
Abstract
Objective
To observe the therapeutic effect of RelB-silenced dendritic cells (DCs) in experimental autoimmune myasthenia gravis (EAMG), and further to investigate the mechanism of this immune system therapeutic.
Methods
(1) RelB-silenced DCs and control DCs were prepared and the supernatants were collected for IL-12p70, IL-6, and IL-23 measurement by ELISA. (2) RelB-silenced DCs and control DCs were co-cultured with AChR-specific T cells, and the supernatant was collected to observe the IL-17, IFN-γ, IL-4 production. (3) EAMG mice with clinical scores of 1 to 2 were collected and enrolled randomly into the RelB-silenced DC group or the control DC group. RelB-silenced DCs or an equal amount of control DCs were injected intravenously on days 0, 7, and 14 after enrollment. Clinical scores were evaluated every other day. Twenty days after allotment, serum from individual mice was collected to detect serum concentrations of anti-mouse AChR IgG, IgG1, IgG2b, and IgG2c. The splenocytes were isolated for analysis of lymphocyte proliferative responses, cytokine (IL-17, IFN-γ, IL-4) production, and protein levels of RORγt, T-bet, GATA-3, and FoxP3 (the special transcription factors of Th17, Th1, Th2, and Treg, respectively).
Results
(1) RelB-silenced DCs produced significantly reduced amounts of IL-12p70, IL-6, and IL-23, as compared with control DCs. (2) RelB-silenced DCs spurred on the CD4+ T cells from Th1/Th17 to the Th2 cell subset in the co-culture system. (3) Treatment with RelB-silenced DCs effectively reduced myasthenic symptoms and levels of serum anti-acetylcholine receptor autoantibody in mice with ongoing EAMG. Th17-related markers (RORγt, IL-17) and Th1-related markers (T-bet, IFN-γ) were downregulated, whereas Th2 markers (IL-4 and GATA3) and Treg marker (FoxP3) were upregulated.
Conclusions
RelB-silenced DCs were able to create a particular cytokine environment that was absent of inflammatory cytokines. RelB-silenced DCs provide a practical means to normalize the differentiation of the four T-cell subsets (Th17, Th1, Th2, and Treg) in vivo, and thus possess therapeutic potential in Th1/Th17-dominant autoimmune disorders such as myasthenia gravis.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
The human autoimmune disease myasthenia gravis (MG) is a T cell-dependent, acetylcholine receptor antibody-mediated autoimmune neuromuscular disease [1]. Experimental autoimmune myasthenia gravis (EAMG) induced in C57BL/6 mice after repeated immunizations with emulsified Torpedo Californica AChR (TAChR) is a useful model for the study of pathogenic mechanisms and therapeutic strategies relevant to MG in humans [2]. Although antibodies to AChR are directly responsible for the destruction of the muscle endplate resulting in both MG and EAMG, the autoantibody response is T cell dependent, with CD4+ T cells providing help for B cells to produce anti-AChR antibodies [3, 4]. Traditionally, Th cells were divided into the Th1, Th2, and regulatory T-cell (Treg) functional subsets [5, 6]. Recently, Th17 cells, the IL-17-producing Th subset, have been identified, and the traditional classification of Th cells has been supplemented [7]. The Thl, Th2, and Treg cells are associated with special transcription factors T-bet, GATA3, and FoxP3, respectively, and it has now been confirmed that ROR-γt (retinoid-related orphan receptors-γt) is the special transcription factor that specially regulates the differentiation of Thl7 cells [8]. Recent studies indicate that Th17, Th1, Th2, and Treg cells play a very important role in the development and progression of EAMG via a complex network of interactions among the cells and their cytokines [9, 10]. Thus, modulation of the differentiation of the four T-cell subsets in vivo would be an alternative to treatment of this autoimmune disorder.
Dendritic cells (DCs), the most potent antigen-presenting cells (APCs), have important functions in T-cell immunity and T-cell tolerance. Because of their expression of soluble or membrane-bound T-cell-polarizing molecules, DCs exert an important function in determining the balance among Th1, Th2, Th17, and Treg development and have become an attractive cell type for therapeutic manipulation of the immune system in order to enhance insufficient immune responses in infectious diseases and cancer or to attenuate excessive immune responses in allergy, autoimmunity, and transplantation [11].
We previously demonstrated that RelB (an NF-κB family member that is responsible for DC differentiation)-silenced DCs had profound effects on the induction of EAMG partly by enhancing IL-10 and IL-4 expression [12]. In this study, we investigated the therapeutic effect of RelB-silenced DCs in an established EAMG. To have a deep investigation on the immune-modulation mechanism of RelB-silenced DCs, we have observed the effector cytokines (IL-17, IFN-γ, IL-4) and special transcription factors (ROR-γt, T-bet, GATA3, FoxP3) of Th17, Th1, Th2, and Treg. Our data show that RelB-silenced DCs pulsed with Tα146–162 peptide effectively reduced myasthenic symptoms in mice with ongoing EAMG. The protective effect was associated with a shift of a T helper Th1/Th17 to a Th2 and FoxP3+ regulatory T cell profile.
Materials and methods
Animals and reagents
Eight-week-old female C57BL/6 mice were purchased from Shanghai Slac Laboratory Animal Co. and kept under pathogen-free conditions at the animal core facility of the Central South University. All experiments were performed according to the Animal Care and Use Committee guidelines. Tα146-162 (Leu-Gly-Ile-Trp-Thr-Tyr-Asp-Gly-Tlu-Lys-Val-Ser-Ile-Ser-Pro-Glu-Ser) peptide was synthesized at the GL Biochem (Shanghai, China) and purified by HPLC to >95% purity. TAChR and mouse AChR were purified from the electric organ of Torpedo or mouse muscle by neurotoxin affinity column [13, 14]. Complete Freund’s adjuvant was obtained from Sigma–Aldrich.
Preparing and assaying cytokines of RelB-silenced BMDCs
RelB-silenced DCs and control DCs were prepared as described previously [12]. Briefly, bone marrow dendritic cells (BMDCs) were flushed from the femurs and tibiae of C57BL/6 mice, then cultured in the medium with 10 ng/ml recombinant murine GM-CSF (PeproTech) and 10 ng/ml IL-4 (PeproTech). After 120 h, BMDCs were purified by positive immunomagnetic selection with anti-CD11c-conjugated MicroBeads (Miltenyi Biotec, Germany). Purified DCs were plated in a 12-well plate containing 1 ml of medium supplemented with mGM-CSF/mIL-4 and polybrene (5 μg/ml). Concentrated lentiviruses containing RelB-specific small interfering RNA (siRNA) (CGATGGCTTTGCCTATGAT) or nonsilencing siRNA (TTCTCCGAACGTGTCACGT) were then added to the cells at a multiplicity of infection (MOI) of 50. At 5 days post-transduction, DCs were stimulated by LPS (1 μg/ml) for 48 h. The resulting DCs were designated as RelB-silenced DCs or control DCs, respectively. Supernatants were collected for IL-12p70, IL-6, and IL-23 measurement by ELISA in accordance with the manufacturer’s instructions (eBioscience, USA).
TAChR-activated T-cell enrichment and DC T-cell co-cultures in vitro
Eight-week-old female C57BL/6 mice were immunized in the footpads with 20 μg TAChR in CFA. After 7 days, single cells from the pooled draining (popliteal and inguinal) lymph nodes were collected, and CD4+ T cells were positively selected magnetically with mAbs directly bound to CD4 MACS MicroBeads (Miltenyi Biotec, Germany) according to the manufacturer’s protocols. The cell purity was examined by flow cytometry (>92%). RelB-silenced DCs (or control DCs) were cultured with 4 × 104 CD4+ T cells in each well of 48-well plates in the presence of TAChR (5 μg/ml) or Tα146–162 peptide (50 μg/ml). After co-culture for 96 h, the supernatant was used for analysis of IFN-γ, IL-4, and IL-17 by ELISA in accordance with the manufacturer’s instructions (BD Pharmingen).
Induction of EAMG and treatment with transduced DCs
Fifty 8-week-old female C57BL/6 mice were immunized with TAChR (20 μg per mouse) emulsified in CFA on days 0, 30, and 60. Mice were evaluated for clinical EAMG, and their muscle weakness was graded as follows [13]: grade 0: normal strength without any muscle weakness; grade 1: definite weakness of the forelimbs with a characteristic hunched, chin-down posture and flaccid tail after exercise involving 20–30 consecutive paw grips on the top steel grids of the animal cage; grade 2: grade 1 weakness at rest; and grade 3: severe weakness with paralysis involving all the limbs (quadriplegia) accompanied by dehydration and moribund state. On day 70, 24 mice that had developed clinical EAMG clinical scores of 1 to 2 were randomly allotted into two groups, each with equivalent grades. RelB-silenced DCs (2 × 106/0.2 ml PBS each mouse) or equal amounts of control DCs were pulsed overnight with Tα146–162 peptide (50 μg/ml), washed, and injected intravenously on days 0, 7, and 14 after allotment. Clinical scores were evaluated every other day. At day 20 after allotment, serum from individual mice was collected to detect serum concentrations of anti-mouse AChR IgG, IgG1, IgG2b, and IgG2c by ELISA. At day 21 after allotment, splenocytes were isolated for analysis of TAChR- and Tα146–162-induced proliferative responses, cytokine production, and protein levels of T-bet, GATA-3, RORγt, and FoxP3.
ELISA for anti-AChR Ig isotypes
Affinity-purified mouse-AChR (0.5 μg/ml) was coated onto a 96-well micro titer plate in 0.1 M carbonate bicarbonate buffer (pH 9.6) overnight at 4°C. The plates were blocked with 2% BSA in PBS at room temperature for 30 min. A total of 100 μl of diluted serum samples (1:2,000) was added and incubated at 37°C for 90 min. After four washes, HRP-conjugated anti-mouse IgG, IgG1, IgG2b, or IgG2c (southern Biotech, USA), diluted 1/500 in PBS 0.05% Tween-20, was added and incubated at 37°C for 90 min. Subsequently, 0.3 mg/ml ABTS (diammonium) substrate solution was added to the IgG, IgG1, and IgG2b plates, and 100 μl of avidin-labeled peroxidase (Sigma–Aldrich) at 2.5 μg/ml was added to the IgG2c plate and incubated for 30 min. Finally, the peroxidase indicator substrate ABTS, in 0.1 M citric buffer, pH 4.35, was added in the presence of H2O2, and the mixture was allowed to develop at room temperature in the dark. Plates were read at a wavelength of 405 nm. The results were expressed as OD values.
AChR-specific T-cell proliferation and cytokine assays
Splenocytes were harvested and single cell suspensions were prepared. A total of 4 × 105 cells/well were seeded into flat-bottom 96-well plates in RPMI 1640 medium supplemented with 10% FBS, 25 mM HEPES buffer, 2-ME (3 × 10 μM), penicillin (100 U/ml), and streptomycin (100 μg/ml) (complete medium), and stimulated with TAChR (5 μg/ml) or dominant peptide Tα146–162 (50 μg/ml) or ConA (4 μg/ml). Cells were incubated at 37°C in 5% CO2 for 5 days with the addition of [3H]thymidine in the final 16 h of culture. For measurement of cytokine production, cells at a concentration of 4 × 106/ml were seeded into 48-well plates in complete medium with 2.5 μg/ml TAChR or 20 μg/ml dominant peptide Tα146–162. Culture supernatants were analyzed for IFN-γ, IL-4, and IL-17 by ELISA in accordance with the manufacturer’s instructions (BD Pharmingen).
SDS-PAGE and Western blot analysis
Splenocytes were collected, centrifuged into a pellet, and lysed for 30 min on ice, and then were centrifuged again at 12,000 rpm for another 30 min at 4°C. The protein levels were measured by using a BCA protein assay kit (HyClone-Pierce, USA). The protein was separated by a vertical dodecyl sulphate-polyacrylamide gel electrophoresis (SDS-PAGE) and subsequently transferred to a PVDF membrane (Millipore, Bedford, MA, USA). The membrane was blocked in TBS/5% skim milk overnight and then incubated with the appropriate primary antibody (rabbit anti-mouse RelB, Cell Signaling, USA; rat anti-mouse RORγt, rat anti-mouse FoxP3, eBioscience, USA; goat anti-mouse T-bet, GATA-3, Santa Cruz, USA) overnight at 4°C. After washing with TBS/Tween, the membrane was incubated with horseradish peroxidase-conjugated IgG antibody (Pierce, Rockford, IL, USA). After another washing, proteins were detected with an ECL Western blotting detection kit.
Statistical analysis
Data are expressed as mean ± SD. Differences between groups were analyzed using SPSS software for Windows (version 11.0) by unpaired two-tailed parametric Student’s t test or ANOVA test. P values below 0.05 were considered statistically significant.
Results
Altered cytokine production in RelB-silenced DCs
As shown in Fig. 1, RelB-silenced DCs produced significantly reduced amounts of IL-12p70, IL-6, and IL-23, as compared with control DCs. The results indicated that RelB-silenced DCs were able to create a particular cytokine environment that was absent of inflammatory cytokines.

Assaying cytokines of RelB-silenced DCs. Supernatants of control DCs and RelB-silenced DCs were collected for IL-12p70, IL-6, and IL-23 measurement by ELISA. Data are presented as the mean ± SD values of triplicate cultures. Results shown are representative of three independent experiments
RelB-silenced DCs spurred on the CD4+ T cells from Th1/Th17 to the Th2 cell subset in vitro
To define Th differentiation of CD4+ T cells primed by RelB-silenced DCs, TAChR-specific CD4+ T cells purified from TAChR-immunized mice and RelB-silenced DCs or control DCs were mixed at a ratio of 4:1. As shown in Fig. 2, supernatants of co-cultures of T cells and RelB-silenced DCs contained low levels of IFN-γ (P < 0.05) and IL-17 (P < 0.01) but high levels of IL-4 (P < 0.05), as compared with supernatants of co-cultures of T cells and control DCs. These results indicated that RelB-silenced DCs possess the ability to shift CD4+ T cells from Th1/Th17 to the Th2 cell subset.

Effect of RelB-silenced DCs on TAChR or Tα146–162 peptide-activated T-cell subsets in vitro. C57BL/6 mice were immunized with TAChR in CFA. After 7 days, CD4+ T cells were purified from the pooled draining lymph nodes and co-cultured with RelB-silenced DCs or control DCs in the presence of TAChR or Tα146–162. IFN-γ, IL-17, and IL-4 production of supernatant harvested after 96 h of co-culture was determined by ELISA. Values are means ± SD. Results shown are representative of three independent experiments
RelB-silenced DC transfer reduced the severity of ongoing clinical EAMG in mice
We investigated the in vivo effects of RelB-silenced DCs on ongoing clinical EAMG. In the control DC-treated group, two mice died due to severe EAMG. As shown in Fig. 3, the average disease severity of RelB-silenced DC-treated animals gradually lessened, and the clinical scores on the day the experiment was terminated averaged 0.67 ± 0.33, while the control DC-treated animals developed significantly more severe disease, and the clinical scores on the day of termination averaged 2.05 ± 0.49 (P < 0.01).

Kinetics of the mean clinical severity of clinical EAMG in RelB-silenced DC-treated mice and control DC-treated mice. Mice were evaluated every other day beginning at the time of initiation of treatment as described in “Materials and methods.” Disease severity was significantly lower in RelB-silenced DC-treated mice compared to the control DC-treated mice (*P < 0.05, **P < 0.01)
RelB-silenced DC transfer decreased anti-mouse-AChR IgG levels
The effects of RelB-silenced DC treatment on the serum anti-AChR Ab response were detected on day 20 after initiation of the treatment. We measured the serum concentration of anti-mouse AChR Abs by ELISA with affinity-purified mouse AChR as antigen. As shown in Fig. 4, treatment with RelB-silenced DCs resulted in a decrease in total anti-mouse AChR IgG Ab response compared with the control DC group (P < 0.01). This decrease in total anti-mouse AChR IgG levels reflected a prominent decrease in complement-fixing IgG2b isotypes and non-complement-fixing IgG1 isotypes [15], while another non-complement-fixing IgG2c isotype was relatively unaffected.

Serum anti-mouse AChR IgG isotypes measured by ELISA. At day 20 following the first treatment, serum levels of anti-AChR IgG, IgG1, and IgG2b but not IgG2c Abs were reduced significantly in RelB-silenced DC-treated mice as compared with control DC-treated mice. Values are the mean ± SD of three individual experiments conducted in triplicate
RelB-silenced DC transfer led to reduced lymphocyte proliferation to TAChR and its dominant peptide Tα146–162
To test the biological modulatory effect of RelB-silenced DCs in the lymphocyte proliferation responses to AChR and its dominant peptide, splenocytes from TAChR-immunized mice were stimulated in vitro with mouse TAChR or Tα146–162 peptide or ConA at the termination of the experiment. Splenocytes from mice treated with RelB-silenced DCs had a significantly lower (P < 0.01) proliferative response to T-AChR and Tα146–162 peptide compared with mice treated with control DCs. No differences were observed in the RelB-silenced DC-treated, control DC-treated mice with nonspecific stimulation using the ConA (Fig. 5a).

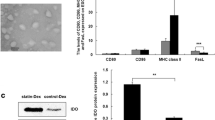
Reduced TAChR- and Tα146–162 peptide-specific lymphocyte proliferative response and cytokine profile shift in RelB-silenced DC-treated mice. Control DC-treated and RelB-silenced DC-treated mice were sacrificed on day 21 to obtain splenocytes. a The proliferative response to TAChR, Tα146–162, or ConA was determined with 3H incorporation. Splenocytes from mice treated with RelB-silenced DCs had a significantly lower proliferative response to T-AChR and Tα146–162 peptide compared with mice treated with control DC (P < 0.01), but no differences were observed in the two groups of mice with nonspecific stimulation using the lectin ConA (P > 0.05). b Cytokine response to TAChR stimulation. Culture supernatants collected at 72 h were examined by ELISA for IFN-γ, IL-4, and IL-17 content. Significantly lower levels of IFN-γ, IL-17, and higher levels of IL-4 were detected in RelB-silenced DC-treated animals compared with the control DC group (P < 0.01). c Western blot analysis of protein expression of T-bet, GATA-3, RORγt, and FoxP3. It was shown that RelB-silenced DC treatment enhanced the expression of GATA-3 and FoxP3, while T-bet and RORγt decreased. β-actin served as an internal control. Results shown are representative of three independent experiments
RelB-silenced DC transfer inhibited Th1/Th17 and promoted the Th2 cell profile in mice with EAMG
To assess the effects of RelB-silenced DC transfer on the production of Th1, Th2, and Th17 cytokines, the levels of IFN-γ, IL-4, and IL-17 in splenocytes were measured by ELISA (Fig. 5b). Our results showed that transfer of RelB-silenced DC led to lower amounts of IFN-γ and IL-17 but higher amounts of IL-4 in the mice compared with the control DC group. The observed reduction of the Th1 and Th17 mediators and the increase of the Th2 mediator after treatment with RelB-silenced DCs was further confirmed by immune-blot analysis of the T-bet, RORγt, and GATA3 protein (Th1, Th17, Th2 lineage commitment factors, respectively). As shown in Fig. 5c, administration of RelB-silenced DCs led to a significant reduction in T-bet and RORγt protein expression compared with the control DC group (P < 0.01). In contrast, RelB-silenced DC transfer led to a significant induction of GATA3 protein expression compared with the control DC group (P < 0.01). These data showed that only RelB-silenced DCs were able to induce a shift towards Th2 cytokine production after cell transfer into mice.
Therapeutic effects of RelB-silenced DCs are associated with Treg formation
To further investigate the mechanism of the therapeutic function of RelB-silenced DC, we next analyzed whether this effect was correlated with enhanced accumulation or preservation of regulatory T cells (Treg). For analysis of Treg of spleen lymphocytes, we used Western blotting to determine the expression of FoxP3, a transcription factor that is specifically expressed by this subset of T cells [16]. As shown in Fig. 5c, we observed significantly increased levels of FoxP3 protein in splenocytes obtained from RelB-silenced DC-treated mice, suggesting that FoxP3+ regulatory T cells could have an important regulatory role in the RelB-silenced DC-mediated suppression of the disease.
Discussion
Dendritic cells (DCs) are professional APCs that play a crucial role as initiators and modulators of adaptive immune responses [17]. The immunostimulatory or immunoregulatory properties of DCs depend on maturation status, phenotype, and source of origin. Owing to the lack of one or more activation signals, immature DCs (iDCs) can inhibit T-cell responses via the mechanisms of apoptosis [18], energy [19], and default Th2 differentiation [20, 21]. In addition, iDCs have the potential to trigger generation of regulatory T cells (Tregs) [22]. RelB is the transcription factor that has been associated most directly with DC differentiation and function [23].
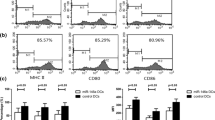
RNA interference (RNAi) is an evolutionarily conserved process of sequence-specific post-transcriptional gene silencing that is triggered by double-stranded RNA (dsRNA) [24]. The mediators of sequence-specific degradation of target mRNA are 21- to 23-nucleotide double-stranded small interfering RNAs (siRNAs) generated by cleavage from longer dsRNA by Dicer, an RNase-III-like enzyme [25, 26]. Exogenous administration of siRNA is capable of blocking gene expression in mammalian cells without triggering the nonspecific panic response [24]. Lentiviral vectors are very efficient at transducing dividing and quiescent cells, which makes them highly useful tools for genetic analysis and gene therapy [27]. We previously demonstrated that silencing RelB using lentiviral-mediated RNAi results in arrest of DC maturation with reduced expression of MHC class II and costimulatory markers [12]. In this study, we also demonstrated that RelB-silenced DCs inhibit LPS-stimulated upregulation of IL-12, IL-23, and IL-6, which are important cytokines that can be secreted by mDC [28, 29].
MG, as well as its animal model, EAMG, is a typical autoantibody-mediated disease. T helper cells specific to AChR and the cytokines they secrete may play a pathogenic role by permitting and facilitating the synthesis of high-affinity anti-AChR antibodies [30]. Traditionally, Th cells were divided into the Th1 and Th2 functional subsets based on whether they express IFN-γ (Th1) or IL-4 (Th2) [31]. IFN-γ appears to promote EAMG. It has been shown to be necessary to the induction and development of EAMG [32], and IFN-γ receptor knockout mice are resistant to EAMG [33, 34].
Th2 effector cytokine IL-4 may have a protective role in EAMG. First, IL-4-deficient mice are more susceptible to EAMG induction [35]. Second, IL-4 appears to be involved in the differentiation of AChR-specific regulatory CD4+ T cells, which can prevent the development of EAMG and its progression to a self-maintaining, chronic autoimmune disease [30, 36]. Third, tolerance procedures in mice have been found to be effective in preventing EAMG through the induction of Th1 unresponsiveness, which was associated with an upregulation of Th2 cytokine synthesis [31, 37]. Consistent with previous reports that iDC or DC absence of IL-12R plays a prominent regulatory role in the induction of antigen-specific Th2 cells [28, 38], our data demonstrated that RelB-silenced DC transfer leads to prominent upregulation of IL-4 and downregulation of IFN-γ in EAMG. To further support these observations, the application of RelB-silenced DC transfer in our study evidently led to a downregulation of T-bet expression, whereas GATA3 protein expression was significantly upregulated. Our results are also in agreement with recent reports that T-bet−/− mice are less susceptible to EAMG [39].
Recent evidence suggests that naive T cells can also be induced to differentiate along a pathway favoring development of Th17 or Treg cells in a mutually exclusive manner [40–42]. The Th17 population, which is characterized by production of IL-17, is important in mediating autoimmune diseases such as rheumatoid arthritis and EAE in animals [43, 44]. A very recent study reported that TAChR-immunized IL-17−/− mice exhibited significantly milder EAMG and reduced anti-AChR IgG2b compared to control B6 mice [10], indicating that Th17 plays a central role in humoral immunity in EAMG. In fact, using mice genetically deficient in IL-12/IL-23 and IFN-λ (dKO mice) to examine the effects of IL-12 and IFN-λ on EAMG susceptibility, Wang et al. provided evidence that Th1 cytokines are not the exclusive players in EAMG-pathogenic Th17 cells and reduced CD4+CD25+ Treg function may be involved in the mechanisms of this Ab-mediated experimental autoimmune disease [9]. Their data are in accordance with the clinical research that CD4+CD25+ Treg cells, which play a key role in maintaining peripheral tolerance to self-Ag, may be functionally impaired in MG patients [45, 46].
On the whole, the role of Th profiles in EAMG indicated above fits with a novel proposed hypothesis with regards to inflammatory and autoimmune diseases [47], namely that skewing of responses towards Th17 or Th1 and away from Treg and Th2 may be responsible for the development and progression of autoimmune diseases in humans. In the present study, in addition to exploring the effect of RelB-silenced DCs on Th1/Th2, we also detected the influence of RelB-silenced DCs on Th17 and Treg. Consistent with reports that inhibiting NF-κB or downregulating RelB endows DCs with the ability to induce Treg formation [48, 49], our data indicated that RelB-silenced DC transfer led to increased FoxP3, a transcription factor that has been implicated as a key controller of development and function of regulatory T cells. In contrast, IL-17 and RORγt decreased significantly. The significant decrease in IL-17 production in splenocytes indicates a low production of biologically active IL-6 and IL-23 [50, 51] by RelB-silenced DCs. Such a decrease could also be due to high IL-4 production, which inhibits differentiation of IL-17-producing CD4+ T cells [7]. The combination of Th2- and Treg-promoting properties and poor proliferation of AChR-reactive T cells suggests that RelB-silenced DCs possess a “tolerogenic” property and would thus be useful in the treatment of EAMG or human MG.
In summary, we propose that RelB-silenced DCs could spur on the AChR-specific T cells from Th1/Th17 to a Th2 subset in vitro and vivo. In addition, administering RelB-silenced BMDCs led to a substantial increase in generation of FoxP3+ Tregs in vivo and effectively suppressed EAMG progression.
References
Vincent A, Palace J, Hilton-Jones D. Myasthenia gravis. Lancet. 2001;357:2122–8.
Berman PW, Patrick J. Linkage between the frequency of muscular weakness and loci that regulate immune responsiveness in murine experimental myasthenia gravis. J Exp Med. 1980;151:204–23.
Hughes BW, Moro De Casillas ML, Kaminski HJ. Pathophysiology of myasthenia gravis. Semin Neurol. 2004;24:21–30.
Elson CJ, Barker RN. Helper T cells in antibody-mediated, organ-specific autoimmunity. Curr Opin Immunol. 2000;12:664–9.
Murphy KM, Reiner SL. The lineage decisions of helper T cells. Nat Rev Immunol. 2002;2(12):933–44.
Mosmann TR, Coffman RL. TH1 and TH2 cells: different patterns of lymphokine secretion lead to different functional properties. Annu Rev Immunol. 1989;7:145–73.
Park H, Li Z, Yang XO, Chang SH, Nurieva R, Wang YH, et al. A distinct lineage of CD4 T cells regulates tissue inflammation by producing interleukin 17. Nat Immunol. 2005;6(11):1133–41.
Ivanov II, McKenzie BS, Zhou L, Tadokoro CE, Lepelley A, Lafaille JJ, et al. The orphan nuclear receptor RORgammat directs the differentiation program of proinflammatory IL-17+ T helper cells. Cell. 2006;126(6):1121–33.
Wang W, Milani M, Ostlie N, Okita D, Agarwal RK, Caspi RR, et al. C57BL/6 mice genetically deficient in IL-12/IL-23 and IFN-gamma are susceptible to experimental autoimmune myasthenia gravis, suggesting a pathogenic role of non-Th1 cells. J Immunol. 2007;178:7072–80.
Bai Y, Liu R, Huang D, La Cava A, Tang YY, Iwakura Y, et al. CCL2 recruitment of IL-6-producing CD11b+ monocytes to the draining lymph nodes during the initiation of Th17-dependent B cell-mediated autoimmunity. Eur J Immunol. 2008;38:1877–88.
Miller SD, McMahon EJ, Schreiner B, Bailey SL. Antigen presentation in the CNS by myeloid dendritic cells drives progression of relapsing experimental autoimmune encephalomyelitis. Ann NY Acad Sci. 2007;1103:179–91.
Zhang Y, Yang H, Xiao B, Wu M, Zhou W, Li J, et al. Dendritic cells transduced with lentiviral-mediated RelB-specific ShRNAs inhibit the development of experimental autoimmune myasthenia gravis. Mol Immunol. 2009;46:657–67.
Yang H, Goluszko E, David C, Okita DK, Conti-Fine B, Chan TS, et al. Mapping myasthenia gravis-associated T cell epitopes on human acetylcholine receptors in HLA transgenic mice. J Clin Invest. 2002;109(8):1111–20.
Wu B, Goluszko E, Christadoss P. Experimental autoimmune myasthenia gravis in the mouse. Curr Protoc Immunol. 2001; chapter 15: unit 15.8.
Yang H, Kala M, Scott BG, Goluszko E, Chapman HA, Christadoss P. Cathepsin S is required for murine autoimmune myasthenia gravis pathogensis. J Immunol. 2005;174:1729–37.
Fontenot JD, Rasmussen JP, Williams LM, Dooley JL, Farr AG, Rudensky AY. Regulatory T cell lineage specification by the forkhead transcription factor foxp3. Immunity. 2005;22(3):329–41.
Vlad G, Cortesini R, Suciu-Foca N. License to heal: bidirectional interaction of antigen-specific regulatory T cells and tolerogenic APC. J Immunol. 2005;174:5907–14.
Lu L, Li W, Zhong C, Qian S, Fung JJ, Thomson AW, et al. Increased apoptosis of immunoreactive host cells and augmented donor leukocyte chimerism, not sustained inhibition of B7 molecule expression, are associated with prolonged cardiac allograft survival in mice preconditioned with immature donor dendritic cells plus anti-CD40L mAb. Transplantation. 1999;68(6):747–57.
Nouri-Shirazi M, Guinet E. Direct and indirect cross-tolerance of alloreactive T cells by dendritic cells retained in the immature stage. Transplantation. 2002;74:1035–44.
Buonocore S, Flamand V, Goldman M, Braun MY. Bone marrow derived immature dendritic cells prime in vivo alloreactive T cells for interleukin-4-dependent rejection of major histocompatibility complex class II antigen-disparate cardiac allograft. Transplantation. 2003;75(3):407–13.
Christensen HR, Frøkiaer H, Pestka JJ. Lactobacilli differentially modulate expression of cytokines and maturation surface markers in murine dendritic cells. J Immunol. 2002;168(1):171–8.
Watanabe N, Wang YH, Lee HK, Ito T, Wang YH, Cao W, et al. Hassall’s corpuscles instruct dendritic cells to induce CD4+CD25+ regulatory T cells in human thymus. Nature. 2005;436(7054):1181–5.
Martin E, O’Sullivan B, Low P, Thomas R. Antigen-specific suppression of a primed immune response by dendritic cells mediated by regulatory T cells secreting interleukin-10. Immunity. 2003;18(1):155–67.
Elbashir SM, Harborth J, Lendeckel W, Yalcin A, Weber K, Tuschl T. Duplexes of 21-nucleotide RNAs mediate RNA interference in cultured mammalian cells. Nature. 2001;411(6836):494–8.
Fire A, Xu S, Montgomery MK, Kostas SA, Driver SE, Mello CC. Potent and specific genetic interference by double-stranded RNA in Caenorhabditis elegans. Nature. 1998;391(6669):806–11.
Hannon GJ. RNA interference. Nature. 2002;418(6894):244–51.
Ageichik A, Collins MK, Dewannieux M. Lentivector targeting to dendritic cells. Mol Ther. 2008;16(6):1008–9.
Jiang HR, Muckersie E, Robertson M, Forrester JV. Antigen-specific inhibition of experimental autoimmune uveoretinitis by bone marrow-derived immature dendritic cells. Invest Ophthalmol Vis Sci. 2003;44:1598–607.
Waibler Z, Kalinke U, Will J, Juan MH, Pfeilschifter JM, Radeke HH. TLR-ligand stimulated interleukin-23 subunit expression and assembly is regulated differentially in murine plasmacytoid and myeloid dendritic cells. Mol Immunol. 2007;44:1483–9.
Milani M, Ostlie N, Wu H, Wang W, Conti-Fine BM. CD4+ T and B cells cooperate in the immunoregulation of experimental autoimmune myasthenia gravis. J Neuroimmunol. 2006;179(1–2):152–62.
Saoudi A, Bernard I, Hoedemaekers A, Cautain B, Martinez K, Druet P, et al. Experimental autoimmune myasthenia gravis may occur in the context of a polarized Th1- or Th2-type immune response in rats. J Immunol. 1999;162(12):7189–97.
Balasa B, Deng C, Lee J, Bradley LM, Dalton DK, Christadoss P, et al. Interferon gamma (IFN-gamma) is necessary for the genesis of acetylcholine receptor-induced clinical experimental autoimmune myasthenia gravis in mice. J Exp Med. 1997;186(3):385–91.
Zhang GX, Xiao BG, Bai XF, van der Meide PH, Orn A, Link H. Mice with IFN-gamma receptor deficiency are less susceptible to experimental autoimmune myasthenia gravis. J Immunol. 1999;162(7):3775–81.
Balasa B, Deng C, Lee J, Christadoss P, Sarvetnick N. The Th2 cytokine IL-4 is not required for the progression of antibody-dependent autoimmune myasthenia gravis. J Immunol. 1998;161(6):2856–62.
Karachunski PI, Ostlie NS, Okita DK, Conti-Fine BM. Interleukin-4 deficiency facilitates development of experimental myasthenia gravis and precludes its prevention by nasal administration of CD4+ epitope sequences of the acetylcholine receptor. J Neuroimmunol. 1999;95(1–2):73–84.
Milani M, Ostlie N, Wang W, Conti-Fine BM. T cells and cytokines in the pathogenesis of acquired myasthenia gravis. Ann NY Acad Sci. 2003;998:284–307.
Sheng JR, Li L, Ganesh BB, Vasu C, Prabhakar BS, Meriggioli MN. Suppression of experimental autoimmune myasthenia gravis by granulocyte–macrophage colony-stimulating factor is associated with an expansion of FoxP3+ regulatory T cells. J Immunol. 2006;177(8):5296–306.
Zhang GX, Yu S, Gran B, Li J, Siglienti I, Chen X, et al. Role of IL-12 receptor beta 1 in regulation of T cell response by APC in experimental autoimmune encephalomyelitis. J Immunol. 2003;171:4485–92.
Liu R, Campagnolo D, Vollmer T, Shi FD. Transcriptional factor T-bet determines the susceptibility to experimental myasthenia gravis. Clin Immunol. 2008;127(Suppl 1):S48.
Bettelli E, Carrier Y, Gao W, Korn T, Strom TB, Oukka M, et al. Reciprocal developmental pathways for the generation of pathogenic effector TH17 and regulatory T cells. Nature. 2006;441(7090):235–8.
Veldhoen M, Hocking RJ, Atkins CJ, Locksley RM, Stockinger B. TGFbeta in the context of an inflammatory cytokine milieu supports de novo differentiation of IL-17-producing T cells. Immunity. 2006;24(2):179–89.
Mangan PR, Harrington LE, O’Quinn DB, Helms WS, Bullard DC, Elson CO, et al. Transforming growth factor-beta induces development of the T(H)17 lineage. Nature. 2006;441(7090):231–4.
Serada S, Fujimoto M, Mihara M, Koike N, Ohsugi Y, Nomura S, et al. IL-6 blockade inhibits the induction of myelin antigen-specific Th17 cells and Th1 cells in experimental autoimmune encephalomyelitis. Proc Natl Acad Sci USA. 2008;105:9041–6.
Notley CA, Inglis JJ, Alzabin S, McCann FE, McNamee KE, Williams RO. Blockade of tumor necrosis factor in collagen-induced arthritis reveals a novel immunoregulatory pathway for Th1 and Th17 cells. J Exp Med. 2008;205(11):2491–7.
Balandina A, Lécart S, Dartevelle P, Saoudi A, Berrih-Aknin S. Functional defect of regulatory CD4(+)CD25+ T cells in the thymus of patients with autoimmune myasthenia gravis. Blood. 2005;105:735–41.
Sun Y, Qiao J, Lu CZ, Zhao CB, Zhu XM, Xiao BG. Increase of circulating CD4+CD25+ T cells in myasthenia gravis patients with stability and thymectomy. Clin Immunol. 2004;112:284–9.
Weaver CT, Harrington LE, Mangan PR, Gavrieli M, Murphy KM. Th17: an effector CD4 T cell lineage with regulatory T cell ties. Immunity. 2006;24(6):677–88.
Yang J, Bernier SM, Ichim TE, Li M, Xia X, Zhou D, et al. LF15–0195 generates tolerogenic dendritic cells by suppression of NF-κB signaling through inhibition of IKK activity. J Leukoc Biol. 2003;74(3):438–47.
Li M, Zhang X, Zheng X, Lian D, Zhang ZX, Ge W, et al. Immune modulation and tolerance induction by RelB-silenced dendritic cells through RNA interference. J Immunol. 2007;178:5480–7.
Langrish CL, Chen Y, Blumenschein WM, Mattson J, Basham B, Sedgwick JD, et al. IL-23 drives a pathogenic T cell population that induces autoimmune inflammation. J Exp Med. 2005;201(2):233–40.
Chen Z, Laurence A, O’Shea JJ. Signal transduction pathways and transcriptional regulation in the control of Th17 differentiation. Semin Immunol. 2007;19(6):400–8.
Acknowledgments
This work was supported by the National Natural Science Foundation of China (30870857) and Hunan Natural Science Foundation (09JJ3079).
Author information
Authors and Affiliations
Corresponding author
Additional information
Responsible Editor: G. Wallace.
H. Yang and Y. Zhang contributed equally to this work.
Rights and permissions
About this article
Cite this article
Yang, H., Zhang, Y., Wu, M. et al. Suppression of ongoing experimental autoimmune myasthenia gravis by transfer of RelB-silenced bone marrow dentritic cells is associated with a change from a T helper Th17/Th1 to a Th2 and FoxP3+ regulatory T-cell profile. Inflamm. Res. 59, 197–205 (2010). https://doi.org/10.1007/s00011-009-0087-6
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00011-009-0087-6