
Abstract
Introduction
Carbon monoxide (CO) released from CORM-2 has anti-inflammatory function, but the critical molecule mediating the inflammation inhibition has not been elucidated. Previous studies indicate that CORM-2 can activate Nrf2, a key transcription factor regulating host defense against oxidative stress and inflammation-related disorders. In this study we use Nrf2 knockout mice to determine the role of Nrf2 in mediating the CO anti-inflammatory action.
Methods
We compared CORM-2’s inhibiting effect on pro-inflammatory cytokine expressions (TNF-α, IL-1β and IL-6 and iNOS) in primary peritoneal macrophages, mouse liver and brain tissues from Nrf2+/+ and Nrf2−/− mice. We further assayed the inflammatory cell infiltration in both liver and brain tissues of the Nrf2+/+ and Nrf2−/− mice. Finally, we examined CORM’s influence on mouse mortality in a mouse sepsis model.
Results
Our results showed that CORM-2 dramatically inhibited the expression of pro-inflammatory cytokines in Nrf2+/+ mice, but not in Nrf2−/− mice. Furthermore CORM-2 substantially decreased LPS-induced mouse mortality of Nrf2+/+ mice, but not of Nrf2−/− mice.
Conclusion
We conclude that Nrf2 is indispensable for CORM-2 inhibition of LPS-induced inflammation.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Sepsis is still one of the major diseases leading to high morbidity and mortality [1]. Data have shown significant growth of sepsis incidence and mortality in the past few decades [2–4]. Lipopolysaccharide (LPS), the principal component of the outer membrane of Gram-negative bacteria, is the main pathogenic factor leading to sepsis [5]. LPS activates several intracellular signaling pathways including NF-κB, AP-1 and MAPK pathways [6]. As a result, the inflammation stimuli induce the expression of inflammatory cytokines, chemokines and extracellular matrix proteins [7]. The regulation of inflammatory cytokines is under a tight control by the body innate immune system. A deregulated inflammatory cytokine production is a cause of many inflammatory diseases [8].
Nrf2 is a basic leucine zipper transcription factor playing a key role in cellular defense against oxidative stresses and electrophilic insults [9]. Under physiological condition, Nrf2 is inactive and degraded through Keap1-mediated ubiquitination. Under oxidative or chemical stress, Nrf2 is dissociated from its inhibitor Keap1, translocates to the nucleus and binds to the antioxidative response element (ARE) located on the promoters of Nrf2 target genes, thereby initiating the transcription and expressions of phase II detoxifying enzymes, such as HO-1, NQO1 and γ-GCS, and deoxidative proteins [10]. Recent studies have indicated that Nrf2 signaling pathway is also involved in attenuating inflammation-associated pathogenesis. During inflammation-mediated tissue damage, the activation of Nrf2 inhibited the production of pro-inflammatory mediators including cytokines, chemokines, cell adhesion molecules, matrix metalloproteinase, cyclooxygenase-2 and iNOS [11]. For instance, Nrf2−/− mice are more sensitive to inflammatory challenges in endotoxin-induced sepsis [12]. In the lung of the Nrf2−/− mice, there is higher expression of cytokines including IL-1α,TNF-β and IL-6 than of the WT mice [13]. In dextran sulfate sodium-induced colitis, Nrf2−/− mice, compared with the WT mice, display substantially increased levels of IL-1β, IL-6, IL-12 and TNF-α [14]. These studies suggest that inflammatory tissue injuries may be aggravated due to disruption or loss of Nrf2 signaling. Sulforaphane and curcumin are able to activate the Nrf2 pathway and inhibit the expression of pro-inflammatory cytokines such as IL-1β, IL-6 and TNF-α [15], which pinpointed the critical role of Nrf2 in inhibiting inflammatory cytokine expression.
The endogenous CO is mainly generated by the activity of constitutive heme oxygenase 2 (HO-2) and inducible heme oxygenase 1 (HO-1) that are responsible for converting heme to biliverdin, iron and CO [16]. In recent years, many studies found that exogenous administration of CO of low concentration was capable of inhibiting LPS-induced production of inflammatory cytokines, both in vivo and in vitro and plays an important role in cytoprotection [17–19]. CO-releasing molecules, which belong to the class of transitional metal carbonyls, were capable of delivering CO and attracted increasing attention for potential pharmaceutical studies and even potential clinic applications [20]. Many studies reported that CO-releasing molecules suppressed LPS-stimulated inflammatory response and played a beneficial role similar to that of CO gas, both in vitro and in animal models of various inflammatory diseases [21, 22]. Although the anti-inflammatory property of CO was well-established, the exact mechanism by which CO exerted its anti-inflammation and protection functions remained elusive. Both exogenous and CO-releasing molecules can activate Nrf2 signaling pathway. Some studies showed that CO exerted its anti-inflammatory effect through interrupting the JNK pathway, but not the cGMP pathway, NO pathway or IL-10 pathway [17, 23].
In this study, we tested the role of Nrf2 in CO-mediated anti-inflammatory action. Our data showed that one of CO-releasing molecules, CORM-2, significantly suppressed LPS-induced inflammation in Nrf2+/+ mice, but not in Nrf2−/− mice. Therefore, our results provided solid evidence indicating that Nrf2 is essential for CO’s anti-inflammatory function.
Materials and methods
Materials
LPS (Escherichia coli 0127:B8) was purchased from Sigma. Antibody to iNOS was purchased from Abcam (Cambridge, MA, USA). Antibodies to phospho-p38, p38, phospho-ERK1/2, ERK1/2, phospho-JNK, JNK, P-NF-ΚB, NF-κB and IκB-α were purchased from Cell Signaling Technology. Antibody to HO-1 was purchased from Stressgen Biotechnologies. Antibody to NQO-1 was purchased from NOVUS Biologicals. Antibody to Nrf2 and γ-GCSm were purchased from Santa Cruz Biotechnology. Antibody to Lamin-B1 and β-actin were purchased from Bioworld Technology. Antibody to ICAM-1 was purchased from R&D Systems. Secondary HRP-conjugated goat anti-rabbit antibody and goat anti-mouse antibody were purchased from Bioworld Technology. Secondary HRP-conjugated rabbit anti-goat antibody was purchased from Santa Cruz Biotechnology.
CORM-2 (mol wt 512.18) was purchased from Sigma-Aldrich (St. Louis, MO, USA) and solubilized in dimethyl sulfoxide. Inactive form of CORM-2 (iCORM-2) was prepared by adding CORM-2 to DMSO and leaving it for 18 h at 37°C in a 5 % CO2-humidified atmosphere to liberate CO. The iCORM-2 solution was finally bubbled with nitrogen to remove the residual CO present in the solution. The inactive form of CORM-2 has no effect on LPS-induced cytokine expression.
Cell culture
The mouse macrophage cell line RAW 264.7 was obtained from the American Type Culture Collection (Manassas, VA). The cells were grown in Dulbecco’s modified Eagle’s medium supplemented with 100 U/ml penicillin, 100 μg/ml streptomycin and 10 % fetal bovine serum. The cells were cultured at 37 °C under 5 % CO2. The cells were used in experiments after 80–90 % confluence was reached.
Peritoneal microphages collection and culture
Mice peritoneal macrophages were harvested from the female WT and Nrf2−/− mice of 6–8 weeks of age. Mice were anesthetized with 2 % isoflurane in oxygen via a facemask. Five to seven ml of cold phosphate-buffered saline and 2–3 ml of air were intraperitoneally injected and the peritoneal lavage fluid was collected and centrifuged at 250×g for 10 min at 4 °C. The cells were resuspended in RPMI 1640 containing 10 % fetal bovine serum, 100 U/ml penicillin and 100 μg/ml streptomycin. The cells were cultured in six-well plates and incubated for 2 h at 37 °C in 5 % CO2 in air. The non-adherent cells were removed by changing the media and the adherent cells were used for the following experimental procedures.
Animals
Nrf2−/− ICR female mice of 6–8 weeks of age (originally from Dr. Yamamoto Masayuki) and wild-type mice (bred from heterozygous Nrf2+/− mice) were used in this study. All experimental protocols were approved by the Institutional Animal Care Committee and were in accordance with the guidelines of Nanjing University Medical School. WT and Nrf2−/− mice were fed on regular sterile chow diet and water ad libitum and were housed under controlled conditions (25 ± 2 °C; 12 h light–dark periods). Genotypes of WT and Nrf2−/− mice were confirmed by PCR of genomic DNA isolated from mouse tail. PCR amplification was carried out by using three sets of primers: 5′-TGGACGGGACTATTGAAGGCTG-3′ (sense primer for both wild-type and Nrf2−/− genotypes), 5′-CGCCTTTTCAGTAGATGGAGG-3′ (antisense primer for wild-type genotype) and 5′-GCGGATTGACCGTAATGGGATAGG-3′ (antisense primer for Nrf2−/− genotype). WT and Nrf2−/− mice were, respectively, randomized into the four groups: control (n = 10); CORM-2 (n = 10); LPS (n = 10); LPS + CORM-2 (n = 10). CORM-2 was administered by intraperitoneal injection 1 h before intraperitoneal administration of LPS. At the end of the study period, animals were euthanized with CO2 inhalation. Subsequently, liver and brain tissues were removed, washed and snap-frozen in liquid nitrogen and stored at −80 °C or fixed in 4 % paraformaldehyde for further analysis. For survival rate study, WT and Nrf2−/− mice were assigned randomly into two groups: LPS (n = 10); LPS + CORM-2 (n = 10). The mouse survival rate was monitored every 6 h for 60 h.
Western blot
The cytoplasmic/nuclear fractionation was separated using nuclear/cytosol fractionation kit (Sangon, shanghai, China) according to the manufacturer’s manual. The protein concentrations were determined by BCA Protein Assay Reagent kit (Pierce, IL, USA). Equal amounts of proteins were separated by 10 % PAGE at 100 V for 100 min in Tris–glycine–SDS running buffer. The gel was transferred for 90 min at 300 mA to PVDF membrane. Membranes were then incubated for 2 h with blocking buffer (5 % nonfat dry milk in TTBS (10 % Tween in Tris-buffered saline), and then incubated overnight in the respective primary antibody. After incubation with primary antibody, the membranes were washed three times in TTBS, then incubated with HRP-conjugated secondary antibodies at room temperature for 2 h and finally washed three times in TTBS. The membranes were visualized using the enhanced chemiluminescence assay (Pierce, IL, USA) according to the manufacturer’s instructions. Image J was used to analyze the intensity of the blots.
RT-PCR
Total RNA was isolated with Trizol reagent (Invitrogen, CA, USA), and single-stranded cDNA was synthesized from 500 ng of total RNA with HiScript Reverse Transcriptase Kit (Vazyme, Jiangsu, China) according to the manufacturer’s protocol. The cDNA was stored in −20 °C. Forward and reverse primers were 5′-CTGGAGGTGCTTGAAGAGTTCCC-3′ and 5′-CGCACAAAGCAGGGCACTG-3′ for NOS2, 5′-GGTGTTCATCCATTCTCTACCCAGCC-3′ and 5′-CTCAGGGAAGAGTCTGGAAA-3′ for TNF-α, 5′-CTGTGTCTTCCTAAAGTATGGGCTGGAC-3′ and 5′-CCACACGTTGACAGCTAGGTTCTGTTC-3′ for IL-1β, 5′-CCTCTGGTCTTCTGGAGTACCATAGCTA-3′ and 5′-TCTGACCACAGTGAGGAATGTCCAC-3′ for IL-6, 5′-TGAAGGAGGCCACCAAGGAGG-3′ and 5′-AGAGGTCACCCAGGTAGCGGG-3′ for HO-1, 5′-AGGCCCGGTGCTGAGTATGTC-3′ and 5′-TGCCTGCTTCACCACCTTCT-3′ for GAPDH. The RT-PCR products were separated on 1 % agarose gel and the intensity of each band was quantified using Image J software. The level of GAPDH was used as an internal standard.
Histology study of mouse liver and brain
The liver and brain specimens harvested from different groups of animals were immersed in 4 % paraformaldehyde solution and embedded in paraffin wax. Tissue sections were cut at a thickness of 5 μm and stained with hematoxylin–eosin (HE). The tissue morphologic characteristics were analyzed with an image analysis system (Olympus, Tokyo, Japan).
Statistical analysis
Data were expressed as mean ± SD and evaluated by one-way ANOVA analysis of variance or Student’s t test. Survival studies were analyzed by using the log-rank test. Statistical significance was accepted at P < 0.05. All analyses were performed using SPSS 18.0 software.
Results
CO inhibits LPS-induced inflammation
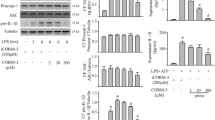
We first tested whether CORM-2-released CO could inhibit LPS-induced pro-inflammatory cytokine expression in RAW264.7 macrophages. As shown in Fig. 1a, the resting RAW264.7 cells expressed minimum, if any, amount of iNOS, TNF-α, IL-1β or IL-6. CORM-2, ranging from 15 to 100 µm, did not change their basal expression levels. LPS dramatically induced their expressions. However, CORM-2 inhibited LPS-induced mRNA levels of TNF-α, IL-1β, IL-6 and iNOS in a dose-dependent manner with 50uM showing a strong inhibition (Fig. 1a). CORM-2 (50 μM) also inhibited LPS-stimulated iNOS expression at protein level in RAW264.7 cells (Fig. 1b; P < 0.05). We then chose 50 μM of CORM-2 for the later experiments.

The effect of CORM-2-released CO on the expression of inflammatory cytokines in RAW264.7 cells. a RAW264.7 cells were pretreated with CORM-2 (15, 50, 100 µM) for 30 min and then stimulated with LPS (1 µg/ml) for 4 h. The inflammatory cytokines TNF-α, IL-1β, IL-6 and iNOS were measured by RT-PCR. b RAW264.7 cells were pretreated with CORM-2 (50 µM) for 30 min and then stimulated with LPS (1 µg/ml) for 16 h for iNOS detection by Western blot. Three separate experiments were performed and the representative results are shown (*P < 0.05)
CORM-2-release CO activates Nrf2 signaling pathway
To confirm CORM-2-released CO activates Nrf2 signaling pathway in our cell system, and we first tested Nrf2 nuclear translocation by measuring Nrf2 protein in nuclear extracts. CORM-2 treatment markedly induced Nrf2 nuclear accumulation 3 h after CORM-2 administration (Fig. 2a). HO-1 is a typical Nrf2 target gene; therefore, we examined the expression of HO-1 after CORM-2 administration. HO-1 expression was induced in a concentration- (15, 50uM and 100 μM) and time-dependent manner (4, 8, 12, and 24 h) (Fig. 2b, c). We also examined the expression of the other two Nrf2 downstream proteins, NQO-1 and γ-GCSm. CORM-2-released CO also induced NQO-1 and γ-GCSm expression in a dose-dependent manner (Fig. 2d), indicating that CORM-2-released CO can effectively activate Nrf2 signaling pathway.

The effect of CORM-2-released CO on Nrf2 pathway activation. a RAW264.7 cells were treated with CORM-2 (50 µM) for 3 h. Nrf2 nuclear translocation was assessed in cellular nuclear fractions. Lamin-B1 was used as a nuclear marker and total Nrf2 as a protein loading control. b The dose–effect of CORM-2-released CO on HO1 induction. RAW264.7 cells were treated with CORM-2 of different doses (15, 50, and 100 µM) for 16 h and HO1 protein induction was assayed by Western blot. c The time course study of CO induction of HO1 expression. RAW264.7 cells were treated with 50 µM CORM-2 for 4, 8, 12, 16 and 24 h and HO1 protein expression was assayed by Western blot. d Nrf2 downstream protein induction by CO. RAW264.7 cells were treated with CORM-2 (15, 50, 100 µM) for 16 h, and NQO1 and γ-GCSm protein levels were detected by Western blot. Three separate experiments were performed and the representative results are shown
CO’s effects on LPS-induced MAPK and NF-κB pathway signaling
Because activation of MAPK pathway is a necessary step for pro-inflammatory cytokine induction by LPS, we examined the potential involvement of this pathway in the inhibitory effects of CORM-2. The p38, ERK1/2 and JNK phosphorylation was assayed using specific antibodies. As shown in Fig. 3a, LPS stimulation rapidly increased phosphorylated forms of p38, ERK1/2 and JNK. Pretreatment of cells with CORM-2 did not significantly affect the phosphorylation of p38, ERK1/2 and JNK, indicating that MAPK pathway might not be the major switch that mediated CO inhibition of inflammatory cytokine induction by LPS.

The effect of CORM-2-released CO on LPS-induced activation of MAPK and NF-κB pathway in RAW264.7 cells. a RAW264.7 cells were pretreated with CORM-2 (50 µM) for 30 min and then stimulated with LPS (1 µg/ml) for 1 h. The phosphorylated and total P38, ERK1/ERK2 and JNK were detected by Western blot. b RAW264.7 cells were pretreated with CORM-2 (50 µM) for 30 min and then stimulated with LPS (1 µg/ml) for 1 h. I-κB-α, phosphorylated NF-κB p65 and total NF-κB p65 were assayed by Western blot. Three separate experiments were performed and the representative results are shown (*P < 0.05)
NF-κB activation is essential for LPS-mediated inflammation. TNF-α, IL-1β, IL-6 and iNOS are all regulated by NF-κB. We then examined the potential involvement of this pathway in the inhibitory effects of CORM-2 on LPS-induced TNF-α, IL-1β, IL-6 and iNOS expression. As shown in Fig. 3b, LPS treatment increased the level of phosphorylated NF-κB (p65), and CORM-2 treatment significantly reduced it (P < 0.05). As NF-κB is regulated by its upstream inhibitor IκB, we also examined the expression of IκB-α. It seemed that CORM-2 alone increased IκB-α protein levels, but CORM-2 did not have significant effect on LPS-induced IκB-α degradation. These results indicated that CO might affect the NF-kB pathway signaling at the p65 activation step.
CO inhibition of LPS-induced inflammation in macrophage is Nrf2 dependent
To clarify the role of Nrf2 in the CORM-2 inhibition of LPS-induced cytokine expression, we performed series experiments on the peritoneal macrophages from Nrf2 knockout mice (Nrf2−/−) and the control littermates. We first confirmed the mouse genotypes by PCR (Fig. 4a). We then tested LPS-induced inflammatory cytokine and HO-1 expression with or without CORM-2 by RT-PCR. When mouse peritoneal macrophages were stimulated with LPS, iNOS, TNF-α, IL-1β, IL-6 and HO-1 mRNA levels were all markedly induced in both WT and Nrf2−/− cells. As expected, Nrf2−/− cells displayed even higher cytokine mRNA levels. CORM-2 treatment significantly inhibited the mRNA levels of iNOS, TNF-α, IL-1β and IL-6 in peritoneal macrophages from WT mice, but not Nrf2−/− mice. In addition, CORM2 treatment did not obviously induce HO-1 expression in Nrf2−/− macrophages,suggesting that Nrf2 was the critical molecule mediating CO’s transcription expression. CORM2 significantly enhanced LPS-induced HO-1 expression in WT macrophages, but not in Nrf2−/− macrophages (Fig. 4b, c; *P < 0.05), suggesting that both signal transducer and transcription factor capacities of Nrf2 contributed to its anti-inflammation functions.

The effect of CORM-2-released CO on LPS-induced expression of inflammatory cytokines in macrophages of WT and Nrf2−/− mice. a PCR genotyping of Nrf2+/+, Nrf2+/− and Nrf2−/− mice. b RT-PCR analysis of inflammatory cytokine expression of mouse peritoneal macrophages. The peritoneal microphages from WT and Nrf2−/− mice pretreated with CORM-2 (50 μM) for 1 h and then stimulated with LPS (1 mg/ml) for 4 h. The inflammatory cytokines TNF-α, IL-1β, IL-6,iNOS and HO-1 were assayed by RT-PCR. Three mice in each group were assayed and the representative results from one mouse from each group are shown. c The quantitation results are presented as mean ± SD. Represented data are shown. At least three independent experiments were performed. *P < 0.05
CO inhibition of LPS-induced inflammation in mouse liver and brain is Nrf2 dependent
To confirm these results in vivo, we injected LPS to WT and Nrf2−/− mice in the presence or absence of CORM-2, and then tested pro-inflammatory cytokine and HO-1 mRNA levels from mouse liver and brain. Mice injected with LPS alone displayed a marked induction of inflammatory cytokine TNF-α, IL-1β and iNOS in the liver and brain. CORM-2 significantly inhibited their levels in WT mice livers, but not in Nrf2−/− mice (Fig. 5a, b; *P < 0.05). LPS induced HO-1 mRNA expression both in WT and Nrf2−/− mice. CORM-2 significantly increased HO-1 mRNA levels in WT mice, but not in Nrf2−/− mice. The mouse brain exhibited a similar CORM-2 effect on LPS-induced cytokine and HO-1 expression. (Fig. 5c, d; *P < 0.05). We also examined mouse liver for iNOS and ICAM-1 protein expression. Similarly, the protein levels of iNOS and ICAM-1 were induced by LPS in both WT and Nrf2−/− mice. Consistent with mRNA results, CORM-2 strongly inhibited iNOS and ICAM-1 protein expression in WT mice, but not in Nrf2−/− mice (Fig. 5e, f; *P < 0.05).

The effect of CORM-2-released CO on LPS-induced expression of inflammatory cytokines in the liver and brain of WT and Nrf2−/− mice. a RT-PCR analysis of inflammatory cytokine expression of mouse livers. The WT and Nrf2−/− mouse livers were collected 4 h after LPS (10 mg/kg) treatment with or without CORM-2 (30 mg/kg) of 1 h pretreatment. The expression of inflammatory cytokines iNOS, IL-1β, IL-6 and HO-1 were detected by RT-PCR. b The quantitation results are presented as mean ± SD. *P < 0.05. c RT-PCR analysis of inflammatory cytokine expression of mouse brains. The WT and Nrf2−/− mouse brains were collected 4 h after LPS (10 mg/kg) treatment with or without CORM-2 (30 mg/kg). The expression of inflammatory cytokines TNF-α, IL-1β, IL-6 and HO-1 were detected by RT-PCR. d The quantitation results were presented as mean ± SD, *P < 0.05. e Western blot assay of iNOS and ICAM-1 protein expression in mouse livers. The WT and Nrf2−/− mice were pretreated with CORM-2 (30 mg/kg) intraperitoneally for 1 h and then challenged with LPS (10 mg/kg) for 24 h. Protein expression of iNOS and ICAM-1 were analyzed by Western blot. f The quantitation results are presented as mean ± SD. At least three independent experiments were performed. *P < 0.05. NS no significant difference
CO inhibition of LPS-induced sepsis is Nrf2 dependent
To further explore the role of Nrf2 in CORM-2′s anti-inflammatory function in vivo, we employed a well-established mouse model of LPS- mediated sepsis. In wild-type mouse, a lethal dose of LPS injection caused 40 and 100 % mouse death in 12 and 30 h, respectively. Although short-term CORM-2 treatment (1 day before LPS injection, or simultaneous CORM2 and LPS injections) only had marginal improvement on mouse mortality, long-term CORM-2 treatment (2 days before LPS injection) markedly delayed mice death until 36 h and decreased mortality by 60 % overall (Fig. 6a). However in Nrf2−/− mice, LPS caused a 100 % Nrf2−/− mice mortality as early as 18 h. Although CORM-2 treatment delayed mouse death, a 100 % mortality was reached by 36 h after LPS injection (Fig. 6b). HE staining also showed that LPS administration caused markedly inflammatory cell infiltration in mouse liver (Fig. 6c) and brain (Fig. 6d) compared to control mice. Administration of CORM-2 markedly decreased the inflammatory cell infiltration in WT mice, but not in Nrf2−/− mice. We also analyzed the expression of inflammatory cytokine TNF-α, IL-6 and HO-1 mRNA levels of mouse liver under such a setting. Similarly, CORM-2 significantly improved LPS-induced TNF-α, IL-6 and HO-1 mRNAs in the liver of WT mice, but not in Nrf2−/− mice (Fig. 6e; *P < 0.05).

The effect of CORM-2-released CO on Nrf2+/+ and Nrf2−/− mice in a mouse sepsis model. Nrf2+/+ (a) and Nrf2−/− mice (b) (10 mice in each group) were pretreated with CORM-2 (30 mg/kg) intraperitoneally daily for 2 days and then injected with a lethal dose of LPS (60 mg/kg). Every 6 h the survival mouse numbers were counted and survival rate was calculated (c, d). The effect of CORM-2-released CO on mouse liver (c) and brain (d) of LPS-induced inflammatory responses in Nrf2+/+ and Nrf2−/− mice. The mouse liver and brain were collected 4 h after LPS injection and prepared for HE staining. Mice injected with LPS showed inflammatory cell infiltration (indicated by arrowhead). The liver and brain (cerebral cortex) sections that had received CORM-2 showed marked decrease of granulocyte infiltration in Nrf2+/+ mice, but not in Nrf2−/− mice. e the WT and Nrf2−/− mice were treated with or without CORM-2 (30 mg/kg) daily for 2 days before LPS (60 mg/kg) was injected intraperitoneally for 4 h. The mouse livers were collected for the detection of TNF-α, IL-6 and HO-1 by RT-PCR. The quantitation results were presented as mean ± SD. At least three independent experiments were performed. *P < 0.05
Discussion
In the present study, we demonstrated that CORM-2-released CO significantly inhibited the LPS-induced production of pro-inflammatory cytokines in peritoneal macrophages, liver and brain of WT mice, while no significant changes were observed in Nrf2−/− mice. CORM-2-released CO also reduced the LPS-induced neutrophil infiltration in the liver and brain of WT mice, but not in the Nrf2−/− mice. Taken together, the current findings suggest that Nrf2 is essential for CO anti-inflammatory activity in LPS-induced inflammation.
Previously, Otterbein group discovered that low concentration of CO gas had anti-inflammatory effects [17]. Numerous studies also showed that low dose of CO can exert potent anti-inflammatory effects in both in vitro and in vivo models of LPS-induced inflammations, including chronic colitis, aeroallergen-induced inflammation and thrombin-induced neuro-inflammation [22, 24–28]. However, it is not fully clear how significantly the Nrf2 signal pathway contributed to this inhibition. Our study provides solid evidence that Nrf2 is indispensible for CO inhibition of LPS-induced inflammation.
MAPKs are believed to play a significant role in the regulation of LPS-induced inflammatory response. Some studies proposed that CO inhibition of inflammation involved the activation of MAP kinase pathway [17], which normally correlated with increased inflammation. Our study showed that CORM-2 treatment caused slightly reduced p38 phosphorylation, but had no obvious inhibitory effect on both phosphorylation of ERK1/2 or JNK. Some studies demonstrated that blocking phosphatidylinositol-3-phosphate kinase pathway could attenuate CO’s anti-inflammatory effect, while blocking ERK pathway could reinforce it [29]. LPS-induced NF-κB activation meditates the release of many pro-inflammatory cytokines including TNF-α, IL-1β and IL-6. Studies showed that CO inhibited LPS-induced granulocyte macrophage colony-stimulating factor induction via inhibiting the activation of the transcription factor NF-κB and preventing the phosphorylation and degradation of the regulatory subunit IκB-α [30]. One study showed that CO inhibited the expression of pro-inflammatory cytokines and iNOS by inhibiting NF-κB/DNA binding as well as the I κB-α phosphorylation and degradation in Caco-2 cells [31]. These findings reflect the complexity of CO’s interruption with different inflammatory pathways, but did not reveal the upstream molecule that CO acts on. In this study, we confirmed that the NF-kB pathway at the p65 level was involved in CORM-2-mediated inflammatory inhibition, but MAP kinase pathway was less involved in the process. We further found that Nrf2 is essential for CO’s anti-inflammatory function. We speculated that Nrf2 might act upstream of MAP kinase and NF-kB pathways in the inhibition of LPS-induced inflammatory cytokine expression.
The Nrf2 pathway is critical for maintaining intracellular homeostasis and suppressing oxidative stress. In addition to its role in regulating redox levels, accumulated studies have reported that Nrf2 can increase the expression of a variety of phase II detoxifying enzymes including HO-1 and NQO-1, and subsequently alleviate inflammatory tissue damage. For example, Nrf2 suppressed TNF-α-induced monocyte chemoattractant protein-1 and vascular cell adhesion molecule-1 expression and inhibited TNF-α-induced monocytic U937 cell adhesion to human aortic endothelial cells. Nrf2 also inhibited IL-1β-induced monocyte chemoattractant protein-1 gene expression in human mesangial cells [32]. CDDO-Imidazolide, polyunsaturated fatty acids, docosahexaenoic acid, eicosapentaenoic acid, sulforaphane and triterpenoids were able to inhibit LPS-induced inflammatory response via activating the Nrf2 pathway [33–36]. Our experiments confirmed that CORM-2 activated Nrf2 by inducing its nuclear translocation and increasing the expression of Nrf2′s downstream genes such as HO-1,NQO-1 and γ-GCSm. Previous studies showed that Nrf2−/− mice are more susceptible to allergen-driven airway inflammation, asthma, airway inflammation induced by low-dose diesel exhaust particle and dextran sulfate sodium-induced experimental colitis [37–39]. One recent study further demonstrated that inhaled CO can rescue mice from lethal Staphylococcus aureus sepsis through Nrf2 [40]. Our study was in line with this observation and indicated that Nrf2 was critical in CO’s anti-inflammatory effects. We have previously shown that CO inhalation reduced stroke-induced brain damage through the Nrf2 pathway [41]. The current study extended this observation and showed that Nrf2 molecule was essential for CO’s anti-inflammation not only in stroke-induced brain inflammation, but also in systematic sepsis. We further found that CORM2 could not alleviate LPS-induced inflammatory cytokine induction, or significantly enhance HO-1 expression in Nrf2−/− mice, suggesting that both signal transducer and transcription factor functions of Nrf2 contributed to its anti-inflammatory effects. Therefore, Nrf2 could be a good target for potential anti-sepsis therapies.
Although our results confirmed that Nrf2 was essential for CO inhibition of LPS-induced inflammatory cytokine expression, the connection between Nrf2 and inflammatory signaling pathway has still not been fully resolved. LPS can directly activate the major innate immune signaling pathways, such as NF-κB and MAP kinase pathways. Some studies showed that the embryonic fibroblasts and peritoneal macrophages in Nrf2−/− mice had stronger activation in reaction to LPS stimulus and a greater rise in nuclear levels of p65 in the LPS-treated lungs of Nrf2−/− mice than those of WT mice. The degradation of IκB-α needed for NF-κB activation was more obvious in Nrf2−/− mice embryonic fibroblasts induced by LPS [13]. Nrf2−/− mice showed more NF-κB DNA binding in their brains and lungs after traumatic brain injury in comparison with the WT Nrf2 counterparts [42, 43]. These observations suggested that CO acted at the upper level above NF-kB along the signaling. The complex relation between Nrf2 and NF-κB pathway deserves further investigation.
In conclusion, our results indicate that CORM2-released CO can mitigate LPS-induced inflammation through activating Nrf2, which functions as both a signaling transducer that reduces inflammatory signaling and a transcription factor that negatively regulates inflammation response through downstream proteins. Our study presented direct evidence showing that the anti-inflammatory effect of CO essentially relied on Nrf2 and uncovered the anti-inflammatory mechanism of CO in LPS-induced inflammation. The study provided new insights for designing a strategy for Nrf2-based anti-inflammation therapy.
Abbreviations
- ARE:
-
Antioxidant-responsive element
- CO:
-
Carbon monoxide
- CORM-2:
-
Carbon monoxide-releasing molecule-2
- ERK:
-
Extracellular-regulated protein kinases
- HO-1:
-
Heme oxygenase 1
- HO-2:
-
Heme oxygenase 2
- IL-1β:
-
Interleukin-1β
- IL-6:
-
Interleukin-6
- iNOS:
-
Inducible nitric oxide synthase
- ICAM-1:
-
Intercellular cell adhesion molecule-1
- IκB-α:
-
Inhibitor of nuclear factor kappa B-alpha
- JNK:
-
C-jun N-terminal kinase
- Keap1:
-
Kelch-like ECH-associated protein 1
- LPS:
-
Lipopolysaccharide
- MAPK:
-
Mitogen-activated protein kinase
- NF-kB:
-
Nuclear factor kappa B
- Nrf2:
-
Nuclear factor-erythroid 2-related factor-2
- NQO1:
-
NAD(P)H:quinine oxidoreductase-1
- p38:
-
P38-Mitogen-activated protein kinase
- TNF-α:
-
Tumor necrosis factor α
- γ-GCS:
-
γ-Glutamylcysteine synthetase
References
Kung H-C, et al. Deaths: final data for 2005. Natl Vital Stat Rep. 2008;56(10):1–120.
Dombrovskiy VY, et al. Rapid increase in hospitalization and mortality rates for severe sepsis in the United States: a trend analysis from 1993–2003. Crit Care Med. 2007;35(5):1244–50.
Control CFD. Increase in National Hospital Discharge Survey rates for septicemia-United States, 1979–1987. MMWR Morb Mortal Wkly Rep. 1990;39:31–4.
Martin GS, et al. The epidemiology of sepsis in the United States from 1979 through 2000. N Engl J Med. 2003;348(16):1546–54.
Cohen J. The immunopathogenesis of sepsis. Nature. 2002;420(6917):885–91.
Guha M, Mackman N. LPS induction of gene expression in human monocytes. Cell Signal. 2001;13(2):85–94.
Rahman I, Biswas SK, Kirkham PA. Regulation of inflammation and redox signaling by dietary polyphenols. Biochem Pharmacol. 2006;72(11):1439–52.
Martinon F, Tschopp J. Inflammatory caspases: linking an intracellular innate immune system to autoinflammatory diseases. Cell. 2004;117(5):561–74.
Lee J-M, Johnson JA. An important role of Nrf2-ARE pathway in the cellular defense mechanism. BMB Reports. 2004;37(2):139–43.
Chen X-L, Kunsch C. Induction of cytoprotective genes through Nrf2/antioxidant response element pathway: a new therapeutic approach for the treatment of inflammatory diseases. Curr Pharm Des. 2004;10(8):879–91.
Kim J, Cha Y-N, Surh Y-J. A protective role of nuclear factor-erythroid 2-related factor-2 (Nrf2) in inflammatory disorders. Muta Res Fundam Mol Mech Mutagen. 2010;690(1):12–23.
Hernandez ML, et al. Comparative airway inflammatory response of normal volunteers to ozone and lipopolysaccharide challenge. Inhalation Toxicol. 2010;22(8):648–56.
Thimmulappa RK, et al. Nrf2 is a critical regulator of the innate immune response and survival during experimental sepsis. J Clin Invest. 2006;116(4):984–95.
Osburn WO, et al. Increased colonic inflammatory injury and formation of aberrant crypt foci in Nrf2-deficient mice upon dextran sulfate treatment. Int J Cancer. 2007;121(9):1883–91.
Yoshida S-I, et al. Inhibition of IL-6 and IL-8 induction from cultured rheumatoid synovial fibroblasts by treatment with aurothioglucose. Int Immunol. 1999;11(2):151–8.
Motterlini R, Otterbein LE. The therapeutic potential of carbon monoxide. Nat Rev Drug Discovery. 2010;9(9):728–43.
Otterbein LE, et al. Carbon monoxide has anti-inflammatory effects involving the mitogen-activated protein kinase pathway. Nat Med. 2000;6(4):422–8.
Lee T-S, Chau L-Y. Heme oxygenase-1 mediates the anti-inflammatory effect of interleukin-10 in mice. Nat Med. 2002;8(3):240–6.
Otterbein LE, et al. Heme oxygenase-1: unleashing the protective properties of heme. Trends Immunol. 2003;24(8):449–55.
Motterlini R, et al. Bioactivity and pharmacological actions of carbon monoxide-releasing molecules. Curr Pharm Des. 2003;9(30):2525–39.
Sawle P, et al. Carbon monoxide-releasing molecules (CO-RMs) attenuate the inflammatory response elicited by lipopolysaccharide in RAW264.7 murine macrophages. Br J Pharmacol. 2005;145(6):800–10.
Cepinskas G, et al. Carbon monoxide liberated from carbon monoxide-releasing molecule CORM-2 attenuates inflammation in the liver of septic mice. American Journal of Physiology-Gastrointestinal and Liver. Physiology. 2008;294(1):G184–91.
Morse D, et al. Suppression of inflammatory cytokine production by carbon monoxide involves the JNK pathway and AP-1. J Biol Chem. 2003;278(39):36993–8.
Mayr FB, et al. Effects of carbon monoxide inhalation during experimental endotoxemia in humans. Am J Respir Crit Care Med. 2005;171(4):354–60.
Sarady JK, et al. Carbon monoxide protection against endotoxic shock involves reciprocal effects on iNOS in the lung and liver. FASEB J. 2004;18(7):854–6.
Hegazi RA, et al. Carbon monoxide ameliorates chronic murine colitis through a heme oxygenase 1–dependent pathway. J Exp Med. 2005;202(12):1703–13.
Chapman JT, et al. Carbon monoxide attenuates aeroallergen-induced inflammation in mice. Am J Physiol Lung Cell Mol Physiol. 2001;281(1):L209–16.
Bani-Hani MG, et al. Modulation of thrombin-induced neuroinflammation in BV-2 microglia by carbon monoxide-releasing molecule 3. J Pharmacol Exp Ther. 2006;318(3):1315–22.
Bani-Hani MG, et al. A carbon monoxide-releasing molecule (CORM-3) attenuates lipopolysaccharide-and interferon-gamma-induced inflammation in microglia. Pharmacol Rep. 2006;58:132.
Sarady JK, et al. Carbon monoxide modulates endotoxin-induced production of granulocyte macrophage colony-stimulating factor in macrophages. Am J Respir Cell Mol Biol. 2002;27(6):739–45.
Megias J, Busserolles J, Alcaraz M. The carbon monoxide-releasing molecule CORM-2 inhibits the inflammatory response induced by cytokines in Caco-2 cells. Br J Pharmacol. 2007;150(8):977–86.
Chen X-L, et al. Activation of Nrf2/ARE pathway protects endothelial cells from oxidant injury and inhibits inflammatory gene expression. Am J Physiol Heart Circ Physiol. 2006;290(5):H1862–70.
Thimmulappa RK, et al. Nrf2-dependent protection from LPS induced inflammatory response and mortality by CDDO-Imidazolide. Biochem Biophys Res Commun. 2006;351(4):883–9.
Wang H, et al. Role of Nrf2 in suppressing LPS-induced inflammation in mouse peritoneal macrophages by polyunsaturated fatty acids docosahexaenoic acid and eicosapentaenoic acid. Mol Pharm. 2010;7(6):2185–93.
Lin W, et al. Sulforaphane suppressed LPS-induced inflammation in mouse peritoneal macrophages through Nrf2 dependent pathway. Biochem Pharmacol. 2008;76(8):967–73.
Thimmulappa RK, et al. Preclinical evaluation of targeting the Nrf2 pathway by triterpenoids (CDDO-Im and CDDO-Me) for protection from LPS-induced inflammatory response and reactive oxygen species in human peripheral blood mononuclear cells and neutrophils. Antioxid Redox Signal. 2007;9(11):1963–70.
Li YJ, et al. Disruption of Nrf2 enhances susceptibility to airway inflammatory responses induced by low-dose diesel exhaust particles in mice. Clin Immunol. 2008;128(3):366–73.
Rangasamy T, et al. Disruption of Nrf2 enhances susceptibility to severe airway inflammation and asthma in mice. J Exp Med. 2005;202(1):47–59.
Khor TO, et al. Nrf2-deficient mice have an increased susceptibility to dextran sulfate sodium-induced colitis. Cancer Res. 2006;66(24):11580–4.
MacGarvey NC, et al. Activation of mitochondrial biogenesis by heme oxygenase-1-mediated NF-E2-related factor-2 induction rescues mice from lethal Staphylococcus aureus sepsis. Am J Respir Crit Care Med. 2012;185(8):851–61.
Wang B, et al. Carbon monoxide–activated Nrf2 pathway leads to protection against permanent focal cerebral ischemia. Stroke. 2011;42(9):2605–10.
Mao L, et al. Disruption of Nrf2 enhances the upregulation of nuclear factor-kappaB activity, tumor necrosis factor-α, and matrix metalloproteinase-9 after spinal cord injury in mice. Mediat Inflamm. 2010;2010:238321. doi:10.1155/2010/238321.
Jin W, et al. Influence of Nrf2 genotype on pulmonary NF-κB activity and inflammatory response after traumatic brain injury. Ann Clin Lab Sci. 2008;38(3):221–7.
Acknowledgments
This work was supported by research grants from the National Nature Science Foundation of China (Grant No. 81271301, 81470940, 21372233 and 81070940) and by the open fund of State Key Laboratory of Pharmaceutical Biotechnology, Nanjing University, China (Grant No. KF-GN-201402).
Conflict of interest
The authors state no conflict of interest.
Author information
Authors and Affiliations
Corresponding author
Additional information
Responsible Editor: Yoshiya Tanaka.
S. Qin and R. Du made equal contributions.
Rights and permissions
About this article

Cite this article
Qin, S., Du, R., Yin, S. et al. Nrf2 is essential for the anti-inflammatory effect of carbon monoxide in LPS-induced inflammation. Inflamm. Res. 64, 537–548 (2015). https://doi.org/10.1007/s00011-015-0834-9
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00011-015-0834-9