial-scale distribution patterns of flora and fauna are ultimately influenced by a habitat filtering dynamic based on this hierarchical perspective (Poff 1997), supporting ideas founded in niche theory (Thompson and Townsend 2006).
Human-dominated landscapes are a common feature on the globe (Meyer and Turner 1994). Consequently, rivers networking through these landscapes have been strongly influenced from recent human-based multiple stressors along with legacies of humans-past. Indeed, these land-use legacies may even confound our understanding of biological pattern and process in many riverscapes today (Harding et al. 1998; Allan 2004). For instance, response patterns of biota to river restoration measures may be limited by constraints from past land-use legacies. Present day land-use practices also may confound regional distribution patterns by overriding coarse-scale habitat properties influenced by, e.g. geologic differences between catchments (Bernot et al. 2010). This so-called landscape homogenization may weaken pattern and process among streams in a river network in response to land-use effects or gradients in environmental stressors (Matson et al. 1997; King et al. 2005), thus derived spatial patterns may reflect dispersal dynamics following ideas from neutral theory (Thompson and Townsend 2006) while not corresponding strongly to spatial patterns in environmental conditions. Because of the ever increasing dominance of humans in the landscape, it is even more imperative to better understand land-use effects on river structure and function to sustain the various services provided by these ecosystems.
Streams are often assessed in respect to abiotic and biotic responses to gradients in agricultural and urban land use (Richards et al. 1996; Strayer et al. 2003; Wassen et al. 2010). Four different response patterns are typically associated with gradients in environmental stress: a nonlinear threshold response at the high end of the gradient, a subsidy-stress response (e.g. Niyogi et al. 2007), a linear response, and a non-linear threshold response at the low end of the gradient (Allan 2004). The influence of multiple stressors is a current theme in many recent catchment studies as a mixture of juxtapositioned land uses are often common (Wagenhoff et al. 2011), potentially causing stressor nodes (hotspots) via local changes in water quality due to point-source and diffuse pollution, stream morphology, riparian vegetation or flow/temperature regimes (Matthaei et al. 2010). Urban areas in the catchment also show a disproportional effect on river ecosystems (Busse et al. 2006), especially in smaller basins where they can represent a larger percentage of the land cover (Allan 2004). Consequently, more ecological information is needed to elucidate the linkages and hydrological flow paths along river networks in human-dominated landscapes (e.g. Walsh and Kunapo 2009), as response patterns may differ substantially and be less clear than those in more pristine catchments.
Water quality is a primary factor affecting aquatic organisms through a range of different mechanisms. Urban and agricultural land-use may change the level of inorganic nutrients in streams, thereby influencing primary producers in the system. Degradation of organic matter may lead to low levels of dissolved oxygen. Concomitantly, toxic compounds like heavy metals and organic micropollutants (pesticides, pharmaceuticals, endocrine disruptors) may impair organisms at different levels in the aquatic food web. A primary goal of this study was to test the hypothesis that, within a catchment, coarse-scale spatial patterns in macroinvertebrate composition can be explained by differences in water quality and habitat properties as influenced by catchment land-use; essentially testing ideas of niche theory (e.g. habitat filtering) versus those of neutral theory (e.g. dispersal distance) (see Thompson and Townsend 2006). We predicted that niche theory dynamics would still dominate spatial patterns of aquatic macroinvertebrates in human-dominated catchments. For this purpose, we compared stream macroinvertebrate assemblages at different sites in two medium-sized catchments of the Swiss Plateau comprising a mixed land-use of forest, agriculture and urban areas (Gürbe, Mönchaltorfer Aa). Both catchments have been investigated in several previous studies that showed the stream networks to be affected by nutrients and other pollutants from agricultural and urban sources (Hanke et al. 2010; Wittmer et al. 2010, 2011; Stamm et al. 2012).
Methods
Catchment descriptions
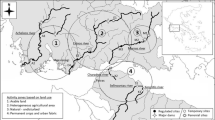
The Gürbe catchment is located south of Bern, Switzerland and covers an area of 137 km2. Altitude ranges from 2,176 m a.s.l. in the headwaters to ca. 500 m a.s.l. in the lowlands. The lower part is relatively flat and intensively used for arable cropping, and includes most urban areas in the catchment. The upper basin is characterized by steep slopes of partially karstic geology. Heavy rainfall in the upper catchment (1,260 mm average annual precipitation, 1,140 mm in the lower plain) often results in sudden floods, thus the entire main channel was heavily modified (channelized) during the 19th century mainly to reduce flood hazards in the lower valley. However, the lower section near the confluence with the Aare River has been morphologically improved in recent years. Land-use in the catchment consists of agriculture (57 %), forest (30 %), and urbanization (4 %). About 25,000 people live in the catchment, and treated wastewater is discharged into the main channel between sites G5 and G6 (Fig. 1). Treated wastewater of the village Belp further downstream has been discharged outside the catchment since 2009. Eight sampling sites were located along the Gürbe main channel and three along its primary tributary the Müsche (Fig. 1).

Map of the two study catchments (Gürbe, Mönchaltorfer Aa) in Switzerland (catchments in red) with site locations shown. Site G4, G7, and G8 are situated on the primary tributary of the Gurbe, the Müsche (color figure online)
The Mönchaltorfer Aa catchment covers an area of 46 km2 and is embedded in the Greifensee basin ca. 20 km southeast of Zürich, Switzerland in the Swiss Plateau (Fig. 1). Altitude ranges from 853 to 445 m a.s.l. and topography is moderate with headwater slopes typically being <15 %. Predominant soil types are cambisoles on hillsides and gley soils in flat areas. The climate is moderate with an average annual precipitation of 1,220 mm (20-year mean). The stream network consists of three main branches that merge a few kilometers upstream of the catchment outlet into Lake Greifen. The stream network was substantially modified during the 20th century with the lower reaches of streams being channelized during the process of land reclamation in the 1940s–1950s. Many small stream sections are now in culverts, while others are straight drainage channels with wooden boards forming the stream bottom. Land-use is mostly agriculture (63.2 %), although only ca. 20 % of this is used for arable farming of maize, winter wheat and barley. The most intensive agricultural area is near the outlet of the catchment. Forest covers about 16.7 % and urban areas about 10.9 % of the catchment. Fourteen sampling sites were located at various locations within the catchment to encompass the range of land uses (Fig. 1).
In both catchments, urban areas have combined and separated sewer systems. In combined sewer systems, urban storm-water and household wastewater is collected in the same sewer and directed to the WWTP. During intensive rainfall, combined sewer systems discharge excess water via combined sewer overflows to surface waters. In separated sewer systems, urban storm-water is collected separately and is discharged directly into surface waters.
Field and laboratory protocols
Water quality
Water grab samples (0.5 L) were collected from each site on a monthly basis from September/October 2010 to November 2011 (Gürbe, N = 10; Mönchaltorfer Aa, N = 12). Samples were returned to the laboratory on ice for analysis of dissolved organic carbon (DOC), particulate organic carbon (POC), alkalinity, total inorganic carbon (TIC), NH4-N, NO3-N, dissolved-N, particulate-N, PO4-P, dissolved-P, and particulate-P. Methods of analysis were the same as detailed in Tockner et al. (1997). On each sample date, spot measures of conductivity and temperature were taken using a field portable meter (WTW, Germany). Temperature also was recorded hourly at each site during the study using field-placed data loggers (Hobo tidbit). Some data gaps occurred in the logged temperature data and these were filled using a calibrated temperature model (M. Honti, unpubl. data).
Micropollutants, heavy metals, PCBs
Measures for micropollutants, heavy metals, and PCBs were collected at three sites in each catchment upstream and downstream of wastewater treatment plants (G1, G4, G5, M2, M3, M5, see Fig. 1). Two passive sampling campaigns were performed for organic, polar micropollutants in 2011 (Mönchaltorfer Aa: 31 May–17 June, 25 August–7 September; Gürbe: 6–20 June, 18 August–1 September) using preconditioned 3M Empore extraction disks (47 mm) (Vermeirssen et al. 2012) as sampling devices and a Supor-450 membrane filter as a protection cover and to reduce the uptake rate of samplers. Preconditioning consisted of placing disks in methanol in an ultra-sonic bath for 5 min, transferring to nano-pure water in an ultra-sonic bath for 5 min, and then storing in fresh nano-pure water. In the field, disks were placed along one transect in the stream, incubated for 2 weeks, collected and stored in acetone until processed. The subsequent analytical procedure was a multi-analyte LC-MSMS method as described in Singer et al. (2010).
Heavy metals and PCBs were assessed once in autumn 2011 (Mönchaltorfer Aa: 19 September–6 October; Gürbe: 20 September–5 October). For sampling heavy metals, DGT disks (DGT Research Ltd., Lancaster, UK) were deployed. Heavy metals were measured by ICP-MS after 24 h extraction in 2 M HNO3 following methods in Odzak et al. (2002). Pure silicon stripes were used for collecting PCBs, and time-averaged concentrations were estimated based on continuous water temperature measurements at each site (Zhang and Davison 1995).
Site characterization
Stream habitat characteristics at each 50-m study reach were assessed for percent substrate embeddedness, percent macrophyte cover, and percent canopy cover (shading). Sites also were characterized at baseflow for average wetted width (average of 5 transects), average substrate size (n = 50), water velocity at 0.6 depth (n = 50, MiniAir2, Schiltknecht Ag, CH), and average depth (n = 50). The abundance of emergent stones was categorized at each reach as well for estimates of ovipositioning substrata potentially used by adult aquatic insects.
Benthic samples
Benthic samples for macroinvertebrates were collected in autumn 2010 (October), spring 2011 (April) and summer 2011 (August) to account for seasonal changes in assemblage structure at each site in each catchment. Three samples were collected from riffle/run habitats at each site and date using a Hess sampler (0.048 m2, 250 μm mesh). Samples were preserved in the field with 75 % ethanol and returned to the laboratory for processing. In the laboratory, macroinvertebrates were handpicked from each sample using a microscope at 10× magnification. Taxa were identified to lowest practical level, usually family/genus, and counted. The remaining material from each sample was dried at 60 °C, weighed, combusted at 550 °C, and reweighed for determination of benthic organic matter as Ash-free Dry Mass (AFDM).
On each sample date for macroinvertebrates, five stones were randomly collected at each site for determination of periphyton biomass as chlorophyll a (Chl-a) and AFDM. Stones were returned to the laboratory in a cooler and frozen at −20 °C until processed. For processing, a known area was scraped from the surface of each stone using a metal brush. Two volumetric aliquots of the material were filtered (Whatman GFF filters), one filter was used for chlorophyll a analysis and the other for analysis of AFDM. Filters for chlorophyll were extracted in 90 % hot ethanol (70 °C) and analyzed using an HPLC. Filters for AFDM were dried at 60 °C, weighed, combusted at 550 °C, and reweighed with the difference in weights used to quantify the AFDM.
Data analysis
General habitat characteristics of each site were summarized as means and standard deviations. The physical–chemical measures of each site were treated separately for each catchment because of landscape-scale differences in catchment geology, land-use gradients, geography and climate. The water quality data for each site within a catchment were log(x + 1) transformed and then assessed using a principal components analysis (3-factor PCA with varimax rotation). The PCA was used to illustrate the spatial clustering of sites within each catchment based on similarities in water quality attributes. Relationships between land-use, habitat measures and PCA factor scores were analyzed using correlation (Zar 1984).
Each general environmental attribute (land-use, habitat, water quality) consisted of a set of single parameters (e.g. DOC, DON, TP, etc. for water quality). For each general attribute, the dissimilarity between sites was calculated as the Euclidian distance in multidimensional space where each coordinate consisted of single parameters normalized between zero and the maximum value in the catchment. The analysis was carried out with the statistics and graphics software R (R Development Core Team 2011). Subsequently, linear regressions were calculated between the dissimilarity values for the general environmental attributes between sites.
Macroinvertebrate data were summarized in terms of taxon richness, taxon and total density, and taxon relative abundance (percentage of the overall assemblage). Log(x + 1) transformed taxon density data and asin(sqrt(x)) transformed taxon relative abundance data for sites (and dates) within each catchment were analyzed using non-metric multi-dimensional scaling (NMDS). Results (site and date) were plotted as means (±SE) of sites along NMDS axis-1 and axis-2 for each catchment to illustrate spatio-temporal relationships in assemblage structure among sites within each catchment.
Lastly, relationships between macroinvertebrate densities and taxon richness against respective land-use, habitat and water quality (PCA results) attributes at sites within each catchment were assessed using correlation. In addition, the macroinvertebrate NMDS axis scores (based on taxa densities and taxa relative abundances) were correlated against respective land-use, habitat measures and the water quality PCA factor scores for the sites in each catchment. Finally, common macroinvertebrate taxa were averaged (±SE) for site groups within each catchment based on the water quality PCA results above with differences between groups tested using ANOVA on log (x + 1) transformed taxa density data (Zar 1984).
Results
Land-use and habitat characteristics in each catchment
In both catchments, study sites covered a wide range of land-use, in-stream habitat, and water quality characteristics (Table 1). Correlations between general environmental attributes (land-use, habitat, water quality) including the physical distance between study sites revealed different patterns for the two study areas (see Figs. S1, S2 in the supplementary material). Correlations among environmental attributes explained 50 % of the observed variance in the Gürbe, whereas the observed variance was only 21 % in the Mönchaltorfer Aa. This implies that two sites that are more distant in the stream network in the Gürbe are also dissimilar with regard to the environmental influence of land-use, habitat and water quality, whereas this distance effect is much less so in the Mönchaltorfer Aa (see detailed results below).
Gürbe
The percentage forest ranged from 12 % (G4) to 82 % (G10), although most sites had 20–30 % forest cover (Table 1). The percentage agriculture ranged from 17 % (G10) to 78 % (G4) with most sites having 50–60 % agricultural cover. The percentage of urban land-use ranged from zero (G10) to 6 % (G7) with most sites having 2–5 % urban cover. Most sites had relatively low values of percentage embeddedness (range 0–50 %), except for G4 at 75 % and G8 at 100 % (Table 1). The percentage macrophyte cover also ranged from 0 to 50 % at most sites, except G7 and G8 at 75 % coverage. The abundance of ovipositioning stones for adult aquatic insects was highest at sites G8–G11, with very few emergent stones (0–10 stones per reach) found at the other sites. Percent shading was <25 % at most sites except forested G10 with 100 %. Average stream width and depth reflected the general longitudinal location of the sites with width ranging from ca. 1 m at upper sites to 9 m at lower sites and depth from 5.7 cm at smaller sites to 65.3 cm at larger sites in the catchment. Average substrate size ranged from 2.0 to 9.5 cm, and flow velocities from 0.19 to 0.63 m s−1 (except G10 at 0.05 m s−1) between sites (Table 1).
Mönchaltorfer Aa
The percentage forest ranged from 8 % (M6) to 32 % (M14), although most sites had ca. 15 % forest cover (Table 1). The percentage agriculture ranged from 42 % (M14) to 83 % (M8) with most sites having ca. 65 % agriculture cover. The percentage urban cover ranged from zero (M11) to 19 % (M4) with most sites having between 8 and 15 % urban cover. Percentage embeddedness was relatively high in the catchment with most sites averaging >50 % and all sites being >25 % (Table 1). Percentage macrophyte cover was low, ranging from 0 to 25 % at most sites except M3 and M4 at >50 %. The percent shading was typically over 50 % with most sites being >75 %; an exception was G8 with 0 % shading. Average stream width reflected general stream size and ranged from 1 to 5 m. Average flow velocity and depth were quite similar among sites, ranging from 0.10 to 0.45 m s−1 and 5.4 to 19.3 cm, respectively.
General water quality conditions in each catchment
During baseflow conditions, as reflected in the grab samples, nutrient levels were generally low at most sites in both catchments. According to the Swiss Modular Concept for Stream Assessment (Bundi et al. 2000; Liechti 2010), most average concentrations for DOC, NH4-N, nitrate–N and DRP are classified as very good or good (DOC <4.0 mg L−1, NH4-N < 0.2 mg L−1, NO3-N < 5.6 mg L−1, DRP <0.04 mg L−1). Nutrient levels were high at a few sites, mostly those located downstream of WWTPs in the Mönchaltorfer Aa where the average NO3-N values of 12.2 mg L−1 at M5 are classified as poor.
Time averaged concentrations of measured toxicants were generally low. Heavy metal concentrations also were low (<1 μg L−1) (data not shown), ranging between 10.9 and 18.3 % of the water quality criteria in Switzerland (2.0 μg L−1) at the Gürbe sites and between 24.2 and 29.8 % in the Mönchaltorfer Aa. Cu and Zn exhibited the highest concentrations, with ranges for Zn being 3.5–11.6 % (Gürbe) and 22.4–30.8 % (Mönchaltorfer Aa). A comparison between sites upstream (G4, M5) and downstream (G1, G5, M2, M3) of WWTPs showed that WWTPs were not major sources of Cu or Zn. High values of Fe and Mn at G4 suggest naturally reduced sub-soils indicative of drained wetlands. PCB levels (10–250 ng kg−1 silicon in the Gürbe, 40–320 ng kg−1 silicon in the Mönchaltorfer Aa) were in the range of many Swiss streams without a pollution problem (M. Zenneg, pers. comm.). The toxic equivalent concentrations ranged between 0.055 ng kg−1 silicon (Gürbe: G1) and 0.208 ng kg−1 silicon (Mönchaltorfer Aa: M1), suggesting more of a regional than local effect. Because of the relatively low values, no further analysis was conducted for these parameters in this study.
Temporal and spatial distributions of organic micropollutants corresponded to expected patterns. Below WWTPs, for instance, indicator compounds for urban areas like human pharmaceuticals (e.g. carbamazepine: time-averaged concentrations of 12–14 ng L−1 in the Gürbe, 17–234 ng L−1 in the Mönchaltorfer Aa) and the anti-corrosive benzotriazole (24–55 ng L−1 in the Gürbe, 77–1226 ng L−1 in the Mönchaltorfer Aa) had high concentrations, but were present at very low levels at the other sites. Major herbicides like metolachlor and terbutylazine were present during the period of herbicide application at all sites with a substantial fraction of arable land use. Lastly, the urban source seemed more important for pesticides having a dual use (urban and agricultural) like diazinone (insecticide) and carbendazime (fungicide).
Environmental relationships within each catchment
Gürbe
The first three axes of the PCA results explained 80 % of the variation in measured water quality attributes between sites in the Gürbe (Fig. 2). The data revealed a clear spatial differentiation within the catchment. PCA axis-1 was best explained by values of electrical conductivity, alkalinity, TIC, NO3-N, and DN. PCA axis-2 was best explained by values of DOC, POC, PN, PP, PO4-P, and DP, whereas axis-3 was best explained by differences between sites in mean and maximum temperatures. Three groups were distinguished via the PCA results, reflecting lower sites along the main stream and G4 (Group 1), the Müsche tributary sites G7 and G8 (Group 2), and upper sites along the main stream (sites G9–G11) (Group 3).

Scatterplots of the principal components analysis based on the physical–chemical measures recorded for each site in the two catchments during the study period. Variables with factor loadings >0.70 are shown for each respective axis. G represents sites in the Gürbe catchment and M represents sites in the Mönchaltorfer Aa catchment
Examination of water quality attributes between groups illustrated the differences shown by the PCA (Table 2). For example, parameters explaining PCA axis-1 were notably lower in Group 3 sites in the upper catchment and increased downstream along the main stream (Group 1) and were highest in Müsche tributary sites (Group 2). For instance, the upper Gürbe is characterized by low alkalinity (mean 3.6 mmol L−1), moderate levels of conductivity (mean 423 μS cm−1) and low levels of TIC (mean 42.9 mg L−1) and nitrate–N (mean 0.57 mg L−1). In contrast, the lower Gürbe had higher mean values of alkalinity (4.8 mmol L−1), conductivity (523 μS cm−1), TIC (57.9 mg L−1) and nitrate–N (2.3 mg L−1), whereas the Müsche tributary had the highest mean values of alkalinity (6.2 mmol L−1), conductivity (547 μS cm−1), TIC (73.8 mg L−1), and nitrate–N (3.2 mg L−1).
PCA axis-2 was explained by values of particulates and phosphorus, with values being lower and somewhat similar between Groups 1 and 3, but much higher for the tributary sites of Group 2. For instance, the upper Gürbe had low levels of DOC (mean 2.4 mg L−1), and DP (mean 3.5 μg L−1), whereas downstream levels increased moderately in the main channel (mean DOC = 2.4 mg L−1, DP = 7.6 μg L−1) and were more than twofold higher in the Müsche tributary (mean DOC = 6.3 mg L−1, DP = 39.8 μg L−1). Lastly, PCA axis-3 was explained by temperature differences between groups that reflected the longitudinal position of sites in the main stream (lower temperatures in Group 3, higher temperatures in Group 1), but with similar temperatures in Group 2 sites as in Group 3 (Table 2).
Land-use attributes were significantly correlated with PCA axes 1 and 3, being negatively related to % forest (r = −0.82, −0.66, respectively) and positively with % agriculture (r = 0.94, 0.57, respectively) and % urban (r = 0.84, 0.85, respectively). Land-use also was significantly related to the algal biomass (AFDM, Chl-a) at the different sites, being negatively related to percentage forest (r = −0.72, −0.57, respectively) and positively related to % agriculture (r = 0.84, 0.73, respectively) and % urban (r = 0.77, 0.61, respectively). Lastly, percentage macrophyte cover was positively related to PCA axes 1 and 2 (r = 0.72, 0.75, respectively) as well as algal biomass (r = 0.68, 0.77, respectively) and benthic organic matter (r = 0.67).
Mönchaltorfer Aa
The first three axes of the PCA results explained 64 % of the variation in measured water quality attributes between sites in the Mönchaltorfer Aa (Fig. 2). PCA axis-1 was best explained by values of DN, NO3-N, and electrical conductivity. PCA axis-2 was best explained by values of DP, PO4-P and mean and maximum temperature, whereas axis-3 was best explained by values of POC, PN, and PP. The PCA results distinguished four groups of sites within the catchment. Group 1 included the lower basin sites M2, M3 and M5, all which are influenced by treated wastewater. Group 2 sites included M1, M4, M6 and M14, sites generally located in the middle region of the catchment. Group 3 sites included sites M7-M10, M12 and M13 (more upper basin sites), and Group 4 was site M11 (a small forested headwater stream).
Examination of water quality attributes between groups illustrated the differences shown by the PCA (Table 2). For instance, parameters explaining PCA axis-1 were highest in Group 1 sites, intermediate in Groups 2 and 3, and lowest for Group 4. Here, Group 1 sites had highest conductivities (mean 713 μS cm−1) and NO3-N (mean 8.9 mg L−1), whereas values were lowest for Group 4 (conductivity = 482 μS cm−1, NO3-N = 1.2 mg L−1). PCA axis-2 was best explained by values of phosphorus (PO4-P, DP) and temperature, where Group 4 had the lowest mean values (DP = 9.8 μg L−1, temperature = 9.9 °C), Group 2 had intermediate values, and Groups 1 and 3 had the highest mean values (DP = 30–33 μg L−1, temperature = 11.1–12.4 °C). PCA axis-3 reflected differences in particulates among the groups, where Group 2 had the lowest mean values for PP (4.0 μg L−1) and Groups 3 and 4 had highest values for POC (1.0–1.9 mg L−1). Notably, few major differences were found among groups along PCA axis-3 (see Fig. 2; Table 2).
The percentage forest land-use was negatively (r = −0.66) and % agriculture positively (r = 0.51) related to PCA axis-2 (i.e. phosphorus, temperature), whereas % macrophyte cover was positively related to PCA axis-1 (r = 0.63). There was a distinct water quality signal at sites downstream of WWTPs (e.g. Group 1 sites), in which conductivity ranged between 685 and 800 μS cm−1 while the other sites had values between 480 and 600 μS cm−1. Similarly, pH was slightly below 8.0 (7.7–7.9) for sites downstream of WWTPs but was generally >8.0 at the other sites. Lastly, NO3-N levels were substantially higher at sites below WWTPs (6.6–12.2 mg L−1 compared to 1.4–3.8 mg L−1), but little WWTP effect was observed for phosphorus.
Relationships between macroinvertebrates and environmental attributes
The differences in spatial structure among environmental attributes in each catchment also are reflected in how well the taxonomic dissimilarities between sites can be explained by environmental dissimilarity. In the Gürbe, land-use explained 54 % of the taxonomic dissimilarity between sites based on linear regression (see Fig. S3 in supplementary material). The explained taxonomic dissimilarity increased to 71 % when other environmental attributes were included in the analysis, although the most extreme dissimilarities between sites were not captured by the regression models. In the Mönchaltorfer Aa, the environmental dissimilarity between sites explained little of the taxonomic dissimilarity based on the regression models. Water quality alone accounted for 13 % of the taxonomic dissimilarity, and this percentage increased to 15 % when other environmental attributes were included (see Fig. S4 in supplementary material). Specific relationships between environmental attributes and macroinvertebrates in each catchment are presented below.
Gürbe
There was a positive correlation of total density and taxon richness of sites against water quality PCA axis-1 (r = 0.61, r = 0.44, respectively) (Fig. 3; Table 3). A highly significant correlation was found (r = 0.94, r = 0.95, respectively) if the tributary sites (G4, G7, G8) were excluded in the analysis. These tributary sites showed lower densities and taxon richness relative to the PCA axis scores than main river sites. In main river sites, densities and taxon richness were higher in lower basin sites than in upper basin sites, reflecting the higher values of conductivity, alkalinity, TIC, NO3-N, and DN (i.e. PCA axis-1) in the lower basin sites.
The correlation of total density and taxon richness also was positive against PCA axis-3 (defined as mean and maximum temperature) (r = 0.85, r = 0.55, respectively; Fig. 3; Table 3). The relationship became more positive when the tributary sites (G4, G7, G8) were excluded, especially in respect to taxon richness (r = 0.90 for density, r = 0.96 for taxon richness). In this case, lower main stream sites had warmer temperatures than upper basin sites and, in turn, had higher densities and taxon richness. In contrast, the tributary sites showed no relationship of taxon richness with temperature (Fig. 3). Lastly, % forest land-use was negatively (r = −0.61) and the % agriculture/urban positively (r = 0.54, 0.69, respectively) related to total density.

Scatterplots of significant regressions of macroinvertebrate total densities and taxon richness against principal component scores from the physical–chemical analysis shown in Fig. 2 for the respective sites in each catchment (G Gürbe, M Mönchaltorfer Aa)
The NMDS analysis (which incorporated time as well) using taxon densities and taxon relative abundances separated sites along the first two axes with stress levels of 0.18 and 0.13, respectively (data not shown). There was a strong correlation of NMDS axis-1 site scores against water quality PCA axis-1 site scores for taxon density (r = −0.86) and taxon relative abundances (r = 0.90) (Fig. 4; Table 3). The correlation of NMDS axis-2 site scores against water quality PCA axis-2 site scores was poor (r = −0.22 for taxon density, r = 0.48 for taxon relative abundance), but clearly showed the tributary sites G7 and G8 to differ from the main stream sites along PCA axis-2.

Scatterplots of site scores from the NMDS analysis based on macroinvertebrate assemblage structure (taxon densities and taxon relative abundances) in each catchment against respective principal component scores from the physical–chemical analysis (G Gürbe, M Mönchaltorfer Aa). NMDS-1 is axis-1 site scores from the NMDS analysis and NMDS-2 is axis-2 site scores from the NMDS analysis. PCA-1 is axis-1 site scores and PCA-2 axis-2 site scores from the physical–chemical principal components analysis shown in Fig. 2
The NMDS axis-1 scores (taxon densities and relative abundances) were related to the % land-use (% forest: r = −0.59 for density, r = 0.67 for relative abundance; % agriculture: r = 0.76, r = −0.78, respectively; % urban: r = 0.75, r = −0.87, respectively) (Table 3). Lastly, NMDS axis-1 scores showed either a negative (taxon relative abundances: r = −0.70 AFDM, r = −0.56 Chl-a) or positive relationship (taxon densities: r = 0.85 AFDM, r = 0.80 Chl-a) with algal biomass.
There were some significant differences in the density of different taxa among site groups in the Gürbe (Fig. 5). Trichopterans had higher densities in Group 2 (G7, G8) than the other two groups. Plecopterans and simuliids had higher densities in Group 3 (G9–G11) than the other two groups, whereas, leptophlebiids, chironomids and dipterans (excluding simuliids and chironomids) were highest in Group 1 (G1–G6). Gastropods, coleopterans, hirudinids, turbellarians, and gammarids were highest in Groups 1 and 2, but absent or low in density in Group 3. Lastly, heptageniids were abundant in Groups 1 and 3, but low in density in Group 2, whereas differences for acari, baetids and rhyacophilans were not significant.

Average (+1 SE) densities of macroinvertebrate taxa collected in each catchment for site groups distinguished from the physical–chemical principal components analysis in Fig. 2. Site groups are described in Table 2 with three groups being distinguished in the Gürbe and four groups distinguished in the Mönchaltorfer Aa. Asterisks show taxa with significant density differences among groups in each catchment (ANOVA, p < 0.05). TRIC trichopterans, RHYA rhyacophilans, PLEC pleclopterans, ACAR hydracarina, GAMM gammarids, LEPT leptophlebiids, HEPT heptageniids, BAET baetids, TURB turbellarians, HIRU hirudinids, COLE coleopterans, DIPT dipterans, SIMU simuliids, CHIR chironomids, GAST gastropods, EPHE ephemeropterans, BIVA bivalves, ASSE asselids
Mönchaltorfer Aa
There was a positive correlation of total density and taxon richness against water quality PCA axis-1 (r = 0.66, r = 0.53, respectively) (Fig. 4; Table 3). Sites M2, M3, and M5 had higher values of NO3-N, DN, and electrical conductivity that was reflected in higher total densities than other sites in the basin. The relationship of PCA axis-1 with taxon richness was less pronounced, although M2 and M3 had higher taxon richness values and higher PCA factor scores than the other sites but M5 had a relatively low taxon richness. The correlation of total density and taxon richness against PCA axis-2 (defined by PO4-P, DP, temperature) was poor (r = 0.35, r = 0.07, respectively) (data not shown). Total density and taxon richness were positively related to the % macrophyte cover at a site (r = 0.56, r = 0.54, respectively).
The NMDS analysis (which incorporated time as well) using taxon densities and taxon relative abundances separated sites along the first two axes with stress levels of 0.20 and 0.19, respectively (data not shown). There was a negative relationship of NMDS axis-1 site scores against water quality PCA axis-1 site scores for taxon density (r = −0.59) and taxon relative abundance (r = −0.46) (Fig. 4). Here, sites M2, M3, and M5 (lower basin sites) had higher values of NO3-N, DN, and conductivity but lower taxon density and taxon relative abundance scores than the other sites, especially M12, M13, and M14 (the most upper basin sites). The correlation of NMDS axis-2 site scores against water quality PCA axis-2 site scores showed no relationship (r = 0.28 for taxon relative abundance, r = −0.11 for taxon density), except site M11 had low values for each (Table 3).
The % forest land-use was positively related to both NMDS axis-1 scores (taxon relative abundances: r = 0.60, taxon densities: r = 0.55), whereas the NMDS axis-1 scores for taxon relative abundances were negatively related to % agriculture land-use (r = −0.51) (Table 3). Lastly, NMDS axis-1 scores for taxon densities were negatively related to algal AFDM (r = −0.60), whereas both NMDS axis-1 scores (taxon densities and relative abundances) were positively related to benthic organic matter (r = 0.75, r = 0.44, respectively) and % shading (r = 0.70, r = 0.49, respectively).
In the Mönchaltorfer Aa (Fig. 5), ephemeropterans, acari, and coleopterans had high densities in Groups 1–3, but low densities in Group 4 (M11). Gastropods and chironomids had highest densities in Group 1 (M2, M3, M5), although chironomids were also abundant in the other groups. Dipterans excluding chironomids were most abundant in Groups 1 and 2 (M1, M4, M6, M14). Plecopterans showed a trend of higher densities in Groups 2 and 3 (M7–M10, M12–13) than Groups 1 and 4. The other examined taxa (trichopterans, gammarids, bivalves, turbellarians, hirudinids, asselids) showed similar densities between groups.
Discussion
This study examined the spatial relationships between land-use, habitat, water quality, and stream macroinvertebrates in two human-dominated catchments. The results revealed contrasting findings for the two catchments; i.e. a catchment dependent context. In the Gürbe catchment, where environmental attributes were closely correlated, macroinvertebrate assemblages reflected coarse-scale patterns in water quality as influenced by local land-use. Specifically, the Gürbe catchment was dominated by agricultural land-use and displayed a strong longitudinal gradient in biotic response along the main channel with a small primary tributary showing a more pronounced agricultural land-use effect. Whereas the Gürbe is dominated by the main channel in hydrologic terms, the Mönchaltorfer Aa catchment has a more complex stream network of three main branches of similar size (see Fig. 1). Reflecting this more complex stream network, there also was a more complicated spatial relationship among environmental attributes punctuated by areas of urban land-use that influenced biotic patterns in the landscape. Here, more localized response patterns reflected inputs from urban areas, e.g. WWTPs, and outlier sites in more forested headwaters. Results from both catchments suggest niche theory dynamics dominate observed macroinvertebrate spatial patterns as discussed further below.
Relationships between land use and stream physical-chemistry
In many Swiss lowland catchments, streams typically originate in managed, forested headwaters before flowing through a landscape dominated by agricultural lands and urban settings. Humans have dominated European landscapes for 1,000s of years and this historical legacy of land-use may be evident in many human-dominated catchments today. Most streams have been regulated and channelized to reduce flood hazards and to optimize the area available for agricultural use. Such major land-use changes affecting the stream network occurred mainly during the last 120 years (Gürbe) and over the last 60 years (Mönchaltorfer Aa) in our study areas. Thus the potential for legacy effects on present-day response patterns is probable. Many smaller streams, e.g. 1st order tributaries, in the agricultural and urban landscape also are in culverts, thus simplifying the river network in such landscapes and influencing the overall aquatic biodiversity. We thus expected surface flowing waters to show a strong land-use effect in such human-dominated catchments that ultimately would be reflected in macroinvertebrate spatial patterns.
The Gürbe catchment had a highly simplified network with most sites situated along the main channel (see Fig. 1). The main river had one major tributary (Müsche) with three sites (G4, G7, G8) being sampled for this study. Water quality changes in this system reflected a strong longitudinal dynamic as the gradient of influence from agricultural lands increased downstream. A number of studies have shown a decrease in habitat quality in streams flowing through agricultural landscapes resulting from increases in nutrients and sediments (Richards et al. 1996; Quinn 2000). For instance, electrical conductivity and nitrogen constituents increased downstream and these changes were highly correlated with the percentage agricultural land in the Gürbe. This longitudinal dynamic also was observed in an increase in water temperature from the most headwater sites to the lower basin sites, an expected continuum characteristic of more natural river networks (sensu Vannote et al. 1980), although the degree of increase is likely related to changes in land-use. For instance, water temperature was negatively related to % forest and positively with % agricultural land-use, suggesting the loss of forest and gain in agricultural land-use may have a synergistic effect with longitudinal location on water temperature because of reduced shading by riparian vegetation in the agricultural part. The results suggest that spatial patterns in water quality in the Gürbe followed a general linear response as streams flowed from the forested headwaters to lower basin sites (after Norris and Thoms 1999), reflecting the strong longitudinal nature of the river network in the catchment.
The Gürbe tributary sites G7, G8 deviated from the main channel sites, demonstrating a more localized influence from agricultural land-use on smaller streams. Although being smaller in terms of flow than the main channel sites, water quality at these sites was similar and even lower than that at lower basin sites along the main channel (see Table 2). For example, nutrient concentrations were the highest at these two sites, phosphorus in particular, and carbon measures (POC, DOC, TIC) were double that of the other sites. These two sites also had the highest percentage agricultural land-use and the highest percentage macrophyte cover. Macrophyte cover tends to be higher in agricultural streams due to the increase in nutrients and better light conditions from a more open canopy (Allan and Castillo 2007). In summary, these tributary sites showed a strong influence of local land-use on water quality that overrides regional or more coarse-scale land-use effects. The spatial context of riverine networks in relation land-use is complex, but clearly multiple spatial scales must be embedded towards a holistic understanding of riverine structure and function (Allan 2004). The results from these tributary sites displayed a more subsidy-stress response pattern (Matthaei et al. 2010), and suggest a mix of response patterns may be evident within a catchment depending on the spatial scale of analysis (Richards et al. 1996).
The Mönchaltorfer Aa catchment, in contrast, had a much more complex river network with sites being sampled on three of its major tributaries. Urban land-use also was embedded in the landscape and represented a significant percentage of the catchment (10.9 vs 3.6 % in the Gürbe). Most studies have shown significant urban land-use effects at percentages ca. 10 %, and there is a basic tenant that substantially less percentage urban land-use, e.g. the 10 % threshold, can have significant impacts on water quality than that from agricultural lands (ca. 30–50 % on average) (Allan 2004). However, the urban footprint was more localized in the catchment, e.g. downstream of WWTPs inputs, probably because populations in the villages were on the order of 10s of thousands and not major urban centers. The relatively low gradient within this catchment perhaps facilitated a more homogenized land-use effect from intensive agriculture, although punctuated with urban effects at some sites (e.g. M5).
The coarse-scale longitudinal pattern observed in the Gürbe was less evident in the Mönchaltorfer Aa, although upper basin sites in the catchment had better water quality than lower-most basin sites (e.g. M2, M3, M5; see PCA results Fig. 2). Here, no correlation was observed between the percentage agricultural land-use and water quality differences among sites, further suggesting a spatial homogenization of stream physicochemical features in the catchment. The extensive agricultural setting in the catchment, on average 10 % higher than in the Gürbe, offset any longitudinal patterns expected in physicochemical features, e.g. water temperature and conductivity, resulting from an increasing longitudinal gradient in land-use change. Indeed, only one site situated in a forested headwater (M11) had water quality conditions indicative of more natural landscapes; i.e. lower water temperature and electrical conductivity typical for the geologic region. Indeed, water quality patterns in the Mönchaltorfer Aa displayed a non-linear response that reflected the more homogeneous landscape properties in this catchment (Allan 2004). Water quality changed dramatically once streams flowed out of the forested headwaters and then water quality remained rather similar throughout the lower basin.
Spatial determinants affecting macroinvertebrate assemblages
Macroinvertebrate assemblages in the Gürbe catchment were strongly related with changes in environmental conditions among sites. Here, total densities, taxon richness and NMDS site scores showed significant correlations with the physico-chemical PCA scores. Taxonomic dissimilarity as assessed by NMDS also was well-explained by land-use and the different combinations of (correlated) environmental attributes. These results showed that macroinvertebrate assemblages responded in a relatively linear fashion with the gradient in environmental conditions among sites, as reflected in the strong longitudinal pattern evident in this basin. These findings support ideas from niche theory inferring the importance of habitat filtering in species distributions (after Poff 1997). The tributary sites (G4, G7, G8) deviated from the main channel sites, and indicated a less diverse assemblage in the tributary due to the more pronounced degradation in water quality. These results are not overly surprising as the tributary is strictly used as a drainage canal for adjacent agricultural lands; site G8 even has wooden boards installed on the stream-bed to facilitate drainage. Similar spatial-scale effects on stream habitat conditions have been noted in other studies (e.g. Sponseller et al. 2001), further confirming the general hierarchical habitat-structuring of streams embedded in the landscape (sensu Frissell et al. 1986).
Macroinvertebrate assemblages in the Gürbe were related to percentage land-use in the catchment, in particular agricultural and urban land-use. The relationship with urban land-use is likely a spurious result as most urban center inputs were downstream of the sampling sites and urban land-use was strongly correlated with agricultural land-use in the catchment. This kind of relationship has been thoroughly discussed in the literature regarding inter-related land-use effects in aquatic systems (see Allan 2004). Furthermore, average urban land-use in the catchment was low (3.9 %), and most studies document urban land-use effects at percentages higher than 10 %. Consequently, the relationship of macroinvertebrate assemblages with percentage agricultural land-use is more realistic as this land-use is the predominant land-use in the catchment (see Table 1). Support for this argument also is demonstrated in the strong relationship of algal biomass measures with % agricultural land-use, showing the generally positive relationship of primary producers with increases in nutrients flowing off agricultural lands (Carr et al. 2005; Klose et al. 2012).
Specific macroinvertebrate taxa responded to the longitudinal gradient in land-use effect on water quality. The upper basin sites had cooler water temperatures and better water quality than lower basin sites and this was reflected in the greater abundance of plecopterans, a group well-known to be sensitive to warmer stream temperatures and poor lotic habitat conditions. In contrast, chironomids, gammarids, gastropods, hirudinids (leeches) and other dipterans (mainly empedids and ceratopogonids) were most common in the lower basin and tributary sites, reflecting the slightly lower water quality at these sites. Coleopterans such as dytiscids were also prevalent in lower basin sites, likely due to the presence of slower flowing pools. Lastly, trichopterans (mostly hydropsychids and hydroptilids) were more prevalent in the tributary sites G7 and G8. The filter-feeding hydropsychids were probably responding to the high concentrations of suspended particles, whereas the abundance of algal-piercing hydroptilids was likely related to the presence of macrophytes and associated epiphytes (Merritt and Cummins 1984). Richards et al. (1997) also showed the importance of catchment and reach-scale habitat properties in determining species assemblages and species traits within landscapes (also see Liess et al. 2012). These changes in community composition further support the habitat filtering perspective from niche theory (Thompson and Townsend 2006), as habitat conditions dictated the presence and absence of specific taxa among the sites.
The spatial homogenization in the Mönchaltorfer Aa also was evident in the lack of a land-use relationship with macroinvertebrate assemblage composition among sites. Only the % forest land-use showed a relationship with macroinvertebrate assemblages among sites, this being attributed to the more forested headwater sites in the catchment. This result was supported by significant relationships between the % shading (canopy cover) and benthic organic matter with macroinvertebrate assemblage composition among sites (see Table 3). These findings also confirm the non-linear response pattern of water quality with the gradient in land-use. The forested headwater sites clearly represent an important habitat type in the catchment with potential implications regarding biodiversity distribution patterns. The presence and absence of taxa among sites in this catchment were not related to spatial distances between sites as expected from neutral theory predictions regarding dispersal limitations on species distributions, thus lending additional support for niche theory in this catchment as well.
The lower basin sites (M2, M3, M5) showed the poorest water quality of all sites, mainly reflecting the presence of treated wastewater in the stream, which was absent at all other sites. These three sites were the primary cause in the positive relationship of macroinvertebrate assemblage structure, i.e. density, with changes in water quality, and M5 was located just downstream of the mixing point of a WWTP discharge into the stream. Walsh and Kunapo (2009) showed the importance of urban flow paths on the effects of urban areas on streams. Urbanized flow paths may demonstrate more localized effects on stream ecosystems, analogous to a site distance relationship (Mykrä et al. 2007; Walsh and Kunapo 2009). These data further suggest that even in homogenized catchments, the local influence of habitat quality plays an important role in dictating the local composition and abundance of macroinvertebrate assemblages as inferred from niche theory (Thompson and Townsend 2006).
The abundances of specific macroinvertebrate taxa supported the results in land-use and water quality in the Mönchaltorfer Aa catchment with an overall similar composition occurring at the sites. This finding supports the general premise of a homogenization of the landscape and its ultimate effect on macroinvertebrate diversity patterns. Although not tested, landscape homogenization in the catchment also may reflect legacy effects that reduced the overall species pool available for colonization. Legacy effects have been documented in other studies relating changes in land-use with the structure of macroinvertebrate assemblages (Harding et al. 1998; Brierley et al. 1999). The degraded lower basin sites showed the most significant differences among sites, being dominated by chironomids, gastropods and baetid mayflies. The forested sites showed some separation in ordination space, but the regional species pool likely constrained any major differences from the other sites, and gammarids and chironomids also dominated samples collected at this site. In closing, results from both catchments suggest niche theory dynamics dominate spatial distribution patterns of macroinvertebrates in these running waters. Historical legacies clearly leave a strong footprint on the regional species pool (after Dodds 2008), but the pool of available species still responds to the habitat filtering from regional and local environmental conditions.
References
Allan JD (2004) Landscapes and riverscapes: the influence of land use on stream ecosystems. Annu Rev Ecol Evol Syst 35:257–284
Allan JD, Castillo MM (2007) Stream ecology: structure and function of running waters, 2nd edn. Springer, The Netherlands
Allan JD, Erickson DL, Fay J (1997) The influence of catchment land use on stream integrity across multiple spatial scales. Freshw Biol 37:149–161
Amoros C, Bornette G (2002) Connectivity and biocomplexity in waterbodies of riverine floodplains. Freshw Biol 47:761–776
Bernot MJ, Sobota DJ, Hall RO, Mulholland PJ, Dodds WK, Webster JR, Tank JL, Ashkenas LR, Cooper LW, Dahm CN, Gregory SV, Grimm NB, Hamilton SK, Johnson SL, McDowell WH, Meyer JL, Peterson B, Poole GC, Valett HM, Arango C, Beaulieu JJ, Burgin AJ, Crenshaw C, Helton AM, Johnson L, Merriam J, Niederlehner BR, O’Brien JM, Potter JD, Sheibley RW, Thomas SM, Wilson K (2010) Inter-regional comparison of land-use effects on stream metabolism. Freshw Biol 55:1874–1890
Brierley GJ, Cohen T, Fryirs K, Brooks A (1999) Post-European changes to the fluvial geomorphology of Bega catchment, Australia: implications for river ecology. Freshw Biol 41:839–848
Bundi U, Peter A, Frutiger A, Hütte M, Liechti P, Sieber U (2000) Scientific base and modular concept for comprehensive assessment of streams in Switzerland. Hydrobiologia 422(423):477–487
Busse LB, Simpson JC, Cooper SD (2006) Relationships among nutrients, algae, and land use in urbanized southern California streams. Can J Fish Aquat Sci 63:2621–2638
Carr GM, Chambers PA, Morin A (2005) Periphyton, water quality, and land use at multiple spatial scales in Alberta rivers. Can J Fish Aquat Sci 62:1309–1319
Dodds WK (2008) Humanity’s footprint: momentum, impact, and our global environment. Columbia University Press, New York
Frissell CA, Liss WJ, Warren CE, Hurley MD (1986) A hierarchical framework for stream habitat classification: viewing streams in a watershed context. Environ Manage 12:199–214
Hanke I, Wittmer IK, Bischofberger S, Stamm C, Singer H (2010) Relevance of urban glyphosate use for surface water quality. Chemosphere 81:422–429
Harding JS, Benfield EF, Bolstad PV, Helfman GS, Jones EBD (1998) Stream biodiversity: the ghost of land use past. Proc Nat Acad Sci USA 95:14843–14847
King RS, Baker ME, Whigham DF, Weller DE, Jordan TE, Kazyak PF, Hurd MK (2005) Spatial considerations for linking watershed land cover to ecological indicators in streams. Ecol Appl 15:137–153
Klose K, Cooper SD, Leydecker AD, Kreitler J (2012) Relationships among catchment land use and concentrations of nutrients, algae, and dissolved oxygen in a southern California river. Freshw Sci 31:908–927
Liechti P. (2010) Methoden zur Untersuchung und Beurteilung der Fliessgewässer. Chemisch-physikalische Erhebungen, Nährstoffe. Umwelt-Vollzug Nr. 1005. Bundesamt für Umwelt, Bern, Switzerland
Liess A, Le Gros A, Wagenhoff A, Townsend CR, Matthaei CD (2012) Landuse intensity in stream catchments affects the benthic food web: consequences for nutrient supply, periphyton C: nutrient ratios, and invertebrate richness and abundance. Freshw Sci 31:813–824
Matson PA, Parton WJ, Power AG, Swift MJ (1997) Agricultural intensification and ecosystem properties. Science 277:504–509
Matthaei CD, Piggitt JJ, Townsend CD (2010) Multiple stressors in agricultural streams: interactions among sediment addition, nutrient enrichment and water abstraction. J Appl Ecol 47:639–649
Merritt RW, Cummins KW (1984) An introduction to the aquatic insects of North America. Kendall/Hunt Publishing Company, Iowa
Meyer WB, Turner BL (1994) Changes in land use and land cover: a global perspective. Cambridge University Press, New York
Mykrä H, Heino J, Muotka T (2007) Scale-related patterns in the spatial and environmental components of stream macroinvertebrate assemblage variation. Glob Ecol Biogeogr 16:149–159
Niyogi DK, Koren M, Arbuckle CJ, Townsend CR (2007) Stream communities along a catchment land-use gradient: subsidy-stress responses to pastoral development. Environ Manage 39:213–225
Norris RH, Thoms MC (1999) What is river health? Freshw Biol 41:197–209
Odzak N, Kistler D, Xue HB, Sigg L (2002) In situ trace metal speciation in a eutrophic lake using the technique of diffusion gradients in thin films (DGT). Aquat Sci 64:292–299
Poff NL (1997) Landscape filters and species traits: towards mechanistic understanding and prediction in stream ecology. J N Am Benthol Soc 16:391–409
Quinn JM (2000) Effects of pastoral development. In: Collier KJ, Winterbourn MJ (eds) New Zealand stream invertebrates: ecology and implications for management. Caxton, Christchurch, pp 208–229
R Development Core Team (2011) R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. ISBN 3-900051-07-0, http://www.R-project.org/
Richards C, Johnson LB, Host GE (1996) Landscape-scale influences on stream habitats and biota. Can J Fish Aquat Sci 53:295–311
Richards C, Haro RJ, Johnson LB, Host GE (1997) Catchment and reach-scale properties as indicators of macroinvertebrate species traits. Freshw Biol 37:219–230
Singer H, Jaus S, Hanke I, Lück A, Holleder J, Alder AC (2010) Determination of biocides and pesticides by on-line solid phase extraction coupled with mass spectrometry and their behaviour in wastewater and surface water. Environ Pollut 158:3054–3064
Southwood TRE (1988) Tactics, strategies, and templets. Oikos 52:3–18
Sponseller RA, Benfield EF, Valett HM (2001) Relationships between land use, spatial scale and stream macroinvertebrate communities. Freshw Biol 46:1409–1424
Stamm C, Ochsenbein U, Berset JD, Siber R, Scheiwiller E, Singer H, Muff D (2012) Ereignisbezogenes Pestizidmonitoring am Beispiel der Gürbe (Kanton Bern). Aqua Gas 4:24–32
Strayer DL, Beighley RE, Thompson LC, Brooks S, Nilsson C, Pinay G, Naiman RJ (2003) Effects of land cover on stream ecosystems: roles of empirical models and scaling issues. Ecosystems 6:407–423
Thompson R, Townsend CR (2006) A truce with neutral theory: local deterministic factors, species traits and dispersal limitation together determine patterns of diversity in stream invertebrates. J Anim Ecol 75:476–484
Tockner K, Malard F, Burgherr P, Robinson CT, Uehlinger U, Zah R, Ward JV (1997) Characteristics of channel types in a glacial floodplain ecosystem (Val Roseg, Switzerland). Archiv für Hydrobiologie 140:433–463
Vannote RL, Minshall GW, Cummins KW, Sedell JR, Cushing CE (1980) The river continuum concept. Can J Fish Aquat Sci 37:130–137
Vermeirssen ELM, Dietschweiler C, Escher BI, Van Der Voet J, Hollender J (2012) Transfer kinetics of polar organic compounds over polyethersulfone membranes in the passive samplers pocis and chemcatcher. Environ Sci Technol 46:6759–6766
Wagenhoff A, Townsend CR, Phillips N, Matthaei CD (2011) Subsidy-stress and multiple-stressor effects along gradients of deposited fine sediment and dissolved nutrients in a regional set of streams and rivers. Freshw Biol 56:1916–1936
Walsh CJ, Kunapo J (2009) The importance of upland flow paths in determining urban effects on stream ecosystems. J N Am Benthol Soc 28:977–990
Wassen JG, Villeneuve B, Iital A, Murray-Bligh J, Dobiasova M, Bacikova S, Timm H, Pella H, Mengin N, Chandesris A (2010) Large-scale relationships between basin and riparian land cover and the ecological status of European rivers. Freshw Biol 55:1465–1482
Wittmer IK, Bader HP, Scheidegger R, Singer H, Lück A, Hanke I, Carlsson C, Stamm C (2010) Significance of urban and agricultural land use for biocide and pesticide dynamics in surface waters. Water Res 44:2850–2862
Wittmer IK, Bader HP, Scheidegger R, Singer H, Stamm C (2011) Loss rates of urban biocides can exceed those of agricultural pesticides. Sci Total Environ 409:920–932
Zar JH (1984) Biostatistical analysis, 2nd edn. Prentice Hall, Englewood Cliffs
Zhang H, Davison W (1995) Performance characteristics of diffusion gradients in thin films for the in situ measurement of trace metals in aqueous solution. Anal Chem 67:3391–3400
Acknowledgments
The authors graciously thank C. Jolidon, A. Lück, M. Scheurer and N. Ghielmetti for assistance in the field and laboratory processing of samples. We thank the Aua Lab of Eawag for completing the water chemistry analyses. A. Lück and H. Singer carried out the analyses of the organic micropollutants, D. Kistler analysed the heavy metal measurements and M. Zennegg the PCBs. E. Vermeirssen and N. Odzak provided valuable support for the passive sampling. We thank R. Siber for GIS support and map illustrations. We thank two anonymous reviewers that provided helpful comments that improved the paper presentation. The project was funded by the Swiss National Science Foundation (No. 4061-40_125866/1).
Author information
Authors and Affiliations
Corresponding author
Electronic supplementary material
Below is the link to the electronic supplementary material.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Fig. S1: Correlation between dissimilarities of environmental factors between sites in the Gürbe catchment.
Fig. S2: Correlation between dissimilarities of environmental factors between sites in the Mönchaltorfer Aa catchment.
Fig. S3: Comparison of the observed taxonomic distance between sites (x-axis) and the predicted values (y-axis) based on linear models for all combinations of environmental factors in the Gürbe catchment. The black points represent the full model including all environmental factors, red points represent the 3-factor models, blue the 2-factor models, and green the single factor models. Tax: taxonomic distance in the Gürbe, Dist: physical distance in the stream network Hab: Habitat, qual: dist:, Use: land use, WQ: water quality.
Fig. S4: Comparison of the observed taxonomic distance between sites (x axis) and the predicted values (y axis) based on linear models for all combinations of environmental factors in the Mönchaltorfer Aa catchment (all 14 sites). The black points represent the full model including all environmental factors, red points represent the 3-factor models, blue the 2-factor models, and green the single factor models. Tax: taxonomic distance in the Gürbe, Dist: physical distance in the stream network Hab: Habitat, qual: dist:, Use: land use, WQ: water quality.
Rights and permissions
About this article
Cite this article
Robinson, C.T., Schuwirth, N., Baumgartner, S. et al. Spatial relationships between land-use, habitat, water quality and lotic macroinvertebrates in two Swiss catchments. Aquat Sci 76, 375–392 (2014). https://doi.org/10.1007/s00027-014-0341-z
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00027-014-0341-z