
Abstract
We explored the strength of connectedness among hierarchical system components associated with oxbow lakes in the alluvial valley of the Lower Mississippi River. Specifically, we examined the degree of canonical correlation between land use (agriculture and forests), lake morphometry (depth and size), nutrients (total nitrogen and total phosphorus), primary production (chlorophyll-a), and various fish assemblage descriptors. Watershed (p < 0.01) and riparian (p = 0.02) land use, and lake depth (p = 0.05) but not size (p = 0.28), were associated with nutrient concentrations. In turn, nutrients were associated with primary production (p < 0.01), and primary production was associated with sunfish (Centrarchidae) assemblages (p < 0.01) and fish biodiversity (p = 0.08), but not with those of other taxa and functional guilds. Multiple chemical and biological components of oxbow lake ecosystems are connected to landscape characteristics such as land use and lake depth. Therefore, a top-down hierarchical approach can be useful in developing management and conservation plans for oxbow lakes in a region impacted by widespread landscape changes due to agriculture.
Similar content being viewed by others
Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
A predominant paradigm in aquatic ecology is the connectedness among ecosystem components (Ward 1998; Wiens 2002). Briefly, the paradigm instructs that aquatic systems are linked to their watersheds through nutrient inputs, that nutrients determine algal composition and biomass, and that algal biomass affects the structure and function of aquatic invertebrate and vertebrate communities. The paradigm considers fluvial landscapes as multi-scaled nested hierarchies of interactive terrestrial and aquatic elements (Frissell et al. 1986; Townsend 1996). Hence, changes in terrestrial landscapes can cascade through aquatic ecosystems. However, intensity of the resonance will likely depend on local factors such as a system’s assimilative capacity (e.g., lake size and depth) and interactions with local features (e.g., landscape peculiarities, human populations, management interventions).
Algae are a major biotic component directly affected by increases in nutrients. The primary nutrients exported from the watershed that determine algal composition and biomass are phosphorus and nitrogen (Frost et al. 2009). The importance of N and P in regulating algal growth varies geographically, seasonally, and among types of aquatic systems (Elser et al. 1990). Algal density and activity can influence key water physicochemical features such as temperature and dissolved oxygen (Wetzel 2001). Also, as a primary producer, algae support secondary and tertiary production, including zooplankton, macroinvertebrates, and fish (Rader and Richardson 1992; Vadeboncoeur et al. 2001). Chlorophyl-a has been linked to fish yield in lakes and reservoirs, and the relationship between fish and chlorophyll-a is reportedly stronger than that between fish and nutrients (Jones and Hoyer 1982). Thus, the connectedness paradigm specifies that system components can serve both as response variables and as causal factors (e.g., nutrients respond to watersheds, and more nutrients increase algal biomass).
On the basis of this paradigm, various options are available as response variables to monitor the health of aquatic systems (Jørgensen et al. 2005). Accordingly, researchers have used responses such as nutrient levels, algal biomass, macroinvertebrate assemblages, and fish assemblages to monitor status of rivers, lakes, and reservoirs (Niemi et al. 1990). Choice of the response variable can be made based on sensitivity to change, ease or cost of measurement and analysis, relevance to human use, and the strength of the connectedness between successive stages in the cascade.
We explored the strength of the relationships among hierarchical system components in response to the need to establish nutrient criteria, and to establish their relevance to the functioning of oxbow lake ecosystems in the alluvial valley of the Lower Mississippi River. Specifically, we examined the connectedness between large-scale environmental features and nutrients, nutrients and chlorophyll-a, and chlorophyll-a and fish assemblage descriptors across a range of oxbow lakes in the alluvial valley of the Mississippi River in Mississippi and Arkansas. Large-scale features considered in our study included land use and lake morphology; nutrients included phosphorus and nitrogen; and fish descriptors included various representations of biodiversity, taxonomic structure, and fish function. The specific objectives of this study were to estimate the relative importance of large-scale features on nutrients, the strength of the relationship between nutrients and primary productivity, and expose aspects of the fish assemblage best related to primary productivity. Because our focus was on nutrients, we did not consider potentially strong links such as depth and fish assemblage. We hypothesized that both landcover and lake morphometry would account for much of the variation in nutrients, that nutrients would be strongly related to algal density, and that algal density would be strongly related to various aspects of fish assemblages.
Methods
Study area
The alluvial valley of the Lower Mississippi River extends from near Cairo, Illinois south to the confluence of the Mississippi and Atchafalaya rivers in east-central Louisiana. Because of the rich alluvial soils, this region has been highly modified to develop and protect agricultural interests. The valley was once dominated by bottomland hardwood forests in the 19th century, but about 80 % of the forests were subsequently harvested and converted to agriculture (Forsythe 1985). The valley includes hundreds of oxbow lakes created by the meandering of prehistoric river systems, the Mississippi River, and various other smaller rivers that flow through the valley and join the Mississippi River (Baker et al. 1991). In addition to forest clearing, major modifications have been made to the valley’s hydrology through the construction of an extensive levee system associated with the Mississippi River and tributaries, and the construction of several large reservoirs that control the timing of water discharged into the valley from adjacent highlands. Oxbow lakes in the floodplain accumulated sediments and nutrients introduced over centuries by natural flooding, but accumulation rates have been exacerbated in the last century largely by runoff originating from surrounding agricultural lands (Ritchie et al. 1979; McHenry et al. 1982).
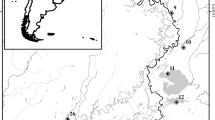
In all, 23 oxbow lakes selected from tributaries to the Mississippi River in Mississippi and Arkansas were investigated in 2007–2011 (Fig. 1). Lakes were selected based on accessibility and diverse representation of landcover and morphometry. In particular, efforts were made to select lakes along gradients of depth, surface area, and watershed composition. The features considered in lake selection have been identified as key driving forces for many processes in lentic systems (e.g., Magnuson et al. 1998; Tejerina-Garro et al. 1998).

Map of the alluvial valley region of Lower Mississippi River of Mississippi and Arkansas, with locations of 23 floodplain lakes included in our study. The inset identifies the location of the study region in the southeastern United States
Landcover
Land-use composition surrounding each lake was calculated using spatial analyst tools in the ArcGIS version 10 software (ESRI, Redlands, CA). Aerial images for lakes were obtained from websites of the Arkansas Geographic Information Office (AGIO; www.gis.state.ar.us, accessed 3 April 2012) and the Mississippi Automated Resource Information (MARIS; www.maris.state.ms.us, accessed 3 April 2012). Individual lake watersheds could not be defined due to the lack of sufficient topographic relief in the region (Baker et al. 1991). Instead, concentric bands (50 m for the riparian zone; 1,000 m for the watershed) were drawn around each lake. Percentages of land use in forests and agriculture available from the AGIO and MARIS databases were calculated within each band. Thus, two variables described the watershed and two variables described the riparian zone. Agriculture in this region consists primarily of row crops (corn Zea mays, soybeans Glycine max, cotton Gossypium hirsutum, and rice Oryza sativa) with limited conservation tillage. Forests consist primarily of bottomland hardwoods that encompass streams and wetlands as forests usually remain in areas too moist to farm.
Lake morphometry
We considered depth and size as key morphometry descriptors in oxbow lakes. Maximum depth was defined as the deepest point detected by depth soundings taken from a boat traveling in a zig-zag pattern along the former thalweg between the two ends of each lake. Depth soundings were made with a handheld depth finder (DF2200PX, NorCross Marine, Orlando, FL). Maximum depth was selected as opposed to mean depth because it was easier to measure and it better characterizes the cross-sectional morphology of abandoned river channels than mean depth, although these two depth descriptors are correlated. Lake size was defined with four descriptors including surface area, length, width, and length-to-width ratio. Area, length, and width were estimated with spatial analyst tools in ArcGIS using aerial images obtained from the AGIO and MARIS databases.
Nutrients and algal levels
Water samples were collected from the study lakes during July–August. This period was selected because it includes the warmest months of the year in the study region, with high primary production and limited rainfall. Collections were made between 1,100 and 1,330 h to ensure similar light conditions. Composite surface (<0.5 m) grab samples included ten 1-L subsamples collected over the deepest part of each lake. A 3-L water sample was taken from the composite, stored in polyethylene bottles, and preserved in ice for transport to a laboratory for nutrient analyses. Approximately 150 mL of the composite sample was filtered through a 0.45-μm glass-fiber filter paper to remove algae cells. The filter paper containing the residual was folded in half twice, wrapped in aluminum foil, placed within a sealed plastic bag and preserved in ice for transport to laboratory facilities. At the laboratory, the 3-L water sample and the filter were frozen at −20 °C until analysis.
Water samples were analyzed for total phosphorus and total nitrogen after micro-Kjehldahl digestion with hydrofluoric acid and hydrogen peroxide (APHA (American Public Health Association) 1998), on a FIA8500 Lachat (HACH, Loveland, CO). Detection ranges were 0.001–10 mg L−1 at 880 nm for phosphorus, and 0.005–10 mg L−1 at 530 nm for nitrogen, in a 50 mm flow cell.
Algal levels were indexed with chlorophyll-a concentrations estimated with spectrophotometric analysis. The glass-fiber filter paper used to collect algae in the field was soaked in 90 % buffered acetone overnight. The following morning samples were vortexed, filters were pressed to the bottom of the tube with a stainless steel spatula, and spun down in a centrifuge for 5 min to remove cellular debris. Chlorophyll-a concentration was measured on a HACH DR5000 using standard, tricolormetric methods (APHA (American Public Health Association) 1998).
Fish collections
Fish were collected during daytime hours by a boat electrofisher equipped with a GPP 7.5 Smith-Root™ pulsator unit (Smith-Root, Vancouver, WA). Pulsed DC electricity was cycled at 60 Hz with voltage output adjusted according to the specific conductance of each lake to maintain a constant output of 6–8 A. Individual samples consisted of 0.25 h of continuous electrofishing along indiscriminate shoreline areas, and 4–8 samples were collected and composited depending on lake area. Fish were netted from the bow of the boat by two netters equipped with 2.7-m (handle length) dip nets with 0.4-cm bar mesh. Fish were identified to species and counted before release near the site of capture. Those species too difficult to identify in the field were preserved in a 10 % formalin solution and transported to the laboratory for identification with taxonomic keys (Ross 2001).
Fish assemblage descriptors
We examined fish biodiversity, taxonomic guilds, and functional guilds. Ten biodiversity metrics descriptive of species richness, evenness, diversity, and dominance were estimated using diversity modules available in the PAST™ software (Hammer et al. 2001). Taxonomic guilds represent species characteristic of oxbow lake ecosystems in the Lower Mississippi River including sunfishes (i.e., Centrarchidae; 12 species), buffalos (i.e., Ictiobus; 3 species), gars (i.e., Lepisosteus; 3 species), and catfishes (i.e., Ictaluridae; 5 species). Functional guilds included spawning (4 classes; Balon 1990) and feeding (9 classes; Goldstein and Simon 1999) guilds. Guild structure was defined in terms of percentage composition of its components (i.e., species, classes) and estimated from counts given by the combined electrofishing samples.
Analyses
Relationships among sets of variables were examined using canonical analysis of principal coordinates (CAP; Anderson and Willis 2003). The CAP procedure is a multivariate data reduction technique that identifies axes running through a cloud of data points, represented by a similarity matrix, that have the strongest correlation with a set of external variables (Anderson and Robinson 2003). Because CAP analysis essentially ordinates one data matrix in consideration of another, it is a constrained analysis that uses an a priori hypothesis about variable dependency to construct correlations between sets of variables. The CAP approach to constrained ordination is similar to a canonical correlation analysis based on a matrix of explanatory variables. In our analyses, watershed (2 variables), riparian zone (2 variables), lake size (4 variables), and depth (1 variable) each separately represented a causal set of external variables analyzed against the nutrients similarity matrix (2 variables). Conversely, the nutrient variables were then made the causal external variables when analyzed against the chlorophyll-a (1 variable) similarity matrix. Similarly, chlorophyll-a was considered the causal external variable when analyzed against the biodiversity similarity matrix, the four taxonomic guild similarity matrices, and the two functional guild similarity matrices.
All resemblance matrices of species or functional composition were constructed with the Bray-Curtis similarity coefficient after transforming (loge x + 1) to reduce skewness. All other resemblance matrices were constructed with the Euclidean similarity coefficient after each variable was normalized to zero mean and one standard deviation. As needed, variables were transformed (loge x + 1) to reduce skewness before normalizing. The CAP analyses were performed using the PERMANOVA + add-on for the PRIMER software (Clarke and Gorley 2006; Anderson et al. 2008).
To further dissect the data, we examined scatter plots and Spearman correlations (rs) between the canonical coordinate scores produced by CAP, and the values of individual dependent variables. The objective of these analyses was to determine the direction of relationships. These analyses were conducted only if canonical correlations (R) among sets of variables estimated by the CAP procedure had a p < 0.20.
Results
Our efforts to select lakes along broad gradients of size, depth, and encircling landcover resulted in a dataset with a broad range of nutrient levels. Surface area ranged from 10 to 532 ha, maximum depth from 0.5 to 8.6 m, and riparian and watershed landcover from primarily agriculture to primarily bottomland hardwood forests (Table 1). Wetlands represented the next largest land cover class in the 50-m (median = 0 %, max = 38 %) and 1,000-m (median = 0.4 %, max = 23 %) bands, but were not considered in the analyses because in most lakes they represented only a small component. Total phosphorus ranged from 0.116 to 0.306 mg L−1, and total nitrogen from 0.634 mg to 2.580 mg L−1. Nutrients showed various strengths of correlations with lake physical descriptors (Fig. 2). There was a strong canonical correlation between nutrients and watershed land use (R = 0.68; p < 0.01), riparian zone land use (R = 0.59; p = 0.02), and between nutrients and maximum depth (R = 0.60; p = 0.05). There was no statistically significant correlation between nutrients and lake size (R = 0.42; p = 0.28).

Canonical correlations suggestive of the connectedness among land use, nutrients, algae, and fish assemblages in oxbow lakes of the alluvial valley of the Lower Mississippi River
Analysis of canonical coordinate scores revealed several multivariate relationships between nutrients and environmental features. For the watershed, as percentage of agriculture land use (min–max = 0−76) increased, and as forests (1–100) decreased, both total nitrogen and total phosphorus increased. The relationship between nutrients and land use was stronger for total nitrogen (rs = 0.91; p < 0.01) than for total phosphorus (rs = 0.48; p = 0.03) as suggested by their correlation with the canonical coordinate scores. For the riparian zone, as percentage of agriculture (0–51) increased, and as forests (0–100) decreased, both total nitrogen and total phosphorus increased. The relationship between nutrients and land use and their canonical coordinate scores were strong for total nitrogen (rs = 0.93; p < 0.01) and for total phosphorus (rs = 0.84; p < 0.01). As maximum depth increased, both total nitrogen and total phosphorus decreased, with the relationship being equally strong for total nitrogen (rs = −0.92; p < 0.01) and for total phosphorus (rs = −0.91; p < 0.01).
The study lakes displayed a broad range of chlorophyll-a values ranging from 5.8 to 139.6 μg L−1. Chlorophyll-a levels were strongly correlated with the set of nutrient variables (R = 0.71; p < 0.01). Analysis of canonical coordinate scores indicated direct relations between chlorophyll-a and total phosphorus (rs = 0.71; p < 0.01), and total nitrogen (rs = 0.69; p < 0.01).
Chlorophyll-a was correlated with some aspects of the fish assemblage. Chlorophyll-a showed a strong canonical correlation with sunfish composition (R = 0.66; p = 0.01), and a marginally significant canonical correlation with fish species biodiversity (R = 0.38; p = 0.08). There was no significant canonical correlation between chlorophyll-a and catfish, gar, or buffalo taxa guilds, or between chlorophyll-a and the spawning and feeding guilds.
Analysis of individual sunfish species identified inverse, direct, and no correlations with chlorophyll-a, depending on species. Spearman correlations between chlorophyll-a and spotted bass, largemouth bass, redspotted sunfish, longear sunfish, and redear sunfish percentage composition in the sunfish assemblages were −0.80, −0.68, −0.75, −0.68, and −0.62, respectively (all p < 0.01). Conversely, correlations for white crappies (primarily small adults <15 cm) and orangespotted sunfish were 0.74 (p < 0.01) and 0.52 (p = 0.03), respectively. Whereas bluegill, green sunfish, and black crappie showed neither a direct nor inverse correlation with chlorophyll-a, a scatter plot suggested possible arch-shaped relationship, with intermediate levels of chlorophyll-a supporting the highest representations of these species in the sunfish assemblage (Fig. 3). Additionally, scatterplots of species composition relative to chlorophyll-a were generally wedge-shaped, suggesting that chlorophyll-a may be a factor limiting species composition, but within a given level of chlorophyll-a, other unmeasured factors are also at work (Cade et al. 1999).

Scatterplots of selected sunfish species composition (percentage of sunfishes) relative to chlorophyll-a. Relationships were often wedge- or envelope-shaped, suggesting that chlorophyll-a may be a factor limiting composition, but within a given level of chlorophyll-a, other unmeasured factors are also at work
Analysis of canonical coordinate scores for biodiversity metrics indicated inverse relations between chlorophyll-a and species richness, diversity, and evenness, but direct relations with dominance metrics. Spearman correlations between the canonical coordinate scores and species richness metrics were −0.70, −0.69, and −0.54 for Margalef, Menhinick, and raw richness, respectively. For diversity metrics, correlations were −0.98, −0.96, and −0.74 for Shannon, Brillouin, and Fisher diversities, respectively. For evenness metrics, correlations were −0.81 and −0.63 for Pielou and Buzas-Gibson evenness, respectively. For dominance, correlations were 0.96 and 0.93 for Simpson and Berger-Parker dominance metrics, respectively. All correlations were statistically significant (p < 0.02).
Discussion
Our results generally support the connectedness paradigm in oxbow lakes of the Lower Mississippi River. Watershed and riparian zone land use, and lake depth but not size, were associated with nutrient concentrations. In turn, nutrients were associated with chlorophyll-a levels, and chlorophyll-a was associated with fish assemblages. Nevertheless, several aspects of the fish assemblages were marginally associated or not associated with chlorophyll-a.
Elongated bodies of water such as oxbow lakes tend to have strong ties to terrestrial landscapes because of long shorelines that allow more inputs of nutrients and inorganic and organic matter. There was a strong connectedness between land use and nutrients. In general, a shift in land use from forest to agriculture in the watershed was better associated with total nitrogen than with total phosphorus, but both nutrients were equally relevant in relation to the riparian zone. It is possible that the high correlations between lake nitrogen concentrations and agricultural land in the vicinity of lakes may be linked to recent agriculture shifts. The alluvial valley of the Lower Mississippi River in Mississippi and Arkansas is highly productive agricultural land, with large portions of the landscape in row crops including soybeans, cotton, and corn. Recently there has been a shift in crops driven by legislation. The 2007 Renewable Fuel Standard program requires that transportation fuel sold in the United States contains a minimum volume of renewable fuel. Because corn is used to make ethanol, this government program has resulted in a 47 % decrease in cotton area and a concomitant 288 % increase in corn area across Arkansas and Mississippi (Welch et al. 2010). This shift in agricultural use has the potential to increase nitrogen exports into lakes and waterways as corn in the region has a 200–250 kg ha−1 nitrogen fertilizer prescription, nearly double that of cotton. Thus, the strong relationship between agriculture land cover and nitrogen may be a function of agricultural practices and may shift depending on crop-specific amount and timing of fertilizer applications.
Lake size was not linked to nutrients, but lake depth was. The lack of a size effect may be a function of the morphology of oxbow lakes. Because oxbow lakes are elongated, the relative size of the transition between land and water is approximately constant across lake sizes. Thus, lake area increases linearly with perimeter, unlike bowl-shaped lakes where lake area increases exponentially with perimeter. Depth affects abiotic and biotic attributes at the local lake scale. It is a causal factor that drives many physical and chemical variables that organize nutrient storage and processing and contribute to shaping fish assemblages in floodplain lakes (Miranda 2011). Thus, correlations between fish and physical (e.g., transparency, temperature, habitat structure), chemical (e.g., pH, oxygen), and biological (e.g., autotrophs, heterotrophs) factors may ultimately be totally or partly regulated by depth. Depth of oxbow lakes is partially determined by lake age, but depth reductions have been intensified by deforestation (Wren et al. 2008) and may require manipulation and management (e.g., dredging, low dams, regulations on water withdrawals for irrigation). Depth affects the extent of sediment resuspension by wave action and by bottom feeders, nutrient releases from sediments, and illumination of the water column. Studies have suggested that shallower lakes have lower phosphorus retention and phosphorus content in the sediments (Nõges et al. 2007) and thus the availability of phosphorus in the water column tends to be higher. In shallow lakes where sediments are in permanent contact with the euphotic/trophogenic layer, the same portion of phosphorus may be recycled and brought back to the water column several times during a season (Nõges 2009) to support higher planktonic photosynthesis (Scheffer and van Nes 2007). In deep lakes, less of the bound phosphorus that settles out becomes available for photosynthetic organisms. Due to faster nutrient cycling and generally higher light availability, shallow lakes have, on average, higher productivity per unit of area than deep lakes at comparable total phosphorus and dissolved inorganic nutrient concentrations (Schindler and Scheuerell 2002; Mehner et al. 2005).
The relationship between nutrients and primary production has been the focus of many investigations. Phosphorus and nitrogen are considered limiting factors, but the importance of these two nutrients may vary between locations and even seasonally within a location. Although phosphorus is a limiting factor for most lakes and reservoirs, in some regions the nutrient paradigm centers on nitrogen rather than phosphorus (Canfield 1983; Pridmore et al. 1985). The reason for nitrogen limitation is not yet understood because of a long-held tenet in limnology that states that nitrogen fixation will compensate for shortfalls (Schindler 1977) and that nitrogen limitation is not a persistent condition. This belief does not seem as universal as once thought (Knowlton and Jones 1996). In some regions, nitrogen limitation may be a function of abundant phosphorus in the geological formation of the region (Canfield 1983). This may be the case in our study lakes. Because the alluvial soils and aquifers are naturally high in phosphorus, oxbow lakes in the Lower Mississippi River tend to have high phosphorus levels and may be nitrogen limited. Recent research in these lakes (J. Murdock, USDA Agricultural Research Service, National Sedimentation Laboratory, unpublished data) has shown that phytoplankton growth can be limited by nitrogen when total nitrogen concentrations were below approximately 2 mg L−1 (91 % of the lakes in our sample).
Chlorophyll-a was strongly correlated with sunfish species composition, marginally correlated with biodiversity, but not correlated with feeding and spawning guilds or various taxonomic guilds. The sunfish family is native to North America and likely originated in remnant-channel lakes in Southeastern floodplain ecosystems (Miller 1959; Branson and Moore 1962) where they are dominant. Thus, evolutionarily their ecology is tightly linked to oxbow lakes and their species composition respond noticeably to direct changes induced by algal biomass (e.g., water clarity) or to indirect changes (e.g., water quality, food resources). Fish species biodiversity was weakly linked with chlorophyll-a, which may be expected considering species association in these floodplain lakes can be greatly influenced by immigration and emigration during wet periods, and this effect varies depending on the degree of connectivity between the lake and adjacent rivers (Miyazono et al. 2010; Dembkowski and Miranda 2011). Taxa guilds such as gars, buffalos, and catfishes move in and out of oxbow lakes to spawn or feed, often to and from adjacent rivers or wetlands, and therefore may not adequately reflect the cascade of events initiated at the watershed level. We suspect the lack of correlation with spawning guilds is also related to the ability of fish to move in and out of most of these oxbow lakes. Moreover, we are not satisfied that the feeding groups classification we applied, or any other currently available, is sufficient to adequately document the complexity of food webs in oxbow lakes.
Our results suggest that because nutrients are merely a link in a chain of connected events, they can be determined directly, or gauged indirectly by measuring predictor or response variables. The choice of variables can be made based on sensitivity to change, cost of measurement and analysis, importance of the variable to a certain lake designated use, and access or availability of data. For example, in the Lower Mississippi River it has been estimated that there are nearly 1,500 oxbow lakes (Baker et al. 1991). Appraisal of this large number of lakes through onsite visits is virtually impractical, and many of them are inaccessible because they are surrounded by private lands. Nevertheless, predictions about their nutrient status and fish assemblages are possible through analyses of land-use data obtained from GIS sources. Estimates of lake depth are also possible through various imaging and non-imaging remote sensing techniques (Gao 2009), and in oxbow lakes of the Lower Mississippi River Landsat reflectance values was effectively correlated with empirical determinations of lake maximum depth made onsite (L.E. Miranda, unpublished data).
Our analysis estimated connections over a linear organization of system components. Nevertheless, interactions among components likely occur and may account for unexplained variability in our results. For example, we hypothesized that depth influenced availability of nutrients, which affected primary production, and eventually the fish assemblage. Nevertheless, depth may also influence the fish assemblage directly by providing vertical diversity in habitat. Similarly, while chlorophyll-a influences fish assemblages through trophic dynamics, it also contributes to the murkiness of the water and thus the cover that small species may need to escape predation. In the study lakes, orangespotted sunfish (a small species generally <7 cm total length) and white crappies (primarily small, possibly stunted, adults) were correlated with chlorophyll-a, we suspect partly due to the added shading provided by chlorophyll-a. Nevertheless, the shading effect of chlorophyll-a is difficult to distinguish from that of other suspended solids. The study lakes are often highly turbid due to sediment loading, and light does not penetrate far into the water column. However, phytoplankton appears to be adapted as they increase cellular chlorophyll under low light conditions and algae can be near maximum photosynthetic rates at low-light levels (J. Murdock, USDA Agricultural Research Service, National Sedimentation Laboratory, unpublished data), suggesting phytoplankton can flourish and provide additional shade in systems with high suspended sediments.
Ecologists have recognized the importance of hierarchical linkages between lotic ecosystems (Frissell et al. 1986; Townsend 1996; Allan and Johnson 1997). These concepts have mostly been applied as a means of identifying a scale at which to assess a set of dynamics, rather than to emphasize the connectedness among levels. Recognition that land use and lake depth influence events that cascade through oxbow lake ecosystems suggests a top-down hierarchy basis for developing management and conservation plans in lentic ecosystems. Resource managers can use surrounding land-use composition to broadly characterize lake states and identify where resources need to be allocated. Looking at surrounding land use may allow for efficient and effective early-decision making in efforts to restore and protect oxbow lakes. Land users in the region are starting to implement best management practices that are designed to retain sediment (Kröger et al. 2013), transform and reduce nutrient concentrations and loads (Kröger et al. 2012), and retain and capture surface runoff which would result in less water moving downstream (Kröger et al. 2008). Holistic landscape management coupled with depth preservation or reclamation can lead to improved water quality (i.e., a decrease in sediments and nutrients) running off agricultural landscapes, and ultimately lead to recovered in-lake conditions and fish community improvements.
References
Allan JD, Johnson LB (1997) Catchment-scale analysis of aquatic ecosystems. Freshw Biol 37:107–111
Anderson MJ, Robinson J (2003) Generalized discriminant analysis based on distances. Aust NZ J Stat 45:301–318
Anderson MJ, Willis TJ (2003) Canonical analysis of principal coordinates: a useful method of constrained ordination for ecology. Ecology 84:511–525
Anderson MJ, Gorley RN, Clarke KR (2008) PERMANOVA+ for PRIMER: guide to software and statistical methods. PRIMER-E, Plymouth, p 237
APHA (American Public Health Association) (1998) Standard methods for the examination of water and wastewater. 20th edn. APHA, Washington. p 1325
Baker JA, Killgore KJ, Kasul RL (1991) Aquatic habitats and fish communities in the lower Mississippi River. Rev Aquat Sci 3:313–356
Balon EK (1990) Epigenesis of an epigeneticist: the development of some alternative concepts on the early ontogeny and evolution of fishes. Guelph Ichthyol Rev 1:1–42
Branson BA, Moore GA (1962) The lateralis components of the acoustico-lateralis system in the sunfish family Centrarchidae. Copeia 1962:1–108
Cade BS, Terrell JW, Schroeder RL (1999) Estimating effects of limiting factors with regression quantiles. Ecology 80:311–323
Canfield DE Jr (1983) Prediction of chlorophyll a concentrations in Florida lakes: the importance of phosphorus and nitrogen. J Am Water Resour Assoc 19:255–262
Clarke KR, Gorley RN (2006) PRIMER V6: user manual/tutorial. PRIMER-E, Plymouth, p 192
Dembkowski DJ, Miranda LE (2011) Comparison of fish assemblages in two disjoined segments of an oxbow lake in relation to connectivity. Trans Am Fish Soc 140:1060–1069
Elser JJ, Marzolf ER, Goldman CR (1990) Phosphorus and nitrogen limitation of phytoplankton growth in the freshwaters of North America: a review and critique of experimental enrichments. Can J Fish Aquat Sci 47:1448–1477
Forsythe SW (1985) The protection of bottomland hardwood wetlands of the Lower Mississippi Valley. Trans N Am Wildl Nat Resour Conf 50:566–572
Frissell CA, Liss WJ, Warren CE, Hurley MD (1986) A hierarchical framework for stream habitat classification: viewing streams in a watershed context. Environ Manag 10:199–214
Frost PC, Kinsman LE, Johnston CA, Larson JH (2009) Watershed discharge modulates relationships between landscape components and nutrient ratios in stream seston. Ecology 90:1631–1640
Gao J (2009) Bathymetric mapping by means of remote sensing: methods, accuracy and limitations. Prog Phys Geogr 33:103–116
Goldstein RM, Simon TP (1999) Toward a united definition of guild structure for feeding ecology of North American freshwater fishes. In: Simon TP (ed) Assessing the sustainability and biological integrity of water resources using fish communities. CRC Press, Boca Raton, pp 123–202
Hammer Ø, Harper DAT, Ryan PD (2001) PAST: Paleontological statistics software package for education and data analysis. Palaeontologia Electronica 4
Jones JR, Hoyer MV (1982) Sportfish harvest predicted by summer chlorophyll-α concentration in Midwestern lakes and reservoirs. Trans Am Fish Soc 111:176–179
Jørgensen SE, Xu FL, Costanza R (eds) (2005) Handbook of ecological indicators for assessment of ecosystem health. CRC Press, Boca Raton, p 448
Knowlton MF, Jones JR (1996) Nutrient addition experiments in a nitrogen-limited high plains reservoir where nitrogen-fixing algae seldom bloom. J Freshw Ecol 11:123–130
Kröger R, Cooper CM, Moore MT (2008) A preliminary hydrological investigation into an innovative controlled drainage strategy in surface drainage ditches: low-grade weirs. Agric Water Manag 95:678–684
Kröger R, Pierce SC, Littlejohn KA, Moore MT, Farris JL (2012) Decreasing nitrate-N loads to coastal ecosystems with innovative drainage management strategies in agricultural landscapes: an experimental approach. Agric Water Manag 103:162–166
Kröger R, Littlejohn KA, Pierce SC, Henderson J, Brandt J, Flora C, Poganski BH, Prevost JD (2013) Evidence towards sediment accumulation characteristics of slotted pipes as best management practices on agricultural landscapes. Ecol Eng 51:249–255
Magnuson JJ, Tonn WM, Banerjee A, Toivonen J, Sanchez O, Rask M (1998) Isolation vs. extinction in the assembly of fishes in small northern lakes. Ecology 79:2941–2956
McHenry JR, Cooper CM, Ritchie JC (1982) Sedimentation in Wolf Lake, lower Yazoo River basin, Mississippi. J Freshw Ecol 1:547–558
Mehner T, Diekmann M, Brämick U, Lemcke R (2005) Composition of fish communities in German lakes as related to lake morphology, trophic state, shore structure and human-use intensity. Freshw Biol 50:70–85
Miller RR (1959) Origin and affinities of the freshwater fish fauna of western North America. In: Hubbs CL (ed) Zoogeography. American Association for the Advancement of Science Publication 51. Arno Press, New York, pp 187–222
Miranda LE (2011) Depth as an organizer of fish assemblages in floodplain lakes. Aquat Sci 73:211–221
Miyazono S, Aycock JN, Miranda LE, Tietjen TE (2010) Assemblage patterns of fish functional groups relative to habitat connectivity and conditions in floodplain lakes. Ecol Freshw Fish 19:578–585
Niemi GJ, DeVore P, Detenbeck N, Taylor D, Lima A, Pastor J, Yount JD, Naiman RJ (1990) Overview of case studies on recovery of aquatic systems from disturbance. Environ Manag 14:571–587
Nõges T (2009) Relationships between morphometry, geographic location and water quality parameters of European lakes. Hydrobiologia 633:33–43
Nõges T, Järvet A, Kisand A, Laugaste R, Loigu E, Skakalski B, Nõges P (2007) Reaction of large and shallow lakes Peipsi and Vortsjaerv to the changes of nutrient loading. Hydrobiologia 584:253–264
Pridmore RD, Vant WN, Rutherford JC (1985) Chlorophyll-nutrient relationships in North Island lakes (New Zealand). Hydrobiologia 121:181–189
Rader RB, Richardson CJ (1992) The effects of nutrient enrichment on algae and macroinvertebrates in the Everglades: a review. Wetlands 12:121–135
Ritchie JC, Cooper CM, McHenry JR (1979) Recent accumulation of sediment in lakes in the Bear Creek watershed in the Mississippi Delta. Southeast Geol 20:173–180
Ross ST (2001) The inland fishes of Mississippi. University of Mississippi Press, Jackson, p 624
Scheffer M, van Nes EH (2007) Shallow lakes theory revisited: various alternative regimes driven by climate, nutrients, depth and lake size. Hydrobiologia 584:455–466
Schindler DW (1977) Evolution of phosphorus limitation in lakes. Science 195:260–262
Schindler DE, Scheuerell MD (2002) Habitat coupling in lake ecosystems. Oikos 98:177–189
Tejerina-Garro FL, Fortin R, Rodríguez MA (1998) Fish community structure in relation to environmental variation in floodplain lakes of the Araguaia River, Amazon Basin. Environ Biol Fishes 51:399–410
Townsend CR (1996) Concepts in river ecology: pattern and process in the catchment hierarchy. Arch Hydrobiol Suppl 113:3–21
Vadeboncoeur Y, Lodge DM, Carpenter SR (2001) Whole lake fertilization effects on distribution of primary production between benthic and pelagic habitats. Ecology 82:1065–1077
Ward JV (1998) Riverine landscapes: biodiversity patterns, disturbance regimes, and aquatic conservation. Biol Conserv 83:269–278
Welch HL, Green CT, Rebich RA, Barlow JRB, Hicks MB (2010) Unintended consequences of biofuels production: The effects of large-scale crop conversion on water quality and quantity. U.S. Department of the Interior, U. S. Geological Survey, Open-File Report 2010–1229
Wetzel RG (2001) Limnology: lake and river ecosystems, 3rd edn. Academic Press, San Diego, p 1006
Wiens JA (2002) Riverine landscapes: taking landscape ecology into the water. Freshw Biol 47:501–515
Wren DG, Davidson GR, Walker WG, Galicki SJ (2008) The evolution of an oxbow lake in the Mississippi alluvial floodplain. J Soil Water Conserv 63:129–135
Acknowledgments
Funding and support was provided by the Vicksburg District of the U.S. Army Corps of Engineers through K. J. Killgore, Mississippi State University, and the U.S. Geological Survey. We thank T. Alfermann, N. Aycock, D. Dembkowski, D. Goetz, and S. Miyazono for assistance with field work; L. Brooks for water quality analyses; and J. Murdock for a helpful review. The use of trade names or products does not constitute endorsement by the federal government.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Miranda, L.E., Andrews, C.S. & Kröger, R. Connectedness of land use, nutrients, primary production, and fish assemblages in oxbow lakes. Aquat Sci 76, 41–50 (2014). https://doi.org/10.1007/s00027-013-0310-y
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00027-013-0310-y