
Abstract
Water stress in Mediterranean countries is the result of both variable and changing climatic conditions and widespread anthropogenic pressures. Evrotas, an intermittent river located in Southern Greece, was used as a case study to assess the impacts of water stress on Mediterranean lotic ecosystems. Based on hydrological analyses, it was revealed that during prolonged drought years, such as the summers of 2007 and 2008, the vast majority of the Evrotas riverbed was completely desiccated, primarily as a result of substantial water abstraction for irrigation. The effects of desiccation on the riverine ecosystem were evaluated using fish and macroinvertebrate assemblages according to the demands of the Water Framework Directive 2000/60/EC (WFD). Faunal responses to water stress were assessed through comparisons of assemblages attributes in perennial and intermittent reaches and pre-drought versus post-drought communities. Effects of hydrological disturbance on fish species richness, density, percentage composition and size structure were more pronounced in intermittent than in perennial sites. The most obvious and immediate impact was the elimination of populations in the intermittent reaches. However, upon flow resumption, the recolonisation from upstream perennial reaches began, thereby permitting partial re-establishment of the depleted fish communities. Nevertheless, the structural integrity of fish communities remained severely impacted and recovery was markedly slow. On the contrary, post-drought macroinvertebrate assemblages were not affected by summer droughts, and the recruitment processes were rapid after flow resumption. Our findings point to the necessity of establishing a distinction between naturally and artificially driven intermittent rivers. We, therefore, propose the introduction of an “artificially intermittent Mediterranean river” condition within the context of the WFD assessment applications.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Intermittent rivers and streams dominate surface runoff in large parts of Mediterranean Europe, especially in regions with semi-arid climatic conditions (Estrela et al. 1996). Intermittent lotic systems are defined as systems that flow only during times when they receive water from springs and surface runoff, while during dry periods they may cease to flow entirely or are reduced to stream pools (Gordon et al. 2004). Seasonal desiccation in lotic systems can be attributed either to natural climate variability (meteorological droughts) or to direct (water resources exploitation) and indirect (climate change) anthropogenic pressures, or a combination of both.
Meteorological droughts are periods of lower than average precipitation and relative humidity, greater solar radiation and higher temperatures of varying severity, duration and scale. In Europe, air temperature has increased by 1.2°C during the 20th century, while precipitation has diminished to 20% in some of its southern regions, particularly in the Eastern Mediterranean (Norrant and Douguédroit 2006; EEA 2008), where the frequency duration and severity of droughts have increased over the past decades (World Bank 2003).
Historically, water use and management practises in the Mediterranean region were well adapted to the natural water availability. During the last decades, however, increased water demand for agricultural, industrial and urban development has created widespread pressures on natural water resources (Bradford 2000). Agriculture is by far the largest consumer of freshwater resources in Southern Europe. Irrigated agricultural land now covers extensive parts of the Mediterranean countries (EEA 2000) and official data on water quantities allocated for irrigation are immense compared to the EU average (Eurostat 2009; EEA 2009).
The effects of direct and indirect anthropogenic pressures on hydrological functions have created ‘water stress’ conditions that have been considerably affecting natural flow regimes during the past half century (Smith 1997; UNEP/MAP 2003; Ludwig et al. 2009). Water stress causes deterioration of freshwater resources in terms of quantity (aquifer over-exploitation, dry rivers, etc.) and quality (organic pollution, eutrophication, saline intrusion, etc.) (UNEP 2004). As it concerns quantity, an overall runoff decrease of at least 20% in Mediterranean rivers within the past half century has been estimated by Ludwig et al. (2009), while Balkan rivers reveal even higher reduction trends (Skoulikidis 2009). An increase in the frequency, duration and intensity of low flows over the past decades has also been reported (Demuth 2001; Skliris et al. 2007). In many cases, natural flow, for at least parts of the year, has vanished (Smakthin 2003), primarily due to water abstraction, as documented for Iberian rivers (Benejam et al. 2010). Evidence from Greece (Chadzichristidi et al. 1991; Economou et al. 1999a; Bobori and Economidis 2006) also indicates a change of former perennial rivers towards intermittent ones during drought years. The alteration of flow regimes is considered to be one of the most serious and recurrent threats to the ecological integrity of natural river ecosystems (Bunn and Arthington 2002; Bond et al. 2008). Numerous studies have indicated that drought episodes can significantly alter the fish community structure and dynamics, through influencing fish growth, survival and reproductive activities, and thereby affecting the species composition, abundance, and age structure of local fish assemblages (Tramer 1977; Boulton and Lake 1992; Mathews 1998; Matthews and Matthews 2003; Lake 2000, 2003; Magoulick and Kobza 2003). Similarly, droughts can have profound effects on macroinvertebrate assemblages, typically by decreasing species richness (Gore 1982; Boulton and Lake 1992; Otermin et al. 2002).
Although rivers in arid and semi-arid climates are extremely sensitive to hydrological and various anthropogenic pressures, they have been poorly studied. In fact, they constitute one of the least known types of fluvial ecosystems (Uys and O’Keeffe 1997; Jacobson et al. 2004; Ryder and Boulton 2005; TempQsim Consortium 2006). Even the EU’s Water Framework Directive (WFD) does not specifically address intermittent rivers. Recently, in view of increased human demands, potential climate change impacts, and ecosystem conservation requirements, temporary running waters have attracted increasingly high economic and ecological interest (Larned et al. 2010). Therefore, it is necessary to include intermittent rivers and intermittent reaches of larger river basins in the ecosystem-based integrated water resources management scheme, as dictated by the WFD. To develop effective River Basin Management Plans for intermittent rivers, it is essential to enhance scientific knowledge on their structural and functional characteristics. This holds particularly true for intermittent rivers which have been largely created by human-induced water stress.
The scope of this article is to explore the ecological effects of human-induced hydrological regime alterations to former perennial stream ecosystems using the Evrotas River (Greece) as a case study. Our approach involves a hydrological analysis and a study of the impacts of water abstractions during particularly dry years on hydrology and aquatic biota, utilising fish and macroinvertebrates. We specifically attempted to: (a) estimate the contribution of human-induced water stress on the river’s hydrological conditions and flow pattern, and (b) assess the impacts of water stress on prominent biotic attributes of the ecosystem.
The following particular objectives form the focus of the investigation:
-
a.
establish the extent and severity (and the spatial and temporal dimensions) of summer droughts,
-
b.
discern the relative contribution of natural and anthropogenic causes to water stress,
-
c.
assess the magnitude of impacts of drought on biological communities in intermittent reaches through comparisons with perennial reaches and pre-drought vs post-drought assemblages,
-
d.
evaluate recolonisation processes of fish and macroinvertebrates in drought-affected areas, and
-
e.
for fish fauna, describe critical features and components of the recovery process (degree and velocity of re-establishment of fish communities in terms of richness, abundance, species composition and age structure).
Materials and methods
Study area
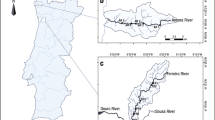
The Evrotas River Basin belongs to a mid-altitude Mediterranean basin, located in the south-eastern Peloponnese (Prefecture of Laconia) (Fig. 1). The river drains a total area of 2,418 km2 and discharges into the Laconic Gulf after crossing 90 km of valley basin. The basin expands between the mountain ranges of Taygetos (2,407 m a.s.l.) and Parnon (1,940 m a.s.l.), where numerous intermittent and ephemeral streams, discharge into the main course. The mountainous area of the basin is formed by Mesozoic-Palaeogene limestones (42% of the basin) and impermeable rocks, such as flysch and schists (29% of the basin), while the lower parts of the valley are filled with Pliocene and Quaternary sediments, which form extensive alluvial aquifers. The Evrotas basin has a typical Mediterranean climate with mild and cool winters and prolonged hot and dry summers, with an average annual temperature of 16°C and a mean annual precipitation of 803 mm (2000–2008) (Nikolaidis et al. 2009). The majority of rainfall occurs during the months of October through March; highest rainfall being in December and lowest in June. Besides surface runoff resulting from rainfall and snow melt, the river system is fed by numerous karstic and alluvial springs.

Geographical and sampling sites in the Evrotas River basin: Macroinvertebrates (encircled bullet), Fish (square), water level automatic gauging stations (star)
Most of the river basin’s landscape is covered by semi-natural areas accounting for 61% of the total river basin, followed by agricultural areas that cover 38%, while urban areas account for approximately 1%. The dominant anthropogenic pressures in the Evrotas River Basin are derived mainly from agricultural activities and include overexploitation of water resources for irrigation, disposal of agro-industrial wastes and agrochemical pollution. There are many, mostly illegal, surface water abstraction points (permanent and temporal weirs) throughout the length of the Evrotas and numerous private and municipal bore-hole drillings (estimated to about 3,500) for irrigational uses scattered over the entire basin. Only one wastewater treatment plant exists in the basin (city of Sparta), while villages are served by private permeable and impermeable cesspools.
The river hosts five native and two alien freshwater fish species. Three of the native species are range-restricted endemic cyprinids of outstanding conservation interest: Squalius keadicus is confined exclusively to this river; Pelasgus laconicus is found only in this basin and parts of the adjacent Alfeios River; while Tropidophoxinellus spartiaticus, occurs in only a few other rivers of the southern Peloponnese (Economou et al. 2007a). Within a wider biogeographic scope, the Evrotas River is considered a biodiversity hotspot (Skoulikidis 2008a) and one of the most biotically distinctive basins of the Ionian ecoregion due to its prolonged isolation from other river systems (Economou et al. 2007a; Zogaris et al. 2009).
Methodological design and data collection
The “undisturbed” hydrological regime of the Evrotas Basin was researched using the Leitbild methodology, which employs a multidisciplinary approach including historical survey and data analysis to construct an image of the river corridor prior to intense human interventions (Bostelmann et al. 1998). This included the following steps: (a) Numerous field surveys in order to investigate particular anthropogenic pressures related to water management throughout the basin (e.g., water abstraction sites, river engineering works etc.); (b) Information on all water uses and land-use changes was collected during field surveys as well as from personal communications with local authorities; (c) Historical literature and maps were surveyed and local citizens were interviewed, particularly the elderly; (d) Past and present hydrological data such as irrigated land area, monthly rainfall, air temperature (data beginning from the 1960s) and water discharge time series (particularly from Vrodamas gauging station for the period 1974–2008), and, finally; (e) The current water balance model of the basin was compared to a theoretical one, which referred to a time period prior to modern agricultural intensification.
For the study of hydrological processes, three automatic water level recording stations were installed and operated during the period from November 2006 to December 2007 at Vivari, at Sparta-bridge and at Oinus-tributary near its confluence with Evrotas (Fig. 1). Daily discharge was calculated from water level data and wet cross-section measurements. To illustrate the extent of drought and to designate the seasonal flow pattern, the hydrological attributes (e.g., surface-water flow and permanence) of the Evrotas River network was surveyed and mapped though extensive site inspections during wet and dry periods in 2007 and 2008.
The annual water balance of the basin for the period 1998–2000 was estimated using Ward’s (1975) classic mass balance equation. This particular period was selected because of data availability, while at the same time the rainfall height recorded in this period approximates the average value of the last decade. The potential evapo-transpiration was approximated according to Hamon’s method (Hamon 1961) and the real evapo-transpiration according to Hillel (1980). For the annual domestic water supply needs, the OECD (2000) guidelines were used (250 L/inh./day). Finally, the irrigation water uses were calculated from the area and the water needs of each cultivation type (Papazafeiriou 1999). The monthly water balance in the study basin was estimated using the Hillel method (1980, soil moisture deficiency) by employing the ISBH water balance software (http://www.intersoft.tk). The temporal distribution for the irrigation water abstractions was based on experience from other agricultural Greek catchments (Zacharias et al. 2003), while 2/3 of the olive trees were usually irrigated according to the Direction of Land Reclamation, Laconia Prefecture. Finally, the cumulative water abstractions and storages were estimated from the monthly water budget. The estimated surface runoff for the period 1998–2000 was compared with respective flow measurements near the outflow of the catchment to validate the results. Moreover, a hypothetical water management scenario was developed to identify the impacts of the intensive olive tree irrigation on Evrotas discharge, assuming that prior to the construction of the existing irrigation infrastructure, the agricultural abstractions were 30% of the today values (pers. Commun. Laconia Prefecture). For this purpose, the same water balance model was used, with the same meteorological and calibration values, in order to avoid inconsistencies and variations in the potential modeling errors that would impact the hydrologic scenarios comparison.
The impacts of desiccation on the biota were studied using fish and macroinvertebrate fauna. Differences in ecology, population dynamics and distribution patterns between fish and macroinvertebrates required different spatial sets of data as well as different sampling strategies and analytical procedures to be applied for drought effect assessments. Fish are entirely water-dependent organisms, and because frequent droughts over the last decades have resulted in the elimination of fish from the majority of the Evrotas’ tributaries (see “Results”), the field surveys were confined to the main river stem that still retains fish. On the contrary, macroinvertebrate species that have higher levels of resilience and resistance to hydrological disturbances are well represented in both the main river stem and in tributaries. However, macroinvertebrates are particularly sensitive to organic pollution, and in order to avoid confounding interpretations of community structure changes arising from both flow reduction and pollution, the survey design included macroinvertebrate sampling only in upstream tributary reaches where anthropogenic pollution is absent or restricted.
An assessment of potential pressures was carried out in the field so as to ensure that the selected upstream sites of macroinvertebrate sampling were not affected by other pressures except of summer river-bed desiccation. Physicochemical and hydrochemical assessments were performed along with macroinvertebrate sampling at Evrotas tributaries and along the main course of Evrotas River where fish sampling was conducted. In addition, physicochemical measurements were carried out along with fish samples. Water temperature, pH, conductivity and dissolved oxygen were measured in situ, whereas major ions (Ca2+, Mg2+, Na+, K+, HCO3 − , CO3 2−, Cl− and SO4 2−), silicate and nutrients (nitrate, nitrite, ammonia, total nitrogen, phosphate and total phosphorous) were analysed in the laboratory (for detailed description of analytical and quality control procedures see Skoulikidis et al. 2006, Skoulikidis 2008a). For the evaluation and classification of the physicochemical status of the sites, the Nutrient Classification System (Skoulikidis et al. 2006) was used.
Fish surveys were intended to examine the temporal sequence of events leading to the recovery of aquatic community integrity following catastrophic mortality during the 2007 severe drought. We used data from 20 sites located in four river reaches. Two reaches (R1, 6.6 km, and R3, 4.7 km) were spring-fed and represented perennial flow characteristics (i.e., never dried-out during the study period); and two reaches (R2, 14.6 km, and R4, 9.2 km) represented intermittent flow characteristics (i.e., dried-out completely in summer 2007) (see Fig. 1). The two intermittent reaches also dried-out during summer 2008, almost to a similar spatial scale as in summer 2007; however, the drying process was not as severe, and some pools persisted until the beginning of the rainy season. The four reaches were sampled in four consecutive periods: spring and summer 2007 and again spring and summer 2008. Densities were estimated as areal densities, i.e., numbers of fish caught in each sampling site divided by the area sampled. For some treatments, density and species composition data from all sites located in each reach were combined to provide an average for the reach. Differences between species areal density for intermittent and perennial sites were analysed, after log transforming the data to improve homoscedasticity and linearity, with a two-way ANOVA which was applied with SPSS 14.0. Proportion of young-of-the-year (YOY) of S. keadicus and T. spartiaticus was calculated based on the growth and reproduction patterns of the two species; all fish below 5 cm were assigned to age 0+, that is YOY. This distinction between YOY and older fish was not possible for P. laconicus, which is a small-bodied minnow (rarely exceeding 7 cm in length) with a prolonged spawning period.
Other fish species (e.g., Rainbow Trout and Eel) having sporadic occurrence and occurring at densities below 3% in the total catch were excluded from any analysis. Sampling was conducted using the CEN (European Committee for Standardisation) (1994) electrofishing sampling standards and a modified version of the FAME (2005) project protocols (Economou et al. 2007b). The sampling site selection criteria (e.g., landscape and macro-habitat representativeness), details of the sampling procedures and the recorded parameters (hydromorphological and physicochemical measurements, pressure variables) are provided by Skoulikidis (2008a). A Hans-Grassl GmbH battery-powered backpack electrofisher (Model IG200-2, DC pulsed, 1.5 KW output power, 35–100 Hz, max. 850) was routinely used as the sampling instrument. The fish caught were identified to species, counted, grouped in 5 cm class intervals, and were normally released back to the system to avoid population depletion. Subsamples were kept from some sites for associated biological studies.
To assess the effects of drought on the macroinvertebrate fauna, sampling was performed in five perennial and six intermittent tributaries (Fig. 1) that share common typological characteristics (RM4 typology as defined within the Intercalibration Exercise, EC 2007). Macroinvertebrates were collected on three occasions with the STAR-AQEM methodology (AQEM Consortium 2002) in May and September 2006, and in March 2007. The methodology includes a recording protocol of 130 variables that aim to give an impression of river and floodplain morphology, hydrology, hydrochemistry and vegetation composition at a local scale (site description data, etc.) and broader-scale variables (land cover at reach and catchment level, geology, etc.). The STAR-AQEM methodology is based on a multi-habitat scheme, designed for sampling major habitats proportionally according to their presence within a sampling reach. Each sample consists of 20 “replicates” taken from all microhabitat types at the sampling site with a share of at least 5% coverage, which must be distributed according to the share of microhabitats. Benthic macroinvertebrates were collected using a rectangular hand net of 25 cm × 25 cm with a mesh size of 500 μm nytex screen. Each of the 20 “replicates” was taken by positioning the net and disturbing the substrate in an area that equals the square of the frame width upstream of the net (0.25 × 0.25 m). Thus, a total of 1.25 m2 (0.25 × 0.25 × 20 replicates) was sampled for each sampling site. Benthic samples were preserved with ethanol concentration of ca 70%; specimens were collected with soft tweezers from the samples and identified to family level.
To compare macroinvertebrate community assemblages of intermittent streams prior to and after desiccation events, a cluster analysis on pre-drought (spring 2006) and post-drought (winter 2007) data was applied for intermittent stream reaches. To compare fauna assemblages in perennial and intermittent streams, a cluster analysis and an Analysis of Similarities (ANOSIM) was used. In addition, the following macroinvertebrate attributes were applied as metrics to assess community structure differences of the two hydrological stream types: Number of families, EPT (Ephemeroptera, Plecoptera, Trichoptera) and COH (Coleoptera, Odonata, Heteroptera). These metrics have been validated as robust indicators of seasonal changes in macroinvertebrate communities in Mediterranean rivers (Bonada et al. 2006). Cluster analysis and ANOSIM were conducted through the use of the PRIMER statistical programme (Clarke and Gorley 2006), while metrics EPT and COH were compared with box plot analysis using the STATISTICA software. Finally, the ratio EPT/(EPT + COH) which varies from 0 to 1 was used to compare differences among sites. To test whether there are differences of the EPT/(EPT + COH) ratio between perennial and intermittent sites, the t test was applied using the STATISTICA software.
Results
History of interventions and agricultural practises
Up until the early 20th century, most of the agriculture in the Evrotas valley was not irrigated and thus maintained a subsistent, low-intensity rural economy without extracting substantial surface and subsurface water resources (Cavanagh et al. 2002; Wagstaff 2002). Land reclamation works helped expand the production and variety of irrigated cultivations, particularly the promotion of citrus plantations after World War II. However, up to about the mid-20th century the greatest part of the river’s network maintained near-natural hydromorphological features and sustained surface water throughout the year, which is also indicated by the former existence of fish in many of the Evrota’s tributaries (Vagiakakos 1989; Economou et al. 1999b). During the period between the 1950s and 1990s, several large-scale irrigation, river engineering and drainage projects were carried out in order to reduce floods, control malaria and increase irrigated agricultural land. Over the last 3 decades, major hydromorphological alterations have occurred; irrigated cultivations have expanded towards natural and semi-natural land including riparian areas, while natural vegetation along the riparian zones has been dramatically altered and, more recently, the irrigation network expanded to olive groves (Bizas 1995; Dimopoulos et al. 2009). This resulted in widespread surface and ground water overexploitation, especially during dry years, which, in turn, affected the basin’s hydrological balance and resulted in the lowering of groundwater tables (Skoulikidis 2008b). During extreme drought periods, such as 1989–1993, increased water needs for irrigation resulted to the depletion of groundwater aquifers and, consequently, to the desiccation of extensive parts of the river network.
The extent and the underlying causes of desiccation during a drought period
During an average hydrological period (1998–2000), the ratio between mean annual precipitation and potential evapo-transpiration in the Evrotas basin was 0.46, indicating the semi-arid character of the basin (semi-arid zones: 0.20 ≤ P/PET < 0.5; UNESCO 1979). Intense droughts occur roughly every 7–8 years (Fig. 2). The years 2006 and 2007 were particularly dry (−18% precipitation compared to the multi-year average for both years). In 2007, the snow cover was unusually sparse and three heat waves affected the area during the summer, which went on to trigger catastrophic wild fires. In contrast, the precipitation in 2008 and 2009 approached the multi-year average.

Thirty-five year rainfall pattern (average of three gauging stations) and discharge variation at Vrodamas and the rainfall-discharge ratio (data: Laconia Prefecture)
Figure 3 illustrates the surface water of the Evrotas River during the wet period (April) and at the end of the dry period (October) in the drought year of 2007. During April, there was flow in most of the river network. By the end of September to mid October, water had vanished in almost all tributaries and most segments along the main river course had completely desiccated. Only some spring-fed riverine reaches (about 20% of the main river’s length), and a small number of headwater tributaries retained surface water. The hydrological condition of the river in summer/autumn 2008 was quite similar to that of summer/autumn 2007. During the desiccation process, in the summers of 2006–2008, isolated pools of various sizes were formed in several riverine and tributary sections. Some of the largest pools persisted throughout the summer, during the dry season, providing shelter for fish and other aquatic organisms. However, pool depth and surface area decreased constantly, resulting in severe crowding of fish with elevated mortality.

Hydrological maps of the Evrotas River Basin in spring (left) and at the end of summer (right) of 2007. In bold are the reaches with surface water. Light lines represent the ideal hydrologic network of the main river course
Precipitation and runoff data for the period 1974–2008 reveal a decreasing trend (Fig. 2). The decrease of discharge is the highest among 15 major Balkan rivers (Skoulikidis 2009). A comparison between the average rainfall of the initial 14 years of the time-series (1974–1988) for three gauging stations (Vasaras, Sellasia and Kastori) and the average rainfall of the last 14 years of the time-series (1994–2008) for the same stations showed a rainfall decline of 10.5% within the last time-period (average rainfall of initial time-period: 1,010 mm—average rainfall of last time-period: 904 mm). A similar comparison for the discharge at Vrodamas station (Fig. 1) showed a 51.2% decline (average discharge of initial time-period: 115 mm – average discharge of last time-period: 56.1 mm). The specific time-periods were selected in order to eliminate the effect of an interfering exceptional prolonged drought period (1989–1993) that affected most of Southern Europe (Skoulikidis 2009). Moreover, the value for the rainfall elasticity of stream flow has been calculated for the period 1974–2008 (Fig. 2). This provides an estimate of the sensitivity of long-term surface discharge to changes in long-term rainfall and contributes to the assessment of climate change on the water resources. The estimated rainfall elasticity to stream flow was very low compared to other catchments elsewhere in the world. In fact, Australian data have shown that a potential 1% change in rainfall leads to a 2–3.5% change in runoff, whereas in the Evrotas basin the respective value was very low (0.54) indicating a relatively weak relationship between rainfall and runoff in the particular area. (Chiew 2006).
Data from the three automatic water level gauging stations indicated that during the summer of 2007, the Evrotas at Sparta-bridge dried-out completely for over a month (end of August–mid October), whereas the downstream reach of Oinus ceased to flow for about 4 months (mid June–mid October). Based on the data from the gauging stations and field observations, flow velocity and water level declined to zero abruptly.
To quantify the effects of agricultural water exploitation on the Evrotas flow, the water balance of the river basin was estimated, based on a soil deficit mass balance equation (Ward 1975) according to: (a) the current water uses, and (b) the current water uses excluding the water used for irrigation of olive groves. The decision to adopt this approach was based largely on the fact that irrigation of olive trees has been only recently introduced in the basin. According to the methods applied, the mean annual precipitation, the potential evapo-transpiration and the real evapo-transpiration were estimated at 727, 1,586 and 5,66 mm, respectively. Finally, 162 mm represented surface runoff and infiltration. Domestic consumption (0.0091 km3) represented only 2% of the annual (surface and ground) water storages, while irrigation reached 73% (0.287 km3) and the remaining 24% covers the environmental demands (approx. 0.095 km3 annually). The amount of remaining water, at the end of the hydrologic period, after subtracting the current water abstractions for anthropogenic purposes was similar to the measured annual surface runoff in the downstream part of the catchment, which verifies the results of the water balance model (current hydrologic scenario).
The results of the monthly water budget analysis indicated that at the end of the hydrologic period the remaining water resources in the catchment was less than 38 mm (2.9 m3/s). Olive groves cover 77% of the Evrotas basin agricultural land. The total irrigation needs prior to the irrigation of olive groves was estimated to be 0.084 km3/year; i.e., 30% of the current uses. Accordingly, the discharge at the river outflow during the dry period was estimated at 122 mm (9.4 m3/s), which is over three times higher than the current one. Figure 4 presents the monthly variation of discharge during the irrigation period at Vrodamas gauging station (which dries out during severe drought years, e.g., in 1990 and 2007): (a) according to the current water uses and (b) after abstracting the water allocated for the irrigation of olive groves. It clearly shows that the water quantity used for olive grove irrigation alone would be enough to maintain river flow during the dry season.

Evrotas River discharge during the irrigation period at Vrodamas for two different water abstraction states: a measured average river discharge for the period 1998–2000 and b hypothetical river discharge without the intense irrigation scheme conditions
Impacts of water stress on biota
Physicochemical parameters and pollutant concentrations at the surveyed reaches for the period 2006–2007 (just before fish sampling initiated) were below levels that may impact fish assemblages. In fact, the chemical-physicochemical status of the Evrotas main stem reaches used for fish sampling was good apart from the most downstream site (Skoura) that was classified as moderate (Skoulikidis 2008a). Moreover, the oxygen concentration during fish samplings never dropped below 5 mg/L, while the mean oxygen concentration in summers 2007 and 2008 in the four reaches examined ranged between 6.3 and 8.8 mg/L, indicating satisfactory oxygenation conditions. In addition, it was observed that the sites selected for macroinvertebrate sampling were not affected by pollution, thus ensuring that any differences in assemblages may result solely from hydrological changes. All sites selected were well oxygenated (dissolved oxygen ≥7 mg/L, average 8.7 mg/L) and their nutrient quality ranged, according to the classification system of Skoulikidis et al. (2006), between high (9 sites) and good (2 sites).
The start of ichthyological research coincided with the end of a period of hydrologically wet years; thus we assume that adequate time had been provided for fish communities to recover from past drought events. Therefore, during our first sampling period (spring 2007), fish communities can be characterised as relatively undisturbed in terms of structural integrity. This holds partially true for R2, a portion of which dried during the summer 2006. Figure 5 shows the fish assemblage composition and the areal fish densities of the three regional endemics in the two perennial and two intermittent reaches sampled during high flow (spring 2007 and spring 2008) and low flow (summer 2007 and summer 2008) periods. Figure 6 presents mean and variance values, derived from site-specific areal densities, for the three endemics over the same period in intermittent and perennial sites. Data show that density fluctuations were more intense in the intermittent than in the perennial sites. This drought event, had a dramatic effect on fish communities, eliminating all fish in the examined intermittent sites, and also affecting fish community attributes in the perennial sites due to flow and water volume reduction. By spring 2008, fish had begun to recolonise the intermittent sites. Species richness was restored as all species where present in the intermittent sites, but areal densities were extremely low compared to densities in the perennial sites (p < 0.05) and to those in the intermittent sites during to the previous spring (p < 0.05), as the two-way ANOVA analysis revealed. Thereafter, fish densities increased substantially and by the summer of 2008 they were similar (p > 0.05) with the density levels in the perennial sites. However, as Fig. 7 demonstrates, this increase was mainly due to recruitment of YOY following spring reproduction. Apparently, spring spawning and downstream larvae drift played a key role in the recolonisation process as populations in the intermittent reaches where composed chiefly of YOY with only a small proportion of the total composed of adults. Despite the enormous increase in densities, fish communities in the intermittent reaches were still disturbed after the drought event in terms of percentage species composition. For example, the stagnophilic, small-bodied and short-lived P. laconicus (an r-strategist) was positively affected from the drought event as its average percentage in the intermittent sites increased from 11% in spring 2007 to 43.3% in spring 2008 and reached up to 62.5% in the summer of the same year. In contrast, the rheophilic, larger-bodied and longer-lived S. keadicus decreased (in terms of percentage composition) from 63.2% in spring 2007 to 41.4% in spring 2008 and declined to 14.8% in the summer of 2008. In the perennial reaches, the percentage composition of both species in the spring of 2007 and 2008 remained relatively similar (for P. laconicus 58.1 and 68.6% and for S. keadicus 34.6 and 30.4%) while during the summers of 2007 and 2008 marked fluctuations were observed (P.laconicus increased from 21.3 to 62.8% while S. keadicus decreased from 60.5 to 29.5%), thus showing similar trends as with the intermittent sites.

Total abundance (areal density) of the three endemic fish species and comparisons between high (spring 2007 and 2008) and low flow periods (summer 2007 and 2008) in four river reaches representing hydrologically homogeneous river segments

Site-specific areal densities (inds/km2) (including means and variance values) for the three endemics fish species between high (spring 2007 and spring 2008) and low flow periods (summer 2007 and 2008)

Proportion of YOY of the two large-bodied fish of Evrotas (S. keadicus and T. spartiaticus) between high (spring 2007 and 2008) and low flow periods (summer 2007 and 2008)
Cluster analysis (Fig. 8a), performed for macroinvertebrates assemblages of intermittent RM4 streams revealed two main cluster groups with a similarity of approximately 70%, representing mainly spring samples (A) and winter samples (C). The distinction of the macroinvertebrate community into two main cluster groups was mostly attributed to seasonality. For example, Odonata were well represented in spring samples compared to winter samples, while the reverse was observed for Plecoptera. Moreover, Hirudinea were only present in winter samples. No marked seasonal differences in abundance and distribution were observed for the orders Ephemeroptera, Trichoptera, Diptera, Coleoptera and Heteroptera. In contrast to intermittent streams, faunal assemblages of perennial streams did not show marked differences between seasons and within sites (Fig. 8b), thus suggesting a stable and well-established community.

Dendrogram presenting benthic macroinvertebrate fauna similarities between and within sites and seasons in intermittent (a) and perennial streams (b). Numbers represent sites and letters seasons (A spring, B summer, C winter)
ANOSIM indicated that there are no differences in assemblages between intermittent and perennial streams (R = 0.063, p < 0.05). Furthermore, no differences were observed between pre-drought and post-drought assemblages in the intermittent sites (R = 0.178, p < 0.05). Differences were detected in the mean percentage of EPT taxa between perennial and intermittent streams as showed by a t test (t = −2.36, d.f. = 25, p = 0.02). However, the mean percentage of EPT taxa did not vary considerably between seasons in perennial (spring—55 ± 33%; summer—56 ± 16%; winter—51 ± 19%) and intermittent (spring—69 ± 20%; winter—75 ± 10%) mountainous streams (t test, p > 0.05). Seasonal changes in the mean percentages of COH taxa were insignificant in intermittent streams (spring—3.9 ± 3.2%; winter—3.1 ± 2.4%) and in perennial streams (spring—3.8 ± 2.2%; summer—15.4 ± 19%; winter—6.5 ± 7.8%) (Fig. 9). In perennial streams, the mean percentage of COH, which comprises many limnophylic taxa, appears to be markedly higher during summer as a result of high variability between sites. Regarding taxon richness, overall, no marked differences were found between perennial and intermittent streams (mean number of families in perennial and intermittent streams was 21 and 22, respectively). Finally, regarding the ratio EPT/(EPT + COH) no differences were observed among intermittent and perennial sites (t test, p = 0.11).

Mean, minimum and maximum of the EPT (Ephemeroptera, Plecoptera, Trichoptera) and COH (Coleoptera, Odonata, Heteroptera) taxa for perennial and intermittent streams
Discussion
Water stress conditions are particularly pronounced in the Mediterranean basin where river runoff follows cyclic seasonal patterns and shows declining trends during the last 50 years (Gasith and Resh 1999; Ludwig et al. 2009). In years with exceptionally prolonged drought, historically perennial rivers may now desiccate in the summer as a result of rainfall decrease, diminishing snow cover, high evapo-transpiration losses, and increased exploitation of water resources (Meybeck 2003; Benejam et al. 2010). In the Evrotas basin, a dramatic recent decrease in discharge is evident; the highest among major Balkan rivers (Skoulikidis 2009). During particularly prolonged extreme drought periods (e.g., 1988–1993), the vast majority of the river dries out in the summer and this was repeated during the extreme drought year of 2007. Although 2006 was exceptionally dry, the spatial extent of desiccation was limited in particular reaches since groundwater level was not yet affected. In summer 2008, despite the increase in rainfall, the spatial extent of desiccation was identical to 2007, since groundwater tables had not recovered from large-scale abstraction and drought over the preceding years.
Seasonal river-bed desiccation occurs naturally in the Evrotas basin within a small part of the downstream portion of the river’s main-stem (Vrodamas gorge) where surface runoff locally disappears due to infiltration in karstic aquifers. This may be part of the reason why the Evrotas as a whole has been erroneously considered as a naturally intermittent river. The following evidence reveal that the driving force of widespread desiccation is the excessive water abstraction, and it is, therefore, not solely a natural response to climatic drought. These are: (a) historical data obtained through the Leitbild approach, indicate that the Evrotas main-stem, its main tributary Oinus, and a number of spring-fed tributaries maintained perennial flow before the 1950s. This argument is further supported by the fact that few decades ago fish were present in most of the Evrotas tributaries where they no longer exist (Economou et al. 1999b); (b) there is a remarkable difference between rainfall reduction (10.5%) and discharge decrease (51.2%) when comparing an early to a more recent period of rainfall and discharge time-series. In addition, the estimated rainfall elasticity to stream flow in the basin is very low compared to other catchments elsewhere in the world, which indicates that factors other than long-term rainfall changes are mostly responsible for the identified decrease in the Evrotas’ discharge; (c) the comparison of the current water balance of the basin to the theoretical one prior to the recent irrigation of olive groves reveals a three-fold discharge difference during the dry period. This means that during dry periods, prior to the intensive agricultural abstractions, an amount of at least 6 m3/s would be available for surface runoff and infiltration in the catchment area. Therefore, according to this scenario and depending on the local soil moisture conditions, the vast majority of Evrotas River main course would probably maintain its perennial character throughout the dry period; and, (d) the abrupt desiccation of the Evrotas, upstream of Sparta during the summer of 2007 coincided with a period of intense surface-water and groundwater abstraction for irrigation. Considering that half a century ago the irrigated agricultural land was significantly less extensive than today, it is realistic to assume that at that time the discharge of the river was sufficient to maintain perennial flow during the summer.
Natural droughts are considered fairly regular and frequent events in Mediterranean lotic ecosystems to which many native species have built resilience through physiological and life-history adaptations (Williams 1996; Poff 1997; Gasith and Resh 1999; Magoulick and Kobza 2003; Magalhaes et al. 2007; Matthews and Matthews 2003). In contrast, an artificial increase in the frequency or severity of water stress due to water abstraction may cause ecological perturbations to which species are evolutionarily inexperienced, and may result in drastic changes in aquatic biocommunities, including widespread fish kills (Stanley et al. 2004; Magalhaes et al. 2007). Overcrowding in reduced space is also known to adversely influence native biota survival due to habitat degradation, water quality deterioration and increased biotic interactions (competition and predation) (reviewed by Magoulick and Kobza 2003; Magalhães et al. 2002). Results from this study show that the summer 2007 drought generated impacts on fish community attributes that persisted after the drought event has passed. The most obvious and immediate impact was the elimination of populations in the intermittent reaches. Drought impacts were not limited to the intermittent reaches; in associated studies we found that populations in the perennial reaches also experienced stressful conditions (extreme temperature, hypoxia, strengthened biotic interactions etc.) resulting in elevated mortality (Skoulikidis 2008a; Nikolaidis et al. 2009).
Following flow resumption, fish communities began to recover (where recovery is defined as structural and functional restoration of the fish community to a state comparable to that in the pre-drought periods or in the perennial reaches). The initial trigger for recovery was recolonisation that led to the re-establishment of fish in the formerly desiccated reaches. Various studies have shown that post-drought recolonisation rates depend on a large variety of factors, including regional and biogeographical characteristics, the intensity of the drought episode, the presence of aquatic refuges, the distance from refuges and the life cycle characteristics of species (Niemi et al. 1990; Detenbeck et al. 1992; Adams and Warren 2005; Lake 2003). Local topography may further inhibit fish movements due to barriers to fish migration or dispersal. Our data indicate that recolonisation of the intermittent reaches was accomplished through both immigration and reproduction. From autumn to spring, recolonisation was effected through migration or downstream transport during winter flood flows. During this period, rates of recolonisation were low and resulted in the establishment of small populations of adults and late juveniles in the spring of 2008. Subsequent recolonisation succeeded the late reproductive period (spring and early summer) and was from YOY, which probably drifted from upstream perennial refuges. However, the possibility that some of the fry were produced by early colonists already present in spring in the intermittent reaches cannot be excluded. This colonisation mode led to an enormous increase of fish densities from spring to summer 2008. In fact, the increase was so high that fish densities in the intermittent reaches in the summer of 2008 far exceeded those in the perennial reaches. This enormous increase of densities could have resulted from a combination of three conditions: (a) a “concentration effect” due to the dramatic reduction of the wetted surface area, (b) significant recruitment of YOY fish following the spring reproduction (May–early June), and (c) higher fry survival rates in the intermittent than in the perennial reaches due to low predation by larger fish and/or other large predators. Whereas all three conditions probably contributed to the increase of summer densities, the dominance of early juveniles in the age structure of the S. keadicus and T. spartiaticus populations suggests that spring reproduction played the major role (Fig. 7).
Whereas recolonisation of formerly desiccated reaches appeared to be a relatively rapid phenomenon accomplished primarily through reproductive processes, the restoration of the fish community to a relatively undisturbed state is a slower and more complex process. This involves various community and population attributes that recover at different speeds. Species richness recovered first, and by May 2008 (6–7 months after the extreme drought event) all three species were present in the samples. In summer 2008 abundance had also largely recovered. However, the percentage species composition and age structure was still disturbed in relation to the pre-drought period. The prominent feature of the fish community in the intermittent reaches in summer 2008 was that populations were composed chiefly of small, 2–4 month old fry. The high densities of these tiny fish may somehow mislead interpretations about the effectiveness or completeness of the recovery process: fry composition and densities in the intermittent reaches are more likely to reflect rates of fry arrival from upstream, rather than habitat-related establishment process and survival rates. Thus, despite extensive recolonisation, full recovery had not been accomplished, and communities were still impacted, by the end of our investigation period. Our evidence from ongoing research shows that it takes at least three “normal hydrological” years following a severe drought episode for population age structure to fully recover. Consequently, it is this relatively slowness to recovery that makes fish particularly useful indicators of hydrological regime alterations since effects of past hydrological disturbances on fish communities can be detected long after the disturbance is over.
Overall, the fish fauna of the Evrotas River appears to be fairly resilient to drought-driven reach-scale desiccation and ultimate recover, provided that aquatic refuges are available to supply adult colonists and no barriers to impede recolonisation exist. These conditions hold for most of the river main-stem but not for the majority of tributary reaches where absence of refugium populations, irreversible hydromorphological changes and existence of weirs or other barriers to immigration have prevented recolonisation and recovery. The data presented in the present paper as well as those reported elsewhere (Skoulikidis 2008a; Nikolaidis et al. 2009) demonstrate that desiccation impacts on fish communities are widespread and increasingly common and severe within the Evrotas basin. This is evident by the following: (a) during the 2007 drought episode all except a few tributary sections dried-out completely (i.e., about 80% of the length of main river stem); (b) drought events appear with increasingly higher frequency and over larger spatial scales in recent decades; and (c) fish communities in most tributary streams have persistently failed to recover despite a series of normal hydrological years.
In contrast to the fish fauna, the macroinvertebrate fauna of the intermittent reaches of the river system do not seem to be markedly affected by summer river-bed desiccations. The post-drought macroinvertebrate community was relatively similar to the pre-drought community (c. 70% similarity). This suggested that there were no significant changes in the structure and composition of the macroinvertebrate fauna after summer desiccation in intermittent sites. The distinction of the macroinvertebrate community into two main cluster groups was evidently attributed to seasonality.
Generally, species richness of aquatic insects is lower in intermittent than in perennial lotic waters (Williams 1996). A study conducted in an intermittent and a perennial stream of NE Spain, showed that marked seasonality appeared to be an important determinant of invertebrate community structure, however, fauna richness was lower in the intermittent than in the perennial stream (Munoz 2003). Prolonged and severe droughts may have greater effects on benthic fauna or on certain benthic groups, as documented in arid and semi-arid regions (e.g., in so-called arid-land or dryland rivers). For example, in two Australian intermittent streams, stoneflies and caseless caddisflies did not recruit successfully the year following a drought despite flow return (Boulton 2003). In this study, richness variation and macroinvertebrate assemblages were not apparent between perennial and intermittent streams. Furthermore, no differences were observed between number of families, EPT and COH taxa. A similar study, carried out by Bonada et al. (2007), showed that annual and seasonal macroinvertebrate richness, EPT and COH metrics did not differ between perennial and intermittent sites. Miller and Golladay (1996) observed similar number of taxa in both stream types, results which are consistent with the results of this study. In intermittent streams, aquatic organisms are subject to severe environmental stresses, such as long droughts and floods; however, macroinvertebrates have countered the recolonisation of their habitats by means of physiological tolerance, dormancy, migration and life history modifications (Williams 1997). In the Evrotas River, the macroinvertebrate fauna recovered successfully, presumably mainly due to downstream dispersal from reaches and pools that retained water, as well as due to their various drought-adaptation mechanisms.
Our knowledge of the drought effects on macroinvertebrate species and their assemblages in Mediterranean intermittent rivers is localised and usually opportunistic (Boulton and Lake 2008). The assessment of direct or indirect effects of drought on macroinvertebrates requires long-term monitoring. Considering the historically perennial character of the Evrotas main course and of most of its tributaries, and taking into account the lack of historical biotic data, it cannot be excluded that repeated and prolonged water stress conditions during the recent past have already altered the initial macroinvertebrate species and communities towards more drought-resistant depauperate assemblages.
Despite the fact that flow regime alteration constitutes a widespread and pervasive threat to streams (Bunn and Arthington 2002), we are still lacking a basic understanding of the processes by which artificial desiccation affects lotic ecosystems. In this study, we attempted to address this important knowledge gap by exploring the consequences of artificial desiccation on aquatic biotic elements. Clearly, one of the most important questions addressed when assessing the status of an intermittent river is to identify whether seasonal desiccation is driven by natural variability in surface-water permanence or by water abstraction; yet, this is often neglected in many policy-relevant assessment applications. In river basins which lack historical hydrological data, the distinction between natural and anthropogenic desiccation is usually not clear. Hence, one of the most challenging aspects of our assessment procedure was the estimation of the river’s hydrological reference conditions, prior to agricultural intensification. The Leitbild approach to researching reference condition attributes of the river helped us identify modern human hydromorphological impacts. Our multidisciplinary hydrological and biological research allowed us to provide evidence for the notion of an artificially intermittent state in much of the Evrotas’ river network, something which was not recognised in past assessments of this river (Angelidis et al. 1995).
This kind of anthropogenic river desiccation is referred to as “artificially induced desiccation” and “neo-arheism” in Meybeck (2003). Intermittent rivers and streams of this type are rarely mentioned by specific terminology in the literature: Uys and O’Keeffe (1997) describe them as “modified temporary”; Larned et al. (2008) as “anthropogenic intermittent” and Benejam et al. (2010) as “artificially dry”. We, therefore, refer to the Evrotas as an “artificially intermittent” river highlighting this prominent artificial alteration in surface-water conditions. In such anthropogenically stressed rivers, the extent of surface water desiccation varies substantially from year to year, since it is directly related to the combination of climate patterns and meteorological events, which in turn affect the degree of water abstraction for irrigation.
The findings of this study support the idea of a distinction between natural and anthropogenic desiccation for management and conservation purposes. Here we introduce the need to identify a “Mediterranean artificially intermittent” condition as a distinct sub-category of intermittent Mediterranean rivers. In order to restore ecosystem health in artificially intermittent rivers and streams, the assessment of ecological status and management strategies should be appropriately developed. Hence, in the context of WFD implementation, if a perennial reach turns to intermittent due to water exploitation, then its status should be assessed bearing in mind that reference conditions are based on perennially flowing conditions and associated biota. Thus, during the period of complete desiccation, the status of river reaches that dry-out artificially should score “bad” since fish have been extirpated. This is in agreement with Benejam et al. (2010), who state that in cases of artificially dry rivers, biotic indices should be given the worst score instead of the customary missing value that biases the overall ecological assessment. Assigning a bad status during artificial desiccation, the overall ecological status during the operational monitoring within the WFD scheme may score lower than good. This will require the restoration of ecological flow regimes in the framework of the implementation of measures within the River Basin Management Plans.
Sensitivity of flow regime alterations may be further complicated by gradual changes in climatic conditions. Climate change projections for the Mediterranean (Isendahl and Schmidt 2006), complemented by a growth in water demand (Tockner et al. 2009), will generally result in a river flow reduction (EEA 2008), thus increasing water stress (Cudennec et al. 2007; Tockner et al. 2009). Water stress will ultimately affect the low flow regimes causing artificial desiccation of river surface water features during the dry season. Thus, the number and length of artificially intermittent rivers is expected to rise (Larned et al. 2010). According to Meybeck (2003), even the world’s major rivers located in semi-arid and arid regions will face a seasonally artificially induced desiccation if a minimum regulated flow is not maintained. This will exert major impacts on aquatic quality and biodiversity (Sabater and Tockner 2010) and may threaten entire fluvial ecosystems (Daufresne and Boët 2007). Thus, artificially intermittent streams will increasingly become a research priority with multiple management implications, especially in semi-arid regions.
References
Adams SB, Warren ML (2005) Recolonization by warm water fishes and crayfishes after severe drought in upper coastal plain hill streams. T Am Fish Soc 134:1173–1192. doi:10.1577/TfW-089.1
Angelidis MO, Markantonatos PG, Bacalis NC (1995) Impact of human activities on the quality of river water: the case of Evrotas River catchment Basin, Greece. Environ Monit Assess 35:137–153. doi:10.1007/BF00633711
AQEM Consortium (2002) Manual for the application of the AQEM method. A comprehensive method to assess European streams using benthic macroinvertebrates, developed for the purpose of the Water Framework Directive. Version 1.0, February 2002, pp 202
Benejam L, Angermeier PL, Munne A, Garcia-Berthou E (2010) Assessing effects of water abstraction on fish assemblages in Mediterranean streams. Freshwater Biol 55:628–642. doi:10.1111/j.1365-2427.2009.02299.x
Bizas C (1995) Restoration and management of European riparian landscapes: the case of Evrotas’ riparian area. The European Association for Environmental Management Master’s thesis, pp 100
Bobori DC, Economidis PS (2006) Freshwater fishes of Greece: their biodiversity, fisheries and habitats. Aquat Ecosyst Health 9(4):407–418. doi:10.1080/14634980601027855
Bonada N, Rieradevall M, Prat N, Resh VH (2006) Benthic macroinvertebrate assemblages and macrohabitat connectivity in Mediterranean-climate streams of Northern California. J N Am Benthol Soc 25:32–43. doi:10.1899/0887-3593
Bonada N, Doledec S, Statzner B (2007) Taxonomic biological trait differences of stream macroinvertebrate communities between Mediterranean and temperate regions: implications for future climatic scenarios. Glob Change Biol 13:1658–1671. doi:10.1111/j.1365-2486
Bond NR, Lake PS, Arthington AH (2008) The impact of droughts on freshwater ecosystems: an Australian perspective. Hydrobiologia 600:3–16. doi:10.1007/s10750-008-9326-z
Bostelmann R, Braukmann U, Briem E, Fleischhacker T, Humborg G, Nadolny I, Scheurlen K, Weibel U (1998) An approach to classification of natural streams and floodplains in South-west Germany. In: de Waal LC, Large ARG, Wade PM (eds) Rehabilitation of rivers: principles and implementation. Wiley, Chichester, pp 31–55
Boulton AJ (2003) Parallels and contrasts in the effects of drought on stream macroinvertebrate assemblages. Freshwater Biol 48:1173–1185
Boulton AJ, Lake PS (1992) The ecology of two intermittent streams in Victoria, Australia: III. Temporal changes in faunal composition. Freshwater Biol 27(1):123–138
Boulton AJ, Lake PS (2008) Effects of drought on stream insects and its ecological consequences. In: Lancaster J, Briers RA (eds) Aquatic insects: challenges to populations. CAB International, Wallingford, pp 81–102. doi:10.1046/j.1365-2427
Bradford RB (2000) Drought events in Europe. In: Vogt JV, Somma F (eds) Drought and drought mitigation in Europe. Kluwer Academic Publishers, Dordrecht, Netherlands, p 319
Bunn SE, Arthington AH (2002) Basic principles and ecological consequences of altered flow regimes for aquatic biodiversity. Environ Manage 30(4):492–507. doi:10.1007/s00267-002-2737-0
Cavanagh W, Crouwel J, Catling RWV, Shipley G (2002) Continuity and change in a greek rural landscape: the laconia survey—methodology and interpretation, vol I. Annual of the British School at Athens, London
CEN (European Committee for Standardization) (1994) Water quality - Methods of biological sampling: Guidance on hand net sampling of aquatic benthic macro-invertebrates. EN 27828:1994 E
Chadzichristidi K, Beltsios S, Papakonstantinou A (1991) Pinios River water quality measurements. 2nd Conference on Environmental Science & Technology, Lesvos September, 630–639 (in Greek with English abstract)
Chiew FHS (2006) Estimation of rainfall elasticity of streamflow in Australia. Hydrol Sci J 51(4):613–625. doi:10.1623/hysj.51.4.613
Clarke KR, Gorley RN (2006) PRIMER v6: User Manual/Tutorial. PRIMER-E, Plymouth, p 190
Cudennec C, Leduc C, Koutsoyiannis D (2007) Dryland hydrology in Mediterranean regions: a review. Hydrolog Sci J 52(6):1077–1087
Daufresne M, Boët P (2007) Climate change impacts on structure and diversity of fish communities in rivers. Glob Change Biol 13:2467–2478. doi:10.1111/j.1365-2486
Demuth, S, Stahl K (eds) (2001) Assessment of the regional impact of droughts in Europe. Final Report to the European Union, ENV-CT97-0553, Institute of Hydrology, University of Freiburg, Germany
Detenbeck NE, DeVore P, Niemi GJ, Lima A (1992) Recovery of temperate-stream fish communities from disturbance: a review of case studies and synthesis of theory. Environ Manage 16:33–53. doi:10.1007/BF02393907
Dimopoulos P, Zotos A, Zogaris S (2009) Protocol for a rapid assessment of riparian zone conservation. Environmental Friendly Technologies for Rural Development, Final Report 2005–09, LIFE-ENVIRONMENT LIFE05ENV/Gr/000245 EE (EnviFriendly), pp 127–141
EEA (2000) Environmental signals: environmental assessment report No 6. Copenhagen, Denmark, p 108
EEA (2008) Impacts of Europe’s changing climate–indicator based assessment. Joint EEA-JRC-WHO report, EEA Report No 4/2008, JRC Reference Report No JRC47756, Copenhagen, Denmark, pp 207
EEA (2009) Water resources across Europe: confronting water scarcity and drought. EEA Report No 2, Copenhagen, Denmark, pp 55
EC (2007) WFD intercalibration technical report. MedGIG Intercalibration technical report: Part 1 Rivers, Section 1 Benthic Invertebrates, 15 June 2007, pp 17
Economou A, Barbieri R, Daoulas C, Psarras T, Stoumboudi M, Bertahas H, Giakoumi S, Patsias A (1999a) Endangered freshwater fish of western Greece and the Peloponnese: Distribution, abundance, threats and measures for protection. Final Technical Report, National Centre for Marine Research, pp 341
Economou AN, Barbieri R, Stoumboudi M (1999b) Threatened endemic freshwater fishes to Greece: the Evrotas case. Workshop on “Wetland Restoration”, 12–14 November 1999, Gythion, Greece
Economou AN, Giakoumi S, Vardakas L, Barbieri R, Stoumboudi M, Zogaris S (2007a) The freshwater ichthyofauna of Greece: an update based on a hydrographic basin survey. Mediterranean Marine Sci 8(1):91–168
Economou AN, Zogaris S, Chatzinikolaou Y, Tachos V, Giakoumi S, Kommatas D, Koutsikos N, Vardakas L, Blasel K, Dussling U (2007b) Development of an ichthyological multimetric index for ecological status assessment of Greek mountain streams and rivers. Technical Report. Hellenic Center for Marine Research – Institute of Inland Waters, Hellenic Ministry for Development, pp 166
Estrela T, Marcuello C, Iglesias A (1996) Water resources problems in southern Europe an overview report. European Topic Centre on Inland Waters, European Environment Agency, pp 45
Eurostat (2009) EuroMediterranean statistics. European Communities, Luxemburg, p 216
FAME (2005) Fish-based assessment method for the ecological status of European rivers—a contribution to the Water Framework Directive. Final Report; Manual for the application of the European Fish Index (EFI), pp 41
Gasith A, Resh VH (1999) Streams in Mediterranean climate regions: abiotic influences and biotic responses to predictable seasonal events. Annu Rev Ecol Syst 31:51–58. doi:10.1146/annurev.ecolsys.30.1.51
Gordon ND, Mcmahon TA, Finlayson BL, Gipple CJ, Nathan RJ (2004) Stream hydrology: an introduction for ecologists, 2nd edn. Wiley, West Sussex
Gore JA (1982) Benthic invertebrate colonization: source distance effects on community composition. Hydrobiologia 94:183–193
Hamon WR (1961) Estimating potential evapotranspiration. J Hydraul Div Proc Am Soc Civil Eng 87:107–120
Hillel D (1980) Fundamentals of Soil Physics. Academic Press, New York
Isendahl N, Schmidt G (2006) Drought in the Mediterranean: WWF Policy Proposals, WWF/Adena, WWF Mediterranean Programme, WWF Germany
Jacobson PJ, Jacobson KM, Angermeier PL, Cherry DS (2004) Variation in material transport and water chemistry along a large ephemeral river in the Namid Desert. Freshwater Biol 44:481–491. doi:10.1046/j.1365-2427
Lake PS (2000) Disturbance, patchiness, and diversity in streams. J N Am Benthol Soc 19:573–592
Lake PS (2003) Ecological effects of perturbation by drought in flowing waters. Freshwater Biol 48:1161–1172. doi:10.1046/j.1365-2427
Larned S, Arscott D, Datry T, Tockner K (2008) Special session—intermittent rivers: global phenomena and unexplored riverscapes I. The North American Benthological Society 56th Annual Meeting, 25–28 May, 2008
Larned S, Datry T, Arscott D, Tockner K (2010) Emerging concepts in temporary-river ecology. Freshwater Biol 55(4):717–738. doi:10.1111/j.1365-2427.2009.02322.x
Ludwig W, Dumont E, Meybeck M, Heussner S (2009) River discharges of water and nutrients to the Mediterranean and Black Sea: Major drivers for ecosystem changes during past and future decades? Prog Oceanogr 80:199–217. doi:10.1016/j.pocean
Magalhães MF, Batalha DC, Collares-Pereira MJ (2002) Gradients in stream fish assemblages across a Mediterranean landscape: contributions of environmental factors and spatial structure. Freshwater Biol 47:1015–1031. doi:10.1046/j.1365-2427.2002.00830.x
Magalhaes MF, Beja P, Schlosser IJ, Collares-Pereira MJ (2007) Effects of multi-year droughts on fish assemblages of seasonally drying Mediterranean streams. Freshwater Biol 52:1494–1510. doi:10.1111/j.1365-2427
Magoulick DD, Kobza RM (2003) The role of refugia for fishes during drought: a review and synthesis. Freshwater Biol 48:1186–1198. doi:10.1046/j.1365-2427
Mathews WJ (1998) Patterns in freshwater fish ecology. Chapman & Hall, New York
Matthews WJ, Matthews EM (2003) Effects of drought on fish across axes of space, time and ecological complexity. Freshwater Biol 48:1232–1253. doi:10.1046/j.1365-2427.2003.01087.x
Meybeck M (2003) Global analysis of river systems: from Earth system controls to Anthropocene symptoms. Phil Trans R Soc Lond 358(1140):1935–1955. doi:10.1098/rstb
Miller AM, Golladay SW (1996) Effects of spates and drying on macroinvertebrate assemblages of an intermittent and a perennial prairie stream. J N Am Benthol Soc 15:670–689
Munoz I (2003) Macroinvertebrate community structure in an intermittent and a permanent Mediterranean stream (NE Spain). Limnetica 22:107–116
Niemi GJ, DeVore P, Detenbeck N, Taylor D, Lima A, Pastor J, Yount JD, Naiman RJ (1990) Overview of case studies on recovery of aquatic systems from disturbance. Environ Manage 14:571–587. doi:10.1007/BF02394710
Nikolaidis N, Skoulikidis N, Kalogerakis N, Tsakiris K (2009) Environmental Friendly Technologies for Rural Development, Final Report 2005–09, LIFE-ENVIRONMENT LIFE05ENV/Gr/000245 EE (EnviFriendly)
Norrant C, Douguédroit A (2006) Monthly and daily precipitation trends in the Mediterranean (1950–2000). Theor Appl Climatol 83:89–106. doi:10.1007/s00704-005-0163-y
OECD (2000) Environmental indicators: towards sustainable development, technical report, organization for economic cooperation and development, pp 132
Otermin A, Basaguren A, Pozo J (2002) Re-colonization by the macroinvertebrate community after a drought period in a first-order stream (Agüera Basin, Northern Spain). Limnetica 21(1–2):117–128
Papazafeiriou ZG (1999) Water demands of crops. Ziti Publications, Thessaloniki
Poff NL (1997) Landscape filters and species traits: towards mechanistic understanding and prediction in stream ecology. J N Am Benthol Soc 16:391–409
Ryder DS, Boulton AJ (2005) Redressing the limnological imbalance: trends in aquatic ecology, management and conservation in Australia. Hydrobiologia 552:159–166. doi:10.1007/s10750-005-1513-6
Sabater S, Tockner K (2010). Effects of hydrological alterations on the ecological quality of river ecosystems. In: Sabaterand S, Barcelo D (eds.) Water scarcity in the mediterranean: perspectives under global change. Springer, Heidelberg
Skliris N, Sofianos S, Lascaratos A (2007) Hydrological changes in the Mediterranean Sea in relation to changes in the freshwater budget: a numerical modelling study. J Marine Sys 65(1–4):400–416. doi:10.1016/j.jmarsys.2006.01.015
Skoulikidis N (editor), Economou A, Karaouzas I, Vardakas L, Gritzalis Κ, Zogaris S, Dimitriou E, Tachos V (2008a) Hydrological and biogeochemical monitoring in evrotas basin. Final technical report 1, H.C.M.R. LIFE-ENVIRONMENT: LIFE05 ENV/GR/000245 « ENVIRON-MENTAL FRIENDLY TECHNOLOGIES FOR RURAL DEVELOPMENT
Skoulikidis N (editor), Economou A, Karaouzas I, Vardakas L, Gritzalis Κ, Zogaris S, Dimitriou E, Tachos V (2008b) Risk assessment of water management in Evrotas River Basin. Final technical report 2, H.C.M.R. LIFE-ENVIRONMENT: LIFE05 ENV/GR/000245 « ENVIRON-MENTAL FRIENDLY TECHNOLOGIES FOR RURAL DEVELOPMENT
Skoulikidis Ν (2009) The environmental state of rivers in the Balkans: a review within the DPSIR framework. Sci Total Environ 407:2501–2516. doi:10.1016/j.scitotenv
Skoulikidis N, Amaxidis Y, Bertahas I, Laschou S, Gritzalis K (2006) Analysis of factors driving stream water composition and synthesis of management tools–a case study on small/medium Greek catchments. Sci Total Environ 362:205–241. doi:10.1016/j.scitotenv.2005.05.018
Smakthin VU (2003) Low flow hydrology: a review. J Hydrol 240:147–186. doi:10.1016/S0022-1694(00)00340-1
Smith B (1997) Water: a critical resource. In: King R, Proudfoot L, Smith B (eds) The mediterranean: environment and society. Arnold, London, pp 227–251
Stanley EH, Fisher SG, Jones JB (2004) Effects of water loss on primary production: a landscape-scale model. Aquat Sci 66:130–138. doi:10.1007/s00027-003-0646-9
TempQsim Consortium (2006) Critical issues in the water quality dynamics of temporal rivers—evaluation and recommendations of the tempQsim project. In: Froebrich J, Bauer M (eds) Enduser Summary. Hannover, Germany
Tockner K, Uehlinger U, Robsinson CT, Tonolla D, Silber R, Peter FD (2009) Introduction to European Rivers. In: Tockner K, Uehlinger U, Robsinson CT (eds) Rivers of Europe. Academic Press, London, pp 1–21
Tramer EJ (1977) Catastrophic mortality of stream fishes trapped in shrinking pools. Am Midl Nat 97:469–478
UNEP (2004) Freshwater in Europe - Facts, Figures and Maps UNEP report on the state of Freshwater in Europe and Central Asia
UNEP/MAP (2003) Riverine transport of water, sediments and pollutants to the Mediterranean Sea. UNEP/Mediterranean Action Plan, Athens, Greece
UNESCO (1979) Map of the world distribution of arid regions. Map at scale 1:25, 000, 000 with explanatory note. UNESCO, Paris
Uys ΜC, O’Keeffe JH (1997) Simple Words and Fuzzy Zones: Early Directions for Temporary River Research in South Africa. Environ Manage 21(4):517–531. doi:10.1007/s002679900047
Vagiakakos DB (1989) Evrotas, terminology. Proceedings of the Special Literary Symposium, 27–29 May 1989, Sparta-Mystras, pp 361–414
Wagstaff M (2002) The formation of the modern landscape of the survey area, Continuity and change in a Greek rural landscape “The Laconia Survey”, vol 1. Methodology and Interpretation. In: Cavanagh WG and Walker SEC (eds) The council, British School of Athens, London, pp 403–420
Ward R (1975) Principles of hydrology. McGraw Hill, UK
Williams D (1996) Environmental constraints in temporary fresh waters and their consequences for the insect fauna. J N Am Benthol Soc 15(4):634–650. doi:10.2307/1467813
Williams D (1997) Temporary ponds and their invertebrate communities. Aquat Conserv 7:105–117. doi:10.1002/(SICI)1099-0755
World Bank (2003) Water Resources Management in Europe. Environmentally and socially sustainable development department Europe and Central Asia Region. Vol. I. Issues and directions, Vol. II. Country water notes and water fact sheets, Washington, USA
Zacharias I, Dimitriou E, Koussouris T (2003) Developing sustainable water management scenarios by using thorough hydrologic analysis and environmental criteria. J Environ Manage 69:401–412. doi:10.1016/j.jenvman
Zogaris S, Economou AN, Dimopoulos P (2009) Ecoregions in the Southern Balkans: should they be revised? Environ Manage 43:682–697. doi:10.1007/s00267-008-9243-y
Acknowledgments
The data for this article is derived from the LIFE-EnviFriendly Project (LIFE05 ENV/GR/000245): “Environmental friendly technologies for rural development” (http://www.envifriendly.tuc.gr), which was carried out in the Evrotas River Basin during 2005–2009. We would like to express our gratitude to V. Papadoulakis (Lakonia Prefecture), who provided historical hydro-meteorological time series and to Andriopoulou A., Koutsodimou M., Kouvarda T., Akepsimaidis K., Tachos V., Kommatas D., Koutsikos N., Chatzinikolaou Y., Charalabous I. and Economou E. for their assistance in field work and laboratory analysis.
Author information
Authors and Affiliations
Corresponding author
Additional information
This article belongs to the Special Issue "Recent Perspectives on Temporary River Ecology".
Rights and permissions
About this article
Cite this article
Skoulikidis, N.T., Vardakas, L., Karaouzas, I. et al. Assessing water stress in Mediterranean lotic systems: insights from an artificially intermittent river in Greece. Aquat Sci 73, 581–597 (2011). https://doi.org/10.1007/s00027-011-0228-1
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00027-011-0228-1