
Abstract
Macrophages are tissue-resident immune cells that play a critical role in maintaining homeostasis and fighting infection. In addition, these cells are involved in the progression of many pathologies including cancer and atherosclerosis. In response to a variety of microenvironmental stimuli, macrophages can be polarized to achieve a spectrum of functional phenotypes. This review will discuss some emerging evidence in support of macrophage phenotypic regulation by physical and mechanical cues. As alterations in the physical microenvironment often underlie pathophysiological states, an understanding of their effects on macrophage phenotype and function may help provide mechanistic insights into disease pathogenesis.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Macrophages are diverse and multifunctional immune cells that reside in all adult tissues. They are central to many biological processes, including development, metabolic regulation, maintenance of tissue homeostasis, and defense against invading pathogens [1]. Most tissue-specific macrophages are derived from the yolk sac during embryonic development and maintained through self-renewal [2]. In adults, hematopoiesis continues in the bone marrow to give rise to circulating blood monocytes. These monocytes can be recruited and differentiated to replenish tissue macrophages, usually at sites of injury [3]. In their basal states, macrophages maintain tissue homeostasis through the clearance of apoptotic cells and assist in tissue repair. However, upon exposure to various microenvironmental stimuli, macrophages can become activated and achieve many functional phenotypes. Due to its complexity, macrophage phenotype polarization remains the focus of much ongoing research and debate [4]. It is now widely accepted that macrophage phenotypes can be viewed as a continuum [5], comprising of multiple distinct functional states and each with its unique transcriptional profile [6]. In the most simplistic form, macrophage activation is often categorized into two extremes: classically activated (M1) and alternatively activated (M2). M1 macrophages, also known as proinflammatory, are generally instigated by the presence of Th-1 cytokines tumor necrosis factor- α (TNF-α) and interferon- γ (IFN-γ) as well as bacterial endotoxin lipopolysaccharide (LPS). This state is marked by high secretion levels of TNF-α, IL-12, and IL-23, cytokines that further propagate the inflammatory response in downstream immune cells. M1 macrophages also produce intracellular nitric oxide (NO) and reactive oxygen intermediates (ROI), which are cytotoxic mediators that help fight infection. On the other hand, secretion of IL-10, a known inhibitor of inflammation, is the hallmark of M2 macrophages. The M2 phenotype can be further subcategorized into M2a, M2b, and M2c, based on their specific yet overlapping functions. Although all three subtypes have anti-inflammatory properties, M2a and M2b macrophages are considered immunoregulatory and are known for mediating the Th-2 response, whereas M2c cells are immunosuppressive and engage in extracellular matrix (ECM) remodeling [7]. Additionally, M2a and M2c macrophages also secrete growth factors that promote angiogenesis and tissue regeneration [8].
The versatility of macrophages stems from their ability to shift phenotypic presentation in response to their ever-changing microenvironment. Aberrant phenotypic polarization, spatially or temporally, often differentiates physiological and pathological conditions [9–11]. Indeed, it has become increasingly recognized that macrophages are essential in the progression of pathophysiological conditions including cancer, cardiovascular diseases, obesity, wound healing, and foreign body response (FBR). However, experimental therapies using systemic depletion of macrophage cells have resulted in detrimental and often lethal consequences [12], as we now know that certain functional populations are necessary for proper wound healing and can impede disease progression. For instance in cancer, M1 macrophages are known to have tumoricidal functions, whereas M2-like macrophages aid tumor cells in evading destruction from host immune cells, and promote angiogenesis, invasion and metastasis. The majority of tumor-associated macrophages adopt an M2-like phenotype, and their presence in tumors has been directly correlated with poor prognosis [13, 14]. Contrarily in atherosclerosis, M1 macrophages are commonly viewed as atherogenic, while M2 macrophages are seen as atheroprotective. To realize the potential of macrophages as a therapeutic target for pathologies, a deeper understanding of macrophage phenotypic adaptation during disease pathogenesis is required.
Much of our current understanding on macrophage functional plasticity is limited to how soluble factors such as cytokines and chemokines influence polarization. The complexity of the microenvironments that macrophages are often present in underscores the necessity to consider other factors that may influence macrophage phenotype polarization. Alterations in the physical microenvironment such as extracellular matrix (ECM) stiffness, architecture and composition are hallmarks of both cancer and atherosclerosis. Seminal work in the past few decades have shown that these physical and mechanical signals can regulate differentiation, proliferation, and migration of many cell types. However, whether these factors play a role in coaxing macrophages into achieving the phenotype necessary for disease progression is still unclear. This review intends to highlight a few recent findings that suggest macrophages may be mechanically sensitive and that their phenotype may be regulated, in part, by physical cues. Macrophage involvement in cardiac diseases, cancer and wound healing are reviewed elsewhere [11, 15, 16]. This review will emphasize how changes in the physical microenvironment associated with disease states may be correlated with changes in macrophage phenotype and function.
Substrate topography and matrix architecture
Much of our current understanding of how physical cues influence macrophage phenotype has come from the field of biomaterials. Macrophages are among the first cells to interact with implanted biomaterials and are key mediators of the host foreign body response (FBR). Macrophage phenotype polarization during FBR is critical as M2 phenotype has been shown to be indicative of constructive remodeling around implanted materials [17, 18]. Therefore, the ability to modulate macrophage phenotype toward M2 polarization has emerged as a promising design strategy for biomaterials. Toward this end, many groups have attempted to exploit the material surface topography as a passive, alternative approach to biochemical surface modification (Fig. 1).

a Substrate topology such as micro- and nano-grooves and fibers can modulate macrophage adhesion, spreading and activation [25–27]. b Scanning electron microscopy images of murine RAW264.7 macrophages cultured on 2 μm-wide gratings and planar control surfaces [25]. Scale bar 20 μm. c Brightfield images of H&E stained RAW264.7 cells seeded on poly(L-lactic acid) (PLLA) planar and micro-aligned fibrous scaffold (fiber diameter = 1.6 ± 0.25 μm) for 7 days [26]. Scale bar 45 μm
It has long been observed that macrophages prefer to adhere to rough surfaces rather than flat [19]. More recently, researchers have examined whether surface roughness plays a role in macrophage secretion of inflammatory cytokines. In the absence of LPS stimulation, sandblasting and acid etching roughened titanium surfaces were shown to increase TNF-α secretion while decreasing production of chemoattractants monocyte chemotactic protein 1 (MCP-1) and macrophage inflammatory protein-1α (MIP-1α) in RAW264.7 macrophages. When activated with LPS, surface roughness synergistically upregulates secretion of all inflammatory cytokines including TNF-α, IL-1β, IL-6, MCP1, and MIP-1α. The surface roughness enhanced secretion was generally observed at the 24 or 48 h time point but not at an earlier, 6 h, time point, implying a temporal dependence of the topographical effect [20]. Contrarily, in murine J744A.1 macrophages, sandblasted and acid etched roughness in titanium showed no significant effect on the production of IL-1β, IL-6, IL-10 and nitric oxide (NO) [21]. However, nano-roughened titanium surfaces resulted in mitigated expression of TNF-α, IL-1β and NO in J744A.1 macrophages. In addition, nanorough substrates also restricted macrophage migration through reduction in migration speed and distance [22]. These seemingly incongruent findings may be attributed to differences in cell line, experimental time points, and perhaps the rather imprecise control of surface topology through roughening.
Using lithographic methods to more precisely control surface topography, studies have examined macrophage response to various polymers imprinted with grooves or gratings. On poly(methyl methacrylate) containing 10 μm-wide and 0.5 μm-deep grooves, murine P388D1 macrophages exhibit substantial cellular elongation and align along the grooves and ridges. The patterned substrate enhanced macrophage migration in terms of speed, distance and persistence [23]. Additionally, microgrooves promote phagocytosis of polystyrene microbeads by increasing the number of phagocytic cells as well as the number of phagocytosed beads per cell [24]. Intriguingly, groove depth, rather than width, was suggested to be the predominating factor influencing migration and phagocytosis. RAW264.7 cells cultured on nanogrooves (250 nm–2 μm in width) imprinted in polymers also display elongated morphologies (Fig. 1b). Macrophage response to various groove widths, in terms of both morphology change and cytokine secretion, appeared to be biphasic. Flat substrates and gratings less than 500 nm in width had little to no effect on macrophages, whereas cells cultured on 1 μm lines exhibit most elongated shape and show marked reduction in inflammatory cytokine secretion. Overall, the grooves appear to require longer durations to influence macrophage cytokine secretion profile, as the reduced secretion was only seen after 48 h [25]. In vivo studies using the same substrates reported similar findings in that microscale grooves reduce cell adhesion and foreign body giant cell formation when compared to planar substrates.
Alternative to surface patterned materials are electrospun polymer meshes, which more closely recapitulate native microenvironments. Saino et al. [26] investigated the activation of RAW264.7 cells on electrospun poly(L-lactic) (PLLA) fibrous materials (Fig. 1c). Both fiber diameter and alignment were found to affect macrophage adhesion and activation. Aligned fibers, of either micro- (~1.6 μm) and nano- (~600 nm) scale, appeared to enhance macrophage adhesion when compared to randomly aligned or flat PLLA substrates. In the presence of LPS, macrophages secreted less inflammatory cytokines on fibrous PLLA than on flat PLLA films. Unlike microgrooved substrates, the attenuation in activation caused by aligned fibers was only observed during the first 24 h, and not at the later 7 days time point. Similarly, electrospun polydioxanone (PDO) scaffolds containing larger fiber/pore size not only mitigated M1 activation but also skewed macrophage polarization toward an M2 phenotype, with increased expressions of arginase-1, VEGF, TGF-β1, and bFGF [27]. Implantation of random and aligned polycaprolactone (PCL) nanofibers in rats led to thinner fibrous capsules when compared to flat PCL films [28], suggesting reduced inflammation and enhanced wound healing around the fibrous materials. Consistent with these findings, Sanders et al. [29] examined a variety of different polymers and fiber sizes and demonstrated that in all materials, smaller fiber (1–5 μm diameter) resulted in thinner fibrous capsule formation when compared to larger fibers (11–15 μm diameter).
While the majority of the studies investigating the effect of substrate topology on macrophage function have been carried out on 2D engineered materials, it has been argued that cells can behave very differently in 3D microenvironments [30]. By embedding electrospun poly(lactic-co-glycolic acid) (PLGA) within a hydrogel, one group created a nanofibrous hydrogel system for investigating the effect of 2D versus 3D substrate architecture on human macrophages derived from peripheral blood mononuclear cells (PBMC). In accordance with previous studies, they report that 2D flat hydrogels stimulate secretion of inflammatory cytokines TNF-α and IL-1β. Interestingly, 3D nanofibrous hydrogels not only downregulate these inflammatory cytokines but also encourage production of pro-angiogenic chemokines IL-8 and MIP-1β [31]. This presents an important consideration for tissue engineering applications, and perhaps is also suggestive of a disease mechanism, as macrophages are known to promote angiogenesis in densely fibrillar tumor matrices [32]. 3D matrix architecture can also profoundly impact macrophage migration. While most leukocytes migrate via an amoeboid migration mode, macrophages employ both amoeboid and mesenchymal migration modes [33]. Using 3D fibrillar and gelled type I collagen, it has been demonstrated compellingly that matrix architecture dictates macrophage migration modes [34]. In gelled collagen, a dense matrix with few distinct fibrils, human monocyte-derived macrophages adopt the mesenchymal migration mode. On the other hand, in fibrillar collagen, macrophages migrate via the amoeboid mode. Remarkably, when presented with matrices of variable stiffness and architecture, macrophages adopt migration modes primarily based on architecture, thus establishing it as the dominant factor.
Stretch
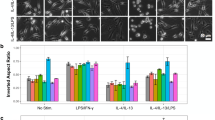
Certain tissues in the body, such as bones, vasculature and lungs, undergo substantial mechanical loading and stretch as part of their normal function. While physiological levels of mechanical stimulation are important for growth and maturation of cells in those tissues, abnormal stimulation has been associated with inflammation and disease [35, 36]. Macrophages present in these tissues are also exposed to these mechanical stimuli and have been known to respond to them (Fig. 2a, b). Osteoclasts, which belong to a special class of macrophages only found in bone tissues, increase their bone resorbing activities when exposed to cyclic stretch [37]. In both human alveolar and THP-1 monocyte derived macrophages, cyclic stretch alone induced secretion of IL-8. When applied concurrently with LPS stimulation, stretch enhanced LPS-mediated secretion of TNF-α, IL-6 and IL-83 [8]. Primary rat peritoneal macrophages subjected to static stretch increased expression of inflammatory genes including inducible nitric oxide synthase (iNOS), cyclooxygenase-2 (COX-2), IL-1β, IL-6, MIP-1α, and MIP-2 [39]. Stretch can also affect ECM integrity by upregulating macrophage production of matrix metalloproteinases such as MMP-1, MMP-3, MMP-9, and TIMP-1 [38, 40]. Cyclic stretch has also been shown to synergistically affect macrophage phenotypic polarization along with other biophysical cues. In human PBMCs seeded on electrospun elastomeric scaffold, a moderate 7 % cyclic strain was shown to increase the ratio of M2/M1 macrophages over a 7-day period when compared to unstretched control. However, a more extensive 12 % strain resulted in marked reduction in M2/M1 macrophages over the time course [41] (Fig. 2c). This biphasic effect perhaps reinforces the concept that an optimal or physiological level of stretch can promote tissue homeostasis, whereas abnormal stretch leads to inflammation.

a Mechanical stretch mediates macrophage morphology and activation [38, 51]. b Immunofluorescence images of control unstretched (top) and uniaxially stretched (bottom) human U937 macrophages stained for F-actin (red), vinculin (green), and nuclei (blue) [51]. c Immunofluorescent images of human peripheral blood mononuclear cell (PBMC) derived macrophages subjected to 0, 7 and 12 % strain on poly-ε-caprolactone (PCL) scaffold, and stained for M1 marker CCR7 (red), M2 marker CD163 (yellow), and dapi (blue) [41]
Substrate stiffness
In addition to applied forces, the stiffness of the surrounding tissue can also contribute to the mechanical environment of cells. The interstitial matrix surrounding cells can possess wide range of mechanical properties in vivo. In recent years, the rigidity of the underlying substrate has been shown to be critically important for cellular behaviors ranging from adhesion and contractility to migration [42] and differentiation. For instance, bone ECM features drastically different elastic modulus than neural tissues, and is an important differentiation cue for stem cells during development [43]. Tissue stiffness also changes dramatically during aging and the development of diseases. The progression of cancer and atherosclerosis are both accompanied by the stiffening of normal tissue. Using techniques such as the indentation test and atomic force microscopy to measure the elastic moduli of normal and diseased tissues, it has subsequently been shown that increase in ECM stiffness is critical for tumorigenesis [44] and atherogenesis [45]. In addition to its contribution to disease progression, stiffness also plays a role in the host response to implanted biomaterials as most synthetic materials have very different elastic moduli than native tissue.
Considering the extensive role of macrophages in diseases and mediating tissue-biomaterial interactions, there have been numerous investigations examining the effect of substrate stiffness on macrophage phenotype. Using Arg-Gly-Asp (RGD)-functionalized poly(ethylene glycol) (PEG) hydrogels, Blakney et al. [46] tuned the compressive moduli of the substrate without changing the amount of cell adhesion ligands. Although it has been previously shown that RAW264.7 cells seeded onto PEG hydrogel with gradient elasticity preferentially adhere to the stiffer parts of the hydrogel [47], this group found no significant difference in adhesion on substrate stiffness ranging from 130 to 840 kPa. They then examined the effect of stiffness on phenotypic polarization of murine bone marrow derived macrophages (BMDMs), and found no significant differences in the absence of soluble co-stimulants. However, in the presence of LPS, the expression of TNF-α, IL-6, IL-1β and IL-10 all increased with increasing rigidity. In accordance with these in vitro results, subcutaneous implantation of stiff hydrogels in mice elicited a more severe foreign body response, evidenced by the recruitment of more inflammatory macrophages, than soft hydrogels.
In studies using human macrophages, the elastic modulus of the substrate and surface adhesion ligand density have also been shown to affect adhesion and cytokine secretion. Macrophages differentiated from human THP-1 cells were examined on PEG-coated polyacrylamide gels of stiffnesses ranging from 1.4 to 348 kPa. Without any RGD adhesion peptide modification, human macrophages preferentially adhered to stiffer substrates when compared to soft substrates. On RGD peptide-modified surfaces, the same trend of more adhesion on the stiffer substrates remained, although the differences were less pronounced. Interestingly, TNF-α secretion by adherent human macrophages was found to be inversely proportional to the substrate stiffness: cells secreted most TNF-α on 1.4 kPa substrate and least on 348 kPa. It is important to note that this result was obtained by normalizing the bulk population secretion to the number of adherent cells, and without normalization, there was no difference in TNF-α secretion. In contrast to TNF-α, the IL-8 secretion profile appeared to be biphasic in that secretion was low on 1.4 and 348 kPa but high on the intermediate 10 kPa substrate [48].
In a separate study, murine and differentiated human macrophages were investigated side by side on polyacrylamide gels with elastic moduli ranging from 0.3 to 76.8 kPa. Cell adhesion over this stiffness range was not significantly different. Without LPS activation, substrate rigidity appears to have little effect on TNF-α secretion in both RAW264.7 and human U937 macrophages. However, with LPS stimulation, TNF-α production inversely correlated with rigidity in both cell types. Increasing stiffness also enhanced opsonization-mediated macrophage phagocytosis in both RAW264.7 and human alveolar macrophages (AM) [49].
The effect of substrate rigidity on macrophage adhesion and activation may still be up for debate. Experimentally, the effect of stiffness on macrophage adhesion and cytokine secretion need to be clearly demarcated, as secretion profiles for adherent and non-adherent macrophages are likely different. Similar to stretch, there may be one or more optimal range of stiffness that is ideal for homeostasis. Moreover, various tissue-specific macrophages are accustomed to ECMs of different elasticity in vivo, and therefore stiffness-dependent effects on macrophage phenotype may be ontogenetic.
Cell shape
Cells change their morphologies as they probe and migrate through their surrounding matrix, and alterations in cell shape have long been associated with changes in cell function [50]. Moreover, many of the aforementioned physical cues, including topography, stretch, and stiffness, profoundly influence cell shape. In studies of macrophages, cyclic biaxial stretch increased the spread cell area [38], whereas uniaxial stretch led to cellular elongation along the direction of stretch [51] (Fig. 2a, b). Multiple studies investigating the effect of substrate stiffness have shown that macrophages are more spread and flattened on rigid substrates, and more rounded on soft [46, 48, 52]. Furthermore, stiffer substrates tend to encourage the formation of long protrusive actin structures such as filapodia [49, 52]. Polymers imprinted with micro- and nano- scale grooves are capable of directing macrophage cells to exhibit elongated morphologies both in vitro and in vivo [24, 25].
Macrophages also undergo morphological changes in response to soluble factors. A number of groups have observed that when murine BMDMs are stimulated with LPS and IFN-γ, they adopt a circular and flatten morphology. However, in the presence of M2-inducing cytokines IL-4 and IL-13, they exhibit much more elongated morphologies [53, 54]. The activation of microglial cells, which are the resident macrophages of the central nervous system, occurs in concert with distinct morphological changes [55]. It was subsequently shown that soluble factors secreted by adipose tissue-derived mesenchymal stem cells (ASCs) lead to elongated cell shape in microglia, which is also accompanied by a noninflammatory phenotype characterized by low TNF-α and IL-6, and high arginase-I production [56] (Fig. 3b).

a Cell elongation, induced by adhesive or soluble cues, modulates macrophages phenotype [53, 56]. b Immunofluorescence images of CD11b expression in microglia cells exposed to medium conditioned (CM) by mesenchymal stem cells and/or LPS (top), and their secretion of IL-6 and TNF-α, and expression of arginase-1 (bottom). Scale bar 10 μm [56]. c Phase contrast images of BMDMs on unpatterned and patterned surfaces (left), and expression of iNOS and arginase-1 in these conditions (right). Scale bar 50 μm [53]
The development of various microfabrication and bioengineering tools in recent years has enabled unprecedented control over cell geometry, either singularly or in groups, to specifically study the effect of cell shape on function. Using these technologies, cell shape has been demonstrated to be an important regulator of growth and apoptosis in endothelial cells [57], differentiation and multipotency in mesenchymal stem cells [58, 59], collagen synthesis in fibroblasts [60], and proliferation [61] and contractility [62] in vascular smooth muscle cells. Using a micropatterning approach to directly control cell shape, we recently demonstrated that elongated cell morphology alone could promote BMDM polarization toward an M2 phenotype, with increased expression of arginase-I, CD206 and YM-1 (Fig. 3c). Elongated cell shape can also modulate soluble factor mediated polarization by either upregulating M2 phenotype marker expression or mitigating M1 activation. Furthermore, when cellular elongation was attenuated, IL-4 and IL-13 stimulated cells produced less arginase-I, implying impaired M2 polarization [53]. Cell shape has also been linked to polarization state in human macrophages. In one study examining the effects of stretch and electrospun scaffold on PBMC-derived macrophages, it was observed that CCR7+ M1 macrophages were small and rounded, whereas CD206+ M2 cells exhibited more actin protusions [41]. Human THP-1 derived macrophages cultured on substrates such as glass, polyurethane, modified chitosan and hyaluronic acid (HA) have been shown to display distinctly different morphologies in 3D. Cells on glass are most spread out, and possess the largest total surface area and volume. Interestingly, these cells also secreted the most TNF-α. Cells on chitosan and HA were nearly spherical and have the smallest surface area and volume, which correlated with low TNF-α secretion. Finally, cells on polyurethane were less spread than glass, and present intermediate surface area, volume as well as TNF-α secretion [63].
Although it is evident that morphology is strongly linked to phenotype in certain macrophages, the phenomenon may not be universal. We, and others, have observed that while M2-polarized BMDMs and microglia display cellular elongation, other macrophages do not exhibit such dramatic changes. Nevertheless, morphological changes are inevitably rooted in cytoskeletal reorganization and regulated by underlying signaling pathways, which may suggest a mechanotransductive pathway that extends to all macrophages.
Intracellular mechanics
3D-reconstructed confocal stacks of phalloidin-stained rat AMs show that the majority of the macrophage actin cytoskeleton are cortical and form a thick submembranous stratum [52], as opposed to organized stress fibers observed in many adherent cells. Stiff substrates have been shown to increase actin polymerization and filopodial projections [46, 49, 52]. Murine macrophages plated on microgrooved substrates exhibit higher overall F-actin content and also along the edges of the microridges [23]. Several studies have noted differences in actin organization between M1 and M2-polarized macrophages [53, 64] (Fig. 4A). M1 cells display more clustered F-actin structures that are more likely to be found around the nuclei, whereas M2 cells present more diffused actin organization that are perhaps more around the periphery of the cells, suggestive of lamellipodia formation.

a Fluorescence images of F-actin of BMDMs on 20 μm-wide patterned fibronectin lines and unpatterned cells that are stimulated with IL-4/IL-13 or LPS/IFN-ϒ. Scale bar 50 μm [53]. b Immunofluorescence images of wt, Rac1−/−, Rac2−/− and Rac1/2−/− BMDMs stained with F-actin (red) and paxillin (green). Inserts are higher magnification images of podosomes. Scale bar 10 μm [54]. c Schematic of macrophage mechanotransduction pathway. Podosomes, which contain focal adhesion proteins, mediate macrophage adhesion and links the actin cytoskeleton to the ECM
A few studies have implicated intrinsic changes in stiffness and elasticity of macrophage cells as a possible mechanism for sensing changes in the physical microenvironment [49, 52]. Rat AMs cultured on stiff substrates exhibit increased cellular elastic modulus. Surprisingly, this phenomenon cannot be attributed to an increase in actin polymerization, as treatment with inhibitor cytochalasin D failed to abolish the difference in cellular stiffness due to substrate rigidity. Using magnetic twisting cytometry (MTC), AMs were shown to have minimum internal tension. Although not detectable by MTC, it is possible for macrophages to generate local stress points that co-localize with punctated podosomes and actin structures, especially during actin-mediated tasks such as migration and phagocytosis [52]. This seems to be consistent with the general understanding that podosome assembly does not require intracellular tension but rather depend on extracellular cues such as substrate flexibility [65]. These results, together with the lack of observable actin stress fibers (Fig. 4a, b), allude to a macrophage mechanotransduction mechanism that may be distinct from most other tissue cells.
Evidence strongly suggests that macrophage elasticity is linked to its activation and function. Using colloidal force microscopy, LPS-activated human macrophages were shown have an average Young’s modulus that is three times less than that of unstimulated resting macrophages. The same study also found that stimulation with LPS stimulation caused an increase in adhesion strength of macrophages [66]. In murine RAW264.7 cells, LPS and IFN-γ stimulation increased cellular elasticity, unlike in human macrophages. However, LPS increased macrophage adhesion strength, as was also shown with human cells [49, 67]. A more rigid substrate increases elasticity, similar to stimulation with LPS and IFN-γ, and this increased elasticity corroborates with enhanced phagocytosis and reactive oxygen species (ROS) production. However, contrary to the study in rat AMs [68], this particular study demonstrated that actin polymerization was required for murine macrophage elasticity while actin-myosin contractility has minimal involvement [49]. It has been previously suggested that cellular stiffness is regulated by both cell shape and substrate rigidity [69]. Although the exact mechanics remain elusive, there is emerging evidence supporting the idea that biophysical regulation of macrophage behavior is mediated by alterations in cell stiffness and shape.
Molecular mechanism for mechanotransduction in macrophages
For macrophages to be mechanically activated, they must be able to probe and discern changes in the mechanical properties of their environment and transduce these changes into biochemical signals that ultimately lead to differential gene and protein expressions. To date, most studies in mechanotransduction have been performed on low motile tissue cells with highly polymerized actin organization. In contrast, macrophages are derived from nonadherent monocytes, and once activated become adherent and migrate through tissue to reach the site of injury or inflammation. Macrophage adhesion to the extracellular matrix environment is mediated by various types of integrins, as well as scavenger receptors. Interestingly, macrophages have been known to deploy different modes of adhesion to type collagen I that may be dependent on species, cell source, or the context of adhesion. Generally, β 1 integrin is thought to be the main receptor for macrophage adhesion to collagen [70]. In rat alveolar macrophages (AMs), CD18, the β 2 subunit of integrin, was shown to be main regulator of adhesion to type I collagen, whereas CD11b, the α M subunit, did not participate in adhesion [52]. In RAW264.7 macrophages, neither β 1 nor β 2 subunits are involved in adhesion to denatured type I collagen. Instead, it was largely modulated by class A macrophage scavenger receptor (SR-A) [71]. SR-A has also been implicated in mediating macrophage adhesion to a number of other modified ECM components, including glycated [72], proteoglycan- [73] and cigarette smoke- [74] modified ECM, suggesting a possible disease mechanism in which modified ECMs associated with disease progression may enhance macrophage adhesion through SR-A.
Once adherent, membrane adhesion receptors of most cells cluster and recruit additional proteins to form focal adhesions (FA), which are linked to the cytoskeleton and provide a signal transduction pathway for physical cues from extracellular space. In place of FAs, macrophages form punctated focal complexes known as podosomes, which are similar to FA in protein composition but different in structure and dynamics. Podosomes are cylindrical structures that contain an actin-rich core and form perpendicular to the membrane rather than tangential (Fig. 4a). They are more dynamic and unstable than FAs, which may play a key role in macrophage migration, invasion and degradation of surrounding matrices [65]. The distribution, size and shape of these focal complexes vary and are a reflection of changes in macrophage adhesion and morphology. The peripheries of podosomes are composed of integrins, the specific types of which are tissue dependent [75, 76]. The cores of podosomes also contain CD44, a protein that can modulate macrophage adhesion through its affinity to hyaluronan, collagen and matrix metalloproteases. Adherent, well-spread macrophage cells generally develop larger podosomes [77], and the traction force generated by podosomes are sensitive to substrate rigidity, which is the reason that they are often considered mechanosensors [78]. Studies on macrophage podosome dynamics in 3D collagen gels recapitulate results from 2D, suggesting similar macrophage mechanosensing mechanisms in 2D and 3D [79].
Adhesion can also lead to integrin clustering and subsequently activate focal adhesion kinase (FAK) [80, 81]. Downstream of that are the Rho family of GTPases, which include RhoA, Rac, and cdc42. These signaling proteins, generally activated following FAK phosphorylation, are the key regulators of actin cytoskeletal dynamics and are directly involved in the assembly of actin stress fibers (RhoA), lamellipodia (Rac), and filipodia (cdc42) [82]. The importance of these proteins in mechanotransduction has been well documented in many cell types, and they have been implicated in mediating adhesion, spreading, phagocytosis, and podosome assembly in macrophages [83–86]. More recently, several studies have highlighted the significance of Rac in modulating macrophage morphology, migration and phenotype polarization. Among the three highly conserved isoforms of Rac, Rac1 is ubiquitous in all mammalian cells including macrophages, whereas Rac2 is only found in hematopoetic and endothelial cells. Deletion of Rac1 and Rac2 in mice has been shown to alter macrophage cell shape. Comparing to wildtype, Rac1−/− BMDMs display elongated morphologies and reduced spread area. Rac2−/− cells also exhibited an elongated cell shape, though to a lesser extent than Rac1−/− cells, but did not show difference in cell spreading. Rac2 knockout also completely abolished podosome formation in BMDMs, whereas Rac1 knockout only led to abnormal podosome assembly (Fig. 4b). Surprisingly, abnormal podosomes due to Rac1 deletion did not affect macrophage migratory abilities. Rac2 deletion reduced the number of migratory cells but did not affect their migration speed [54]. In mouse tumor models, Rac2 was implicated in mediating macrophage migration into tumors via integrin α 4 β 1. It was subsequently shown that Rac2 activation regulates macrophage polarization to tumor-associated M2 phenotype both in vitro and in vivo [87].
Taken together, these studies overwhelmingly support the notion that macrophages respond to changes in the physical microenvironment, although different activation states may result in differential sensitivity to mechanical cues. LPS or M1-like activation has been demonstrated to enhance cell adhesion in both human and murine macrophages [49, 64, 67]. Macrophage-1 antigen (Mac1), an integrin family receptor composed of both CD11b and CD18, is well known for mediating phagocytosis [80] and is upregulated in murine macrophages stimulated with LPS and IFN-γ. Additionally, these cells upregulate adhesion receptors lymphocyte function associated antigen-1 (LFA-1), CD29, and CD11a, which may partially explain their enhanced adhesion [64]. As adhesion proteins are a major component of macrophage membrane mechanosensor podosomes, it is not surprising that podosome assembly can also be modulated by phenotypic states. The cytokine CSF-1, which is known to steer macrophage polarization toward an M2-like phenotype [4], has been shown to increase the assembly and alter the distribution of podosomes in BMDMs [88]. IL-4—polarized human macrophages form podosome rosettes, which are not seen in IFNγ-induced M1 or unstimulated resident macrophages. The formation of these rossettes is linked to increased degradation and migration through the ECM by M2 macrophages, and may also explain the ability of M2 cells to adopt a mesenchymal migration mode [89]. Murine M1 macrophages also exhibit increased cellular elastic modulus, which correlates with their enhanced ability to uptake tissue fragments [64] and opsonized particles [49, 67]. In addition to phagocytosis, intrinsic cell stiffness can affect other cytoskeleton-mediated macrophage behavior such as migration. It is thought that migration speed is inversely correlated with intracellular tension [65], and consistent with this, M2 primary murine macrophages are seen as more motile than M1 cells in the absence of chemoattractant [64]. Alterations in cellular elasticity and podosome assembly, underlied by differential activation of RhoA and Rac, may also explain the different macrophages migration modes, as 3D matrix architecture and stiffness are thought to modulate mesenchymal versus amoeboid modes of migration by macrophages [34]. While the underlying molecular mechanism for mechanically induced macrophage phenotype and function remains to be determined, various studies have implicated molecules including integrins [80, 81], FAK-src activation [22, 77], actin polymerization [49, 53, 68], actin-myosin contractility [53, 78], and Rho GTPases RhoA [34, 64], Rac [34, 54, 64, 87, 90] and cdc42 [49] (Fig. 4c).
Concluding remarks and future directions
Recent studies clearly suggest that physical cues in the microenvironment can modulate macrophage phenotype and function. In addition to affecting change on their own, biophysical cues can also function synergistically with soluble factors to mediate macrophage behavior. Although much of the work highlighted here is performed on engineered substrates, with work in 3D beginning to emerge, the effects observed may very well translate to in vivo conditions. Tissues and various components of tissue can be conceptualized as distinct biomaterials that can possess wide range of mechanical properties and exert physical influences on macrophages in the same way engineered materials can. While soluble factors are the putative reason for abnormal macrophage polarization in diseases, physical and mechanical factors associated with ECM modification may exacerbate disease progression. Experimental approaches by means of selectively targeting macrophages with disease phenotypes or introducing beneficial ones have so far been unsuccessful [102, 103]. Evidence suggests that most likely, more than one macrophage phenotype is required for complex processes such as wound healing. The key may lie in the timely transition from one phenotype to another. Leveraging the functional plasticity of macrophages, by both biochemical and physical means, to “re-educate” them to attain beneficial phenotypes in a timely manner may prove to be a promising therapeutic strategy.
However, despite the general consensus that mechanical factors are important, there are few seemingly contradictory findings in the current body of literature. Likely contributing to this confusion are several factors: (1) There are differences between macrophages from different species. The majority of studies described in this review were performed using murine and human macrophages, with a few using rat cells. While experiments with murine systems have been extremely valuable particularly for molecular-genetic studies, and mouse and human immune systems clearly share some similarities, there are also fundamental differences [91]. Although macrophages from both species adopt a similar continuum of activation states, it has been well established that the markers for each state can be quite different [92, 93]. A cross-species comparison for macrophage activation in response to physical and mechanical stimulation is still lacking. (2) Macrophages from different tissues within the same host may have different sensitivity to mechanical cues. For example, alveolar macrophages and osteoclasts naturally undergo more mechanical loading in the body, and therefore may be more sensitive to physical stimuli. In addition, ECMs from different tissues can have dramatically different stiffness and architecture, which may result in differential responsiveness to physical signals by macrophages in those tissues. (3) The length and time scale at which the effects of mechanical cues are examined is important. It merits reiteration that there is likely a range of ECM stiffness and architecture that best supports tissue homeostasis and physiological macrophage function. Engineered materials should closely mimic the cellular niche of the intended targets of study. Additionally, macrophage secretion profile of different cytokines is likely to vary over time. Several studies have shown a temporal dependence of the effect of mechanical cues, especially substrate topography and rigidity. (4) Macrophages are very sensitive to various soluble and matrix cues, and possibly to different materials including the polymers and metals that are used as biomaterials. It has been demonstrated that adhesion to various ECM proteins can induce macrophage activation [94, 95] and enhance phagocytosis [96]. Therefore, future studies should be assiduous in extricating the effects of matrix and mechanics. (5) Last but not least, immune cells including macrophages are notorious for being very heterogeneous. The ground state activation level of macrophages is known to fluctuate depending on time of the day, seasons of the year, diet and systemic conditions of the host [97], which may explain inconsistencies in studies using primary cells.
Clearly, much work is still needed in elucidating the underlying mechanism for macrophage mechanobiology. The link between adhesion, cytoskeletal dynamics and activation remains ambiguous at best. The pathway between the cytoskeleton, nuclei and possible transcription factors is largely unknown. Topographical studies have shown that surface roughness alone can cause src-mediated FAK phosphorylation [77], which then leads to nuclear translocation of ERK1/2 and increased secretion of proinflammatory cytokines in RAW264.7 cells [20]. It has also been postulated that substrate topography can activate the NF-κB-mediated inflammatory signaling pathway via cell surface receptors found in actin-associated, lipid-enriched rafts on the cell membrane [98]. Stretch-induced inflammation in macrophages has been attributed to NF-κB [38] and NLRP3 inflammasome [99] activation. Stretch-induced MMP expressions have been linked to upregulation in transcription factors c-jun, c-fos and PU.1 [40]. There has also been studies suggesting that mechanically sensitive potassium channels are responsible for modulating macrophage cell shape upon adherence as well as stretch-mediated activation [100]. Alternatively, macrophage activation can be regulated by epigenetics [101], and it has also been postulated that the physical and mechanical activation may be due to post-translational modifications that are not associated with gene expression, such as ligand-receptor interactions and cell membrane extensions [49].
References
Wynn TA, Chawla A, Pollard JW (2013) Macrophage biology in development, homeostasis and disease. Nature 496:445–455
Sieweke MH, Allen JE (2013) Beyond stem cells: self-renewal of differentiated macrophages. Science 342:1242974
Guilliams M et al (2014) Dendritic cells, monocytes and macrophages: a unified nomenclature based on ontogeny. Nat Rev Immunol 14:1–8. doi:10.1038/nri3712
Murray PJ et al (2014) Macrophage activation and polarization: nomenclature and experimental guidelines. Immunity 41:14–20
Mosser DM, Edwards JP (2008) Exploring the full spectrum of macrophage activation. Nat Rev Immunol 8:958–969
Gautier EL et al (2012) Gene-expression profiles and transcriptional regulatory pathways that underlie the identity and diversity of mouse tissue macrophages. Nat Immunol 13:1118–1128
Mantovani A et al (2004) The chemokine system in diverse forms of macrophage activation and polarization. Trends Immunol 25:677–686
Jetten N et al (2014) Anti-inflammatory M2, but not pro-inflammatory M1 macrophages promote angiogenesis in vivo. Angiogenesis 17:109–118
Lumeng CN, DelProposto JB, Westcott DJ, Saltiel AR (2008) Phenotypic switching of adipose tissue macrophages with obesity is generated by spatiotemporal differences in macrophage subtypes. Diabetes 57:3239–3246
Biswas SK, Mantovani A (2010) Macrophage plasticity and interaction with lymphocyte subsets: cancer as a paradigm. Nat Immunol 11:889–896
Fujiu K, Wang J, Nagai R (2014) Cardioprotective function of cardiac macrophages. Cardiovasc Res 102:232–239
Van Amerongen MJ, Harmsen MC, van Rooijen N, Petersen AH, van Luyn MJA (2007) Macrophage depletion impairs wound healing and increases left ventricular remodeling after myocardial injury in mice. Am J Pathol 170:818–829
Lewis CE, Pollard JW (2006) Distinct role of macrophages in different tumor microenvironments. Cancer Res 66:605–612
Cornelissen R et al (2014) Ratio of intratumoral macrophage phenotypes is a prognostic factor in epithelioid malignant pleural mesothelioma. PLoS One 9:e106742
Novak ML, Koh TJ (2013) Phenotypic transitions of macrophages orchestrate tissue repair. Am J Pathol 183:1352–1363
Sica A et al (2008) Macrophage polarization in tumour progression. Semin Cancer Biol 18:349–355
Brown BN, Valentin JE, Stewart-Akers AM, McCabe GP, Badylak SF (2009) Macrophage phenotype and remodeling outcomes in response to biologic scaffolds with and without a cellular component. Biomaterials 30:1482–1491
Brown BN et al (2012) Macrophage phenotype as a predictor of constructive remodeling following the implantation of biologically derived surgical mesh materials. Acta Biomater 8:978–987
Rich A, Harris AK (1981) Anomalous preferences of cultured macrophages for hydrophobic and roughened substrata. J Cell Sci 50:1–7
Refai AK, Textor M, Brunette DM, Waterfield JD (2004) Effect of titanium surface topography on macrophage activation and secretion of proinflammatory cytokines and chemokines. J Biomed Mater Res Part A 70:194–205
Tan KS, Qian L, Rosado R, Flood PM, Cooper LF (2006) The role of titanium surface topography on J774A.1 macrophage inflammatory cytokines and nitric oxide production. Biomaterials 27:5170–5177
Lee S et al (2011) Analysis on migration and activation of live macrophages on transparent flat and nanostructured titanium. Acta Biomater 7:2337–2344
Wójciakâ-Stothard B, Madeja Z, Korohoda W, Curtis A, Wilkinson C (1995) Activation of macrophage-like cells by multiple grooved substrata. Topographical control of cell behaviour. Cell Biol Int 19:485
Wójciak-Stothard B, Curtis A, Monaghan W, MacDonald K, Wilkinson C (1996) Guidance and activation of murine macrophages by nanometric scale topography. Exp Cell Res 223:426–435
Chen S et al (2010) Characterization of topographical effects on macrophage behavior in a foreign body response model. Biomaterials 31:3479–3491
Saino E et al (2011) Effect of electrospun fiber diameter and alignment on macrophage activation and secretion of proinflammatory cytokines and chemokines. Biomacromolecules 12:1900–1911
Garg K, Pullen NA, Oskeritzian CA, Ryan JJ, Bowlin GL (2013) Macrophage functional polarization (M1/M2) in response to varying fiber and pore dimensions of electrospun scaffolds. Biomaterials 34:4439–4451
Cao H, McHugh K, Chew SY, Anderson JM (2010) The topographical effect of electrospun nanofibrous scaffolds on the in vivo and in vitro foreign body reaction. J Biomed Mater Res A 93:1151–1159
Sanders JE, Bale SD, Neumann T (2002) Tissue response to microfibers of different polymers: polyester, polyethylene, polylactic acid, and polyurethane. J Biomed Mater Res 62:222–227
Baker BM, Chen CS (2012) Deconstructing the third dimension - how 3D culture microenvironments alter cellular cues. J Cell Sci 125:3015–3024
Bartneck M et al (2012) Inducing healing-like human primary macrophage phenotypes by 3D hydrogel coated nanofibres. Biomaterials 33:4136–4146
Brown E et al (2003) Dynamic imaging of collagen and its modulation in tumors in vivo using second-harmonic generation. Nat Med 9:796–800
Guiet R et al (2011) The process of macrophage migration promotes matrix metalloproteinase-independent invasion by tumor cells. J Immunol 187:3806–3814
Van Goethem E, Poincloux R, Gauffre F, Maridonneau-Parini I, Le Cabec V (2010) Matrix architecture dictates three-dimensional migration modes of human macrophages: differential involvement of proteases and podosome-like structures. J Immunol 184:1049–1061
Liu M, Post M (2000) Invited review: mechanochemical signal transduction in the fetal lung. J Appl Physiol 89:2078–2084
Klein-Nulend J, Bacabac RG, Mullender MG (2005) Mechanobiology of bone tissue. Pathol Biol (Paris) 53:576–580
Kurata K et al (2001) Mechanical strain effect on bone-resorbing activity and messenger RNA expressions of marker enzymes in isolated osteoclast culture. J Bone Miner Res 16:722–730
Pugin J et al (1998) Activation of human macrophages by mechanical ventilation in vitro. Am J Physiol 275:L1040–L1050
Wehner S et al (2010) Mechanical strain and TLR4 synergistically induce cell-specific inflammatory gene expression in intestinal smooth muscle cells and peritoneal macrophages. Am J Physiol Gastrointest Liver Physiol 299:G1187–G1197
Yang JH, Sakamoto H, Xu EC, Lee RT (2000) Biomechanical regulation of human monocyte/macrophage molecular function. Am J Pathol 156:1797–1804
Ballotta V, Driessen-Mol A, Bouten CVC, Baaijens FPT (2014) Strain-dependent modulation of macrophage polarization within scaffolds. Biomaterials 35:4919–4928
Lo CM, Wang HB, Dembo M, Wang YL (2000) Cell movement is guided by the rigidity of the substrate. Biophys J 79:144–152
Engler AJ, Sen S, Sweeney HL, Discher DE (2006) Matrix elasticity directs stem cell lineage specification. Cell 126:677–689
Paszek MJ et al (2005) Tensional homeostasis and the malignant phenotype. Cancer Cell 8:241–254
Huynh J et al (2011) Age-related intimal stiffening enhances endothelial permeability and leukocyte transmigration. Sci Transl Med 3:112ra122
Blakney AK, Swartzlander MD, Bryant SJ (2012) The effects of substrate stiffness on the in vitro activation of macrophages and in vivo host response to poly(ethylene glycol)-based hydrogels. J Biomed Mater Res A 100:1375–1386
Nemir S, Hayenga HN, West JL (2010) PEGDA hydrogels with patterned elasticity: novel tools for the study of cell response to substrate rigidity. Biotechnol Bioeng 105:636–644
Irwin EF et al (2008) Modulus-dependent macrophage adhesion and behavior. J Biomater Sci Polym Ed 19:1363–1382
Patel NR et al (2012) Cell elasticity determines macrophage function. PLoS One 7:e41024
Folkman J, Moscona A (1978) Role of cell shape in growth control. Nature 273:345–349
Matheson LA, Fairbank NJ, Maksym GN (2006) Characterization of the Flexcell Uniflex cyclic strain culture system with U937 macrophage-like cells. Biomaterials 27:226–233
Féréol S et al (2006) Sensitivity of alveolar macrophages to substrate mechanical and adhesive properties. Cell Motil Cytoskeleton 63:321–340
McWhorter FY, Wang T, Nguyen P, Chung T, Liu WF (2013) Modulation of macrophage phenotype by cell shape. Proc Natl Acad Sci USA 110:17253–17258
Wheeler AP et al (2006) Rac1 and Rac2 regulate macrophage morphology but are not essential for migration. J Cell Sci 119:2749–2757
Hanisch U-K, Kettenmann H (2007) Microglia: active sensor and versatile effector cells in the normal and pathologic brain. Nat Neurosci 10:1387–1394
Neubrand VE et al (2014) Mesenchymal stem cells induce the ramification of microglia via the small RhoGTPases Cdc42 and Rac1. Glia 62(12):1932–1942. doi:10.1002/glia.22714
Chen CS (1997) Geometric control of cell life and death. Science 276:1425–1428
Yang Y, Relan NK, Przywara DA, Schuger L (1999) Embryonic mesenchymal cells share the potential for smooth muscle differentiation: myogenesis is controlled by the cell’s shape. Dev Cambridge Engl 126:3027–3033
Kilian KA, Bugarija B, Lahn BT, Mrksich M (2010) Geometric cues for directing the differentiation of mesenchymal stem cells. Proc Natl Acad Sci USA 107:4872–4877
Li F, Li B, Wang Q-M, Wang JH-C (2008) Cell shape regulates collagen type I expression in human tendon fibroblasts. Cell Motil Cytoskeleton 65:332–341
Thakar RG et al (2009) Cell-shape regulation of smooth muscle cell proliferation. Biophys J 96:3423–3432
Alford PW, Nesmith AP, Seywerd JN, Grosberg A, Parker KK (2011) Vascular smooth muscle contractility depends on cell shape. Integr Biol (Camb) 3:1063–1070
Lee H-S et al (2013) Correlating macrophage morphology and cytokine production resulting from biomaterial contact. J Biomed Mater Res A 101:203–212
Vereyken EJF et al (2011) Classically and alternatively activated bone marrow derived macrophages differ in cytoskeletal functions and migration towards specific CNS cell types. J Neuroinflammation 8:58
Block MR et al (2008) Podosome-type adhesions and focal adhesions, so alike yet so different. Eur J Cell Biol 87:491–506
Leporatti S et al (2006) Elasticity and adhesion of resting and lipopolysaccharide-stimulated macrophages. FEBS Lett 580:450–454
Pi J (2014) Detection of lipopolysaccharide induced inflammatory responses in RAW264.7 macrophages using atomic force microscope. Micron 65:1–9. doi:10.1016/j.micron.2014.03.012
Féréol S et al (2008) Mechanical and structural assessment of cortical and deep cytoskeleton reveals substrate-dependent alveolar macrophage remodeling. Biomed Mater Eng 18:S105–S118
Tee SY, Fu J, Chen CS, Janmey PA (2011) Cell shape and substrate rigidity both regulate cell stiffness. Biophys J 100
Friedl P, Weigelin B (2008) Interstitial leukocyte migration and immune function. Nat Immunol 9:960–969
Gowen BB, Borg TK (2000) Ghaffar, a & Mayer, E. P. Selective adhesion of macrophages to denatured forms of type I collagen is mediated by scavenger receptors. Matrix Biol 19:61–71
El Khoury J et al (1994) Macrophages adhere to glucose-modified basement membrane collagen IV via their scavenger receptors. J Biol Chem 269:10197–10200
Santiago-García J, Kodama T, Pitas RE (2003) The class A scavenger receptor binds to proteoglycans and mediates adhesion of macrophages to the extracellular matrix. J Biol Chem 278:6942–6946
Kirkham PA, Spooner G, Ffoulkes-Jones C, Calvez R (2003) Cigarette smoke triggers macrophage adhesion and activation: role of lipid peroxidation products and scavenger receptor. Free Radic Biol Med 35:697–710
Duong LT, Rodan GA (2000) Pyk2 is an adhesion kinase in macrophages, localized in podosomes and activated by B2-integrin ligation. Cell Motil Cytoskeleton 47:174–188
Pfaff M, Jurdic P (2001) Podosomes in osteoclast-like cells: structural analysis and cooperative roles of paxillin, proline-rich tyrosine kinase 2 (Pyk2) and integrin alphaVbeta3. J Cell Sci 114:2775–2786
Ghrebi S, Hamilton DW, Douglas Waterfield J, Brunette DM (2013) The effect of surface topography on cell shape and early ERK1/2 signaling in macrophages; linkage with FAK and Src. J Biomed Mater Res A 101:2118–2128
Collin O et al (2008) Self-organized podosomes are dynamic mechanosensors. Curr Biol 18:1288–1294
Van Goethem E et al (2011) Macrophage podosomes go 3D. Eur J Cell Biol 90:224–236
Berton G, Lowell C (1999) Integrin signalling in neutrophils and macrophages. Cell Signal 11:621–635
Zaveri TD, Lewis JS, Dolgova NV, Clare-Salzler MJ, Keselowsky BG (2014) Integrin-directed modulation of macrophage responses to biomaterials. Biomaterials 35:3504–3515
Hall A (1998) Rho GTPases and the actin cytoskeleton. Science 279:509–514
Aepfelbacher M, Essler M, Huber E, Czech A, Weber PC (1996) Rho is a negative regulator of human monocyte spreading. J Immunol 157:5070–5075
Allen WE, Jones GE, Pollard JW, Ridley AJ (1997) Rho, Rac and Cdc42 regulate actin organization and cell adhesion in macrophages. J Cell Sci 110(Pt 6):707–720
Ory S, Brazier H, Pawlak G, Blangy A (2008) Rho GTPases in osteoclasts: orchestrators of podosome arrangement. Eur J Cell Biol 87:469–477
Hoppe AD, Swanson JA (2004) Cdc42, Rac1, and Rac2 display distinct patterns of activation during phagocytosis. Mol Biol Cell 15:3509–3519
Joshi S, Singh A, Zulcic M, Bao L, Messer K (2014) Rac2 controls tumor growth, metastasis and M1-M2 macrophage differentiation in vivo. PLoS One 9(4):e95893
Wheeler AR, Smith SD, Ridley AJ (2006) CSF-1 and PI 3-kinase regulate podosome distribution and assembly in macrophages. Cell Motil Cytoskeleton 63:132–140
Cougoule C et al (2012) Blood leukocytes and macrophages of various phenotypes have distinct abilities to form podosomes and to migrate in 3D environments. Eur J Cell Biol 91:938–949
Pradip D, Peng X, Durden DL (2003) Rac2 specificity in macrophage integrin signaling: potential role for Syk kinase. J Biol Chem 278:41661–41669
Mestas J, Hughes CCW (2004) Of mice and not men: differences between mouse and human immunology. J Immunol 172:2731–2738
Raes G et al (2002) Differential expression of FIZZ1 and Ym1 in alternatively versus classically activated macrophages. J Leukoc Biol 71:597–602
Raes G et al (2005) Arginase-1 and Ym1 are markers for murine, but not human, alternatively activated myeloid cells. J Immunol 174:6561–6562
Okamura Y et al (2001) The extra domain A of fibronectin activates Toll-like receptor 4. J Biol Chem 276:10229–10233
Smiley ST, King JA, Hancock WW (2001) Fibrinogen stimulates macrophage chemokine secretion through toll-like receptor 4. J Immunol 167:2887–2894
Newman SL, Tucci MA (1990) Regulation of human monocyte/macrophage function by extracellular matrix by activation of complement receptors and enhancement of fc receptor function monocyte/mrb bactericidal assay. J Clin Invest 86:703–714
Huang E, Wells CA (2014) The ground state of innate immune responsiveness is determined at the interface of genetic, epigenetic, and environmental influences. J Immunol 193:13–19
Waterfield JD, Ali TA, Nahid F, Kusano K, Brunette DM (2010) The effect of surface topography on early NFκB signaling in macrophages. J Biomed Mater Res A 95:837–847
Wu J et al (2013) Activation of NLRP3 inflammasome in alveolar macrophages contributes to mechanical stretch-induced lung inflammation and injury. J Immunol 190:3590–3599
Martin D, Bootcov M, Campell T, French P, Breit S (1995) Human macrophages contain a stretch-sensitive potassium channel that is activated by adherence and cytokines. J Membr Biol 315:305–315
Ishii M et al (2009) Epigenetic regulation of the alternatively activated macrophage phenotype. Blood 114:3244–3254
Jetten N et al (2014) Wound administration of m2-polarized macrophages does not improve murine cutaneous healing responses. PLoS One 9:e102994
Mirza R, DiPietro LA, Koh TJ (2009) Selective and specific macrophage ablation is detrimental to wound healing in mice. Am J Pathol 175:2454–2462
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
McWhorter, F.Y., Davis, C.T. & Liu, W.F. Physical and mechanical regulation of macrophage phenotype and function. Cell. Mol. Life Sci. 72, 1303–1316 (2015). https://doi.org/10.1007/s00018-014-1796-8
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00018-014-1796-8