
Abstract
Neuronal cell dysfunction and apoptosis are the main causes of the invasion of the central nervous system by human immunodeficiency virus type 1 (HIV-1), although the underlying mechanism has not been well understood. Recent research has shown that curcumin might play an important role in regulating HIV-1 development. Heat shock protein 70 (HSP70), a protein induced by heat, was reported to inhibit apoptosis through various cell signaling pathways in brain. Overexpression of HSP70 could effectively protected neurons in many animal and cellular models of dementia. In the present study, the expression of HSP70 in the gp120 V3 loop peptide-induced neuronal apoptosis was investigated. Our results demonstrated that gp120 V3 loop peptide could induce primary rat cortical neuronal apoptosis. We also found that curcumin could increase HSP70 expression. In addition, the expression level of both HSP70 mRNA and HSP70 protein were dependent on the curcumin dose in the rat cortical neurons. Curcumin could improve HSP70 expression in gp120 V3 loop peptide-induced primary rat cortical neuronal apoptosis. In general, our results indicated that curcumin played an important role in the gp120 V3 loop peptide induced neuronal apoptosis by regulating HSP70.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Since the first Acquired Immune Deficiency Syndrome (AIDS) case was reported in the 1980s, the human immunodeficiency virus type 1 (HIV-1) pandemic has claimed over 38.6 million lives worldwide and remains one of the major threats to human health with no effective vaccine available. During the disease progression, HIV-1 enters the central nervous system (CNS) at an early stage. Once HIV-1 invades the CNS, it causes significant damages to the nervous system, resulting in neurodegenerative diseases or neurocognitive disorders. Because HIV-1 does not directly infect neurons, cell death and dysfunction in the CNS caused by soluble virus proteins are the characteristics of HIV-1-related neuronal injury. HIV-1 can infect CD4+ T cells and macrophages, leading to the depletion of CD4+ T cells. During HIV-1 infection, the virus binds to the CD4 receptor on the cell surface through the viral glycoprotein subunit gp120. Next, a conformational change occurs in gp120, allowing the virus to bind to a chemokine receptor (CCR5 or CXCR4). Eventually, the HIV membrane fuses with the cell membrane through a conformational change in the trans-membrane glycoprotein, gp41. Additional studies have shown that gp120 could directly interact with neurons, resulting in the neuronal apoptosis [1]. In fact, the severity of brain damage was positively correlated with gp120 expression levels in HIV-1-infected individuals [2]. Those findings provided further evidence that gp120 plays a key role in HIV-1-associated nervous system impairment. At the end of the last century, highly active antiretroviral therapy (HAART) was introduced into clinical practice. HAART was effective in suppressing the systemic HIV-1 viral load and in reducing the mortality rates and incidence of systemic opportunistic infections in AIDS patients. However, chronic infection in the central nervous system (CNS) by HIV-1 still prevails because the blood–brain barrier (BBB) can prevent the permeation of antiviral drugs, lowering the effectiveness of the HAART treatment [3].
HSP70, a 70-kDa heat-shock protein, is a stress-induced molecule that is expressed in response to various types of central nervous system injuries, including stroke, trauma, and neurodegenerative disorders [4]. HSP70 could stimulate HIV-1 or simian immunodeficiency virus (SIV) inhibitors, such as APOBEC3G [5]. Furthermore, human CD4+ T cells could be inhibited by microbial HSP70 [6]. Recent findings have showed that HSP70 has had neuro-protective capability in neurodegenerative diseases such as Parkinson’s disease. This capability was partially due to its role as a chaperone that could attenuate protein aggregation and toxicity and partially due to more direct anti-apoptosis effects.
Curcumin is a hydrophobic polyphenol derived from the root of turmeric (Curcuma longa), a rhizomatous herbaceous perennial plant of the ginger family. Many studies have shown that curcumin has antioxidant [7], anti-inflammatory [8], antimicrobial [9], and anti-carcinogenic capabilitie [10]. It was previously reported that curcumin could improve the learning and memory of rats with HIV-1-gp120-induced memory disorders [11]. Our study was designed to investigate whether curcumin could prevent HIV-1-gp120-mediated nerve cell neuron damages by inhibiting the neuronal apoptosis through the HSP70 pathway.
Materials and Methods
Animals
A total of 15 1- to 2-day-old Sprague–Dawley rats were obtained from the Animal Center of Guangdong province [License No. SCXK (yue) 2006-0015)]. All animal experiment protocols were reviewed and approved by the Ethic Committee of the Third Affiliated Hospital of Guangzhou Medical University.
Cell Cultures
Primary rat cortical neurons were prepared and cultured following the protocol previously reported [11], with slight modifications. Briefly, 1-day-old neonatal Sprague–Dawley rats were euthanized and sterilized with 75 % alcohol. Their scalps and skulls were incised to harvest cerebral hemispheres. The cerebral cortex was washed twice with ice-cold D-Hanks’ solution, and the blood vessels were removed with straight iris forceps. Fresh cortical tissues were sectioned into approximately 1-mm slices using forceps and then werer placed in 15-mL centrifuge tubes containing D-Hanks’ solution and 0.25 % trypsin (volume ratio = 2:1). The sliced tissues were digested for 10 min at 37 °C before the digestion was quenched with 10 % FBS/DMEM/F12. The digested tissues were then mechanically dispersed by lightly blowing with a pipette. The resultant suspension was centrifuged at 1000 rpm for 5 min, and the supernatant was discarded. The residues were subsequently suspended in 10 % FBS/DMEM/F12, and the suspension was filtered through a 200 mesh stainless steel screen cloth into a new 15-mL centrifuge tube. The filtered suspension was transferred to a culture flask so that fibroblasts could be removed through differential adhesions. Next, neurons were seeded with a density of 1 × 106 cells/well onto poly-l-lysine (0.1 mg/mL, Sigma Aldrich-Fluka, St. Louis, MO, USA)-coated cover slips on a 6-well culture plate, followed by incubation in a saturated and humidified incubator containing 5 % CO2 and 95 % air at 37 °C for 4 h before the culture medium was replaced with 2 % B27 neurobasal medium (Life Technologies-Gibco, Grand Island, NY, USA). In the following weeks, half of the medium was replaced with fresh medium twice a week. The cultures were monitored to ensure that neurons constituted more than 95 % of the total cell population. All of the following experiments were performed using primary rat cortical neurons cultured for 7 days.
HIV-1 gp120 V3 Loop Peptide
According to a previously reported [12]. The HIV-1 IIIB gp120 V3 loop was synthesized by Shanghai Apeptide (Shanghai, China; sequence: NNTRKSIRIERGPGRAFVTIGKIG; molecular formula: C114H199N41O31; molecular weight: 2.64 kDa).
Analysis of Cortical Neuron Apoptosis
The FITC Annexin V/Dead Cell Apoptosis Kit (Life technologies, USA) was used to measure cell apoptosis. Annexin V was labeled with a fluorophore to identify apoptotic cells by binding to phosphatidylserine (PS) exposed on the outer leaflet. Propidium iodide (PI), which labeled with red-fluorescent, could bind tightly to nucleic acid in dead cells. Beriefly, primary rat cortical neurons were seeded with a density of 2 × 105 cells/well in a 6-well culture plate for 5 days. The seeded cells on the plate were then incubated with HIV-1 gp120 V3 loop (1 mg/mL) for another 48 h, followed by the apoptosis analysis using the flow cytometry method. The above cultured cells were washed twice with cold PBS before being resuspended in 1× binding buffer with a concentration of 1 × 106 cells/mL. Next, 100 μL of the resulted suspension (1 × 105 cells) was transferred to a 5-mL culture tube containing 5 μL of FITC Annexin V and 5 μL PI. The resultant mixture was gently vortexed and incubated for 15 min at RT (25 °C) in the dark before 400 μL of 1× binding buffer was added into each tube. The resultant cells were analyzed by flow cytometry within 1 h. The green fluorescence of Annexin V-FITC was measured using FL1, and the red fluorescence of PI was measured using the FL3 channel. Fluorescent compensation was adjusted for normal neurons to remove the spectral overlap and confirm the exact gate site.
Construction of the Adenoviral Vector System
Briefly, according to our previous study [13], the HSP70 gene was linked to the pShuttle Vector. The confirmed pshuttle vector was linearized with the PmeI (NEB) restriction endonuclease. Next, 20 µL of electro competent AdEasier cells were mixed with an additional 8.0 µL of ethanol-precipitated. The linearized shuttle vector was placed in an ice-cold 2-mm cuvette. Pulses were delivered at 2500 V, 200 O and 25 mF in a Gene pulser electroporator (Bio-Rad, USA). The transformation mix was re-suspended in 100 µL of LB medium plated placed in an LB/kanamycin plate and was placed in a 37 °C incubator to grow overnight (16–20 h). The 20 smallest colonies were picked, and each was grown in 2.5 mL of LB medium containing 25 mg m/L kanamycin for 15 h in a 37 °C orbital shaker. Next, we performed mini-preps, using the conventional alkaline lysis method to check the size of super-coiled plasmids by running one-fifth of each mini-prepson on a 0.8 % agarose gel. PacI restriction digestion was performed on candidate selected clones. Correct recombinants yielded a larger fragment with molecular weight approximately 30 kb and a smaller fragment of approxiamtedl 3.0 or 4.5 kb (data not shown).
Western Blot Analysis
Apoptosis Western Blot Cocktail was purchased from Abcam (Cambridge, MA). Cells or tissues were washed twice with ice-cold PBS and were harvested in with sample buffer. The resultant suspensions were centrifuged at 12,000×g for 30 min at 4 °C to yield the soluble extracts with the protein concentration determined using the BCA kit (Pierce Chemical Co., Rockford, IL). The same amount of proteins (40 μg) for each sample was resolved in SDS-PAGE. After electrophoresis, proteins were transferred to nitrocellulose membranes and then incubated in TBST containing 5 % nonfat milk at room temperature for 1 h. The resultant blots were then incubated with primary antibodies overnight at 4 °C. The antibodies used in the Western blot analysis included HSP70 and β-actin antibodies (Abcam, USA). The blots were next washed with TBST, and the membranes were then incubated with the corresponding horseradish peroxidase-conjugated secondary antibodies (Cell signaling, USA). The signals were visualized using ECL (Pierce Chemical Co. USA) and then recorded on X-ray films.
RNA Extraction and Quantification of Real Time PCR
The cells were incubated for 8 min in 25 mL lysis solution containing DNase I at room temperature. The incubation was subsequently quenched with 2.5 μL of stop solution, followed by RT reaction. Each RT reaction mixture contained 1.5 μL of 10× RT Buffer, 0.15 μL of dNTP Mix, 0.19 μL of RNase Inhibitor, 1 μL of MultiScribe (TM) RT, 4.16 μL of nuclease-free water, 3 μL of RT primer, and 5 μL of Cells-to-CT sample lysate. Unless otherwise indicated, all reagents used were purchased from ABI (Applied Biosystems). The RT reaction was carried out on a thermal cycler (gene AMP PCR system 9700) at the following conditions: 30 min at 16 °C, 30 min at 42 °C, and 5 min at 85 °C. The products were stored at 4 °C.
Reactions for qRT-PCR including that those for negative control were performed in duplicate. The hybridization probe assay specifically for real-time PCR detection of mRNAs was optimized using the 2× TaqMan Mastermix and TaqManm RNA assay following the manufacturer’s instructions. To evaluate the repeatability of the RT-PCR at the level of the single cell, the cDNA reaction after RT for individual cell was repeated for three times. The PCR reaction mixture contained 10 μL 2× TaqMan Mastermix, 1 μL of TaqManm RNA assay, 7.67 μL of Nuclease-free water, and 1.33 μL of RT products. Reactions were carried out in a thermal cycler (ABI 7500 real-time PCR) under the following conditions: 95 °C for 10 min, followed by 40 cycles of 95 °C for 10 s and 58 °C for 60 s. We used a relative quantification method with cell-number normalization in this study. ΔCt was calculated as ΔCt = Ct (calibrator) – Ct (test), with Ct value of the first sample set as the default. The change was calculated as ratio (test/calibrator) = 2 Ct (calibrator) – Ct (test). Unless otherwise indicated, all relative quantifications were carried out using this method. Significance was defined according to p values from two-tailed t test analysis, for single-cell ΔCt comparisons, one-tailed t test analysis was performed.
Statistical Analysis
The data are expressed as the mean ± SD. Statistical significance was assessed by Student’s t tests or ANOVA, as appropriate. A p value less than 0.05 indicated statistical significance.
Results
HIV-1 gp120 V3 Loop Peptide Induces Primary Rat Cortical Neuronal Apoptosis
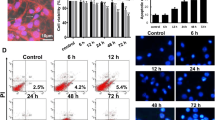
Apoptosis is an important feature of HIV-1-associated central neurological dysfunction. Apoptotic neurons have been observed in the CNS of HIV-1-infected patients [14]. The ratio of apoptotic cells was examined by flow cytometry in this study. After the neuron cells were treated with 0.5 or 1.0 μg/mL peptide, the ratio of apoptotic cells was 14.3 ± 1.5 or 64.72 ± 5.9 % respectively (Fig. 1). The above results indicated that HIV-1 gp120 V3 loop peptide could induce the apoptosis of primary rat cortical neurons by activating the caspase-dependent pathway.

HIV-1 gp120 V3 loop peptide induces the apoptosis of primary rat cortical neurons. Primary rat cortical neurons that had been cultured for 7 days were treated with corresponding medium for 48 h and then were subjected to apoptosis analysis. a–c Apoptosis of neurons was determined by flow cytometry. Experiments were repeated three times (the experimental groups vs. the control group, *p < 0.05)
Curcumin Reverses gp120 V3 Peptide-Induced Cell Apoptosis
We analyzed the effect of gp120 V3 peptide induced cell apoptosis in the presence of curcumin. As shown in the Fig. 2, the ratio of apoptotic cells induced by the peptide was examined by flow cytometry. When the cells were treated with 0.5 μg/mL peptide, the ratios of the later and early apoptotic cells were 12.8 ± 3.4 and 4.4 ± 0.4 %, resepectively (Fig. 2b). When the cells were treated with 1 μg/mL curcumin, the ratio of later and early apoptotic cells were 3.1 ± 0.2 and 2.0 ± 0.1 %, respectively (Fig. 2c, d). In addition, the expression of cleaved caspase-3, an apoptosis-specific protein, increased in a dose-dependent manner. Compared with the gp120-peptide-treated group, curcumin (1 μg/mL) decreased the expression of cleaved caspase-3 in the presence of gp120 peptide, while the expression of procaspase-3 was increased (Fig. 2e, f). These results indicated that curcumin reverses the later and early cell apoptosis induced by gp120 V3 peptide.

Curcumin reverses gp120-peptide-induced cell apoptosis. HIV-1 gp120 V3 loop peptide could induce the apoptosis of primary rat cortical neurons. Primary rat cortical neurons that had been cultured for 7 days were treated with corresponding medium for 48 h and then were subjected to apoptosis analysis. a–d Apoptosis of neurons was determined by flow cytometry. e, f Apoptosis of neurons was determined by Western blot analysis of cleaved caspase-3 and procaspase-3 (experimental groups vs. medium control group, *p < 0.05; **p < 0.05). Experiments were repeated three times
HIV-1 gp120 V3 Peptide Decreases the Expressions of HSP70 in Primary Rat Cortical Neurons
We use Western blot and real-time PCR assay to measure HSP70 protein and gene expression in the primary rat cortical neurons treated with HIV-1 gp120 V3 loop peptide. Our research showed that this peptide could reduce the expressions level of HSP70 and HSP70 protein in primary rat cortical neurons in a dose-dependent manner (Fig. 3). Compared with the control group (100 % of rat cortical neurons), the experimental groups treated with 0.5 and 1 mg/mL HIV-1 gp120 V3 loop peptide had significantly lower HSP70 and HSP70 (65.6 ± 3.5 and 15.1 ± 1.3 %, respectively).

Expression of HSP70 in rat cortical neurons. After rat cortical neurons were treated with peptide for 48 h, the expressions levels of HSP70 and HSP70 were analyzed by real-time PCR and Western blot, respectively. a The expression of HSP70 was analyzed by Western blot. b The expression of HSP70 was analyzed by real-time PCR. Each sample was tested three times, and the data are presented as the mean ± SD. The experiments were repeated twice, and repeatable results were obtained (mean ± SD, n = 3, *p < 0.01)
Curcumin Could Increase the Expression of HSP70 in Primary Rat Cortical Neurons
We used Western blot and real-time PCR assay to measure the effects of HSP70 protein and gene expression on the primary rat cortical neurons treated with curcumin. Our research demonstrated that curcumin increases the expressions level of HSP70 and HSP70 protein in primary rat cortical neurons in a dose-dependent manner (Fig. 4). Compared with medium control (100 % of rat cortical neurons), the experimental groups treated with 0.5 and 1 μg/mL curcumin had significantly higher expression levels of HSP70 and HSP70 (1.5 ± 2.5 and 2.1 ± 1.2 % fold, respectively).

Curcumin increased the expressions of HSP70 and HSP70 protein in primary rat cortical neurons in a dose-dependent manner. a The expression of HSP70 was analyzed by Western blot. b The expression of HSP70 was analyzed by real-time PCR. The experiments were repeated twice, and repeatable results were obtained (mean ± SD, n = 3, *p < 0.01)
Curcumin Could Reverse the Negative Important of HIV-1 gp120 V3 Peptide on the Expression of HSP70 in Primary Rat Cortical Neurons
We used Western blot and real-time PCR assay to measure the impact of curcumin on HSP70 protein and gene expression in neurons treated with HIV-1 gp120 V3 loop peptide. The primary rat cortical neurons were treated with HIV-1 gp120 V3 loop peptide for the first 4 h, before 0.5 or 1 μg/mL curcumin was added. It turned out that curcumin could reverse the impact of the peptide on the expression of HSP70 and HSP70 protein in primary rat cortical neurons in a dose-dependent manner (Fig. 5). Compared with medium control (100 % of rat cortical neurons), the experimental groups treated with 0.5 or 1 μg/mL curcumin had significantly higher expressions levels of HSP70 and HSP70 protein (1.1 ± 0.1 and 1.2 ± 0.2 % fold, respectively).

Curcumin reversed the expressions levels of HSP70 and HSP70 protein in primary rat cortical neurons treated with HIV-1 gp120 V3 loop peptide. a The expression of HSP70 was analyzed by Western blot. b The expression of HSP70 was analyzed by real-time PCR. Each sample was tested in triplicate, and the data were presented as the mean ± SD (the experiments were repeated twice, and repeatable results were obtained, *p < 0.01; **p < 0.05)
Curcumin Decreases the HIV-1 gp120 V3 Loop Peptide Induced Apoptosis Through the HSP70 Pathway
We used flow cytometry to measure the impact of curcumin on the HSP70 pathway in neurons treated with HIV-1 gp120 V3 loop peptide. The primary rat cortical neurons were treated with HIV-1 gp120 V3 loop peptide first for 4 h before we silenced HSP70 by HSP70 siRNA or over-expressed HSP70 by Adenovirus vector system before 1 μg/mL curcumin was added. Curcumin was shown to decrease apoptosis through the HSP70 pathway compared with that in the gp120-peptide-treated group, the ratio of apoptosis cell was decreased in the group treated with HSP70siRNA and curcumin. Furthermore, the ratio of apoptosis cell was increased in the curcumin-treated group with HSP70 overexpression compare with that in the HSP70 siRNA treatment group (Fig. 6). We also used Western blot and real-time PCR assay to measure apoptosis protein expression to determine the protection of curcumin on the HSP70 pathway in neurons treated under the condition of HIV-1 gp120 V3 loop peptide. As shown in Fig. 7, corresponding to the expression of procaspase-3 decreasing, the expression of cleaved caspase-3, an apoptosis-specific protein, increased in the group treated with HSP70siRNA and curcumin Furthermore, corresponding to the expression of procaspase-3 increasing, the expression of cleaved caspase-3 decreased in the curcumin-treated group with HSP70 overexpression. These results indicated that curcumin decreased the gp120 V3 loop peptide-induced apoptosis through the HSP70 pathway.

Curcumin reverses the gp120-peptide-induced apoptosis by targeting HSP70. Primary rat cortical neurons that had been cultured for 7 days were treated with corresponding medium for 48 h and then were subjected to apoptosis analysis by flow cytometry. a Control: b gp120 peptide. c gp120 peptide + HSP70 siRNA + curcumin; d gp120 peptide + HSP70 + curcumin. Each sample was tested in triplicate, and the data were presented in mean ± SD

Curcumin reverses gp120-peptide-induced apoptosis through the caspase-3 pathway. Primary rat cortical neurons that had been cultured for 7 days were treated with corresponding medium for 48 h and then were subjected to apoptosis analysis. a The expression of procaspase-3 and cleaved caspase 3 were analyzed by Western blot. b The expression of procaspase-3 and cleaved caspase 3 were analyzed by real-time PCR. Each sample was tested in triplicate, and the data were presented in mean ± SD (the experiments were repeated twice, and repeatable results were obtained, *p < 0.01; **p < 0.05)
Discussion
Although there is no evidence that HIV-1 could directly infect neurons, the central nervous system becomes damaged once HIV-1 invades into the central nervous system. HIV-1 soluble virus proteins, such as gp120 protein, could induce cell apoptosis and dysfunction in the CNS [15].
Our research also found that HIV-1 gp120 V3 loop peptide directly induced apoptosis of primary rat cortical neurons, in agreement with these previous results. We demonstrated that HIV-1 gp120 V3 loop peptide could lower the expressions of HSP70 in a dose-dependent manner. However, curcumin could reverse the impact of HIV-1 gp120 V3 loop peptide on HSP70 expressions. Curcumin reversed the gp120-peptide-induced apoptosis through the HSP70 pathway. During the course of HIV infection, the binding of gp120 to the CXCR4 or CCR5 receptors triggers the intracellular responses, including activations of the transcription factors, the MAPK pathway, and ion channels [16–18]. It is speculated that the primary mechanism of HIV entry into the central nervous system (CNS) is via a “Trojan Horse” mechanism and that HIV neurotoxicity is primarily an indirect effect of HIV infection. It was reported that the viral products, such as gp120, Tat, Nef, Vpr and Rev, participated in the progression of HIV-1 infection of CNS [19, 20]. HIV-1 gp120, an envelope protein that mediates the entry of the virus into the host cell [21, 22], has been shown to induce neuro-pathological changes in mice similar to that seen in HAND patients [23, 24]. In addition, HIV-1 gp120 can induce the neurotoxicity through various pathways such as caspase-3 pathway [25, 26].
HSP70, a 70-kDa heat-shock protein, is a stress-induced molecule in response to various types of central nervous system injuries, including stroke, trauma, and neurodegenerative disorders. HPS70 has high neuro-protective capability [27] and plays an important role in the folding, assembling, and stabilization of newly synthesized proteins, the refolding of mis-folded proteins, the degradation of abnormal proteins, and the activation of regulatory proteins [28]. Recent findings have suggested that HSP70 showed some neuro-protective effects in treating neurodegenerative diseases such as Parkinson’s disease. The high neuro-protective capability of HPS70 may be due to its role as a chaperone that attenuates protein aggregation and toxicity and its direct anti-apoptotic capability [29]. Others have also reported that HPS70 showed strong protective effects on the CNS in vivo [30] and in vitro [31]. Recently, Boris and colleagues found that HSP70 modulated both Apaf-1 caspase-dependent and AIF caspase-independent pathways, resulting in the attenuation of apoptosis and ultimately inhibition of neuronal cell death both in primary cortical neurons and SH-SY5Y cells [32]. Our data revealed that gp120 V3 loop peptide could induce primary rat cortical neuronal apoptosis and decrease the expression levels of HSP70 and HSP70. These data suggest that there is a relationship between HSP70 and the apoptosis in CNS. Overexpression of HSP70 could alleviate the gp120-peptide-induced apoptosis.
An ideal anti-neurons apoptosis candidate compound should have three properties: low cost, minimal side effects with high capability to penetrate the blood–brain barrier, and high efficacy. Curcumin may fit all three criteria very well. Curcumin exhibited strong anti-oxidant capability comparable to that of vitamins C and E [33]. It is a potent scavenger of various reactive oxygen species, including superoxide anion radicals, hydroxyl radicals [34, 35] and nitrogen dioxide radicals [36]. Another critical characteristic of curcumin is its ability to cross the blood–brain barrier [37]. Our work also proved that curcumin might have high anti-HIV efficacy. In our experiment, curcumin was incubated with primary rat cortical neurons at 2.5 and 5 μM for 48 h. The experiment showed that curcumin could reverse the inhibition on HSP70 induced by gp120 peptide and increase the expressions of HSP70 and HSP70.
Our results showed that gp120 V3 loop peptide could induce primary rat cortical neuronal apoptosis. In addition, the expressions levels of both HSP70 mRNA and HSP70 protein were curcumin-dose-dependent in the rat cortical neurons treated with curcumin for 48 h. Curcumin could improve HSP70 expression in gp120 V3 loop peptide-induced primary rat cortical neuronal apoptosis. Curcumin played an important role in the gp120 V3 loop peptide-induced neuronal apoptosis by regulating HSP70.
References
Chen L, Liu J, Xu C (2011) HIV-1gp120 induces neuronal apoptosis through enhancement of 4-aminopyridine-senstive outward K+ currents. PLoS ONE 6:e25994
Louboutin JP, Agrawal L, Reyes BA, Van Bockstaele EJ, Strayer DS (2010) HIV-1 gp120-induced injury to the blood-brain barrier: role of metalloproteinases 2 and 9 and relationship to oxidative stress. J Neuropathol Exp Neurol 69:801–816
Xia C, Luo D, Yu X, Jiang S, Liu S (2011) HIV-associated dementia in the era of highly active antiretroviral therapy (HAART). Microbes Infect 13:419–425
Shukla AK, Pragya P, Chaouhan HS, Tiwari AK, Patel DK, Abdin MZ, Chowdhuri DK (2014) Heat shock protein-70 (Hsp-70) suppresses paraquat-induced neurodegeneration by inhibiting JNK and caspase-3 activation in Drosophila model of Parkinson’s disease. PLoS ONE 9:e98886
Pido-Lopez J, Wang Y, Seidl T, Babaahmady K, Vaughan R, Lehner T (2009) The effect of allogeneic in vitro stimulation and in vivo immunization on memory CD4+ T-cell APOBEC3G expression and HIV-1 infectivity. Eur J Immunol 39:1956–1965
Babaahmady K, Oehlmann W, Singh M, Lehner T (2007) Inhibition of human immunodeficiency virus type 1 infection of human CD4+ T cells by microbial HSP70 and the peptide epitope 407-426. J Virol 81:3354–3360
Coradini K, Lima FO, Oliveira CM, Chaves PS, Athayde ML, Carvalho LM, Beck RC (2014) Co-encapsulation of resveratrol and curcumin in lipid-core nanocapsules improves theirin vitro antioxidant effects. Eur J Pharm Biopharm 88:178–185
Antoine F, Girard D (2015) Curcumin increases gelatinase activity in human neutrophils by a p38 mitogen-activated protein kinase (MAPK)-independent mechanism. J Immunotoxicol 12:188–193
Sharma G, Raturi K, Dang S, Gupta S, Gabrani R (2014) Combinatorial antimicrobial effect of curcumin with selected phytochemicals on Staphylococcus epidermidis. J Asian Nat Prod Res 16:535–541
Norris L, Karmokar A, Howells L, Steward WP, Gescher A, Brown K (2013) The role of cancer stem cells in the anti-carcinogenicity of curcumin. Mol Nutr Food Res 57:1630–1637
Tang H, Lu D, Pan R, Qin X, Xiong H, Dong J (2009) Curcumin improves spatial memory impairment induced by human immunodeficiency virus type 1 glycoprotein 120 V3 loop peptide in rats. Life Sci 85:1–10
Guo L, Xing Y, Pan R, Jiang M, Gong Z, Lin L, Wang J, Xiong G, Dong J (2013) Curcumin protects microglia and primary rat cortical neurons against HIV-1 gp120-mediated inflammation and apoptosis. PLoS ONE 8:e70565
Shafer TJ, Hughes MF (2010) Accumulation of pyrethroid compounds in primary cultures from rat cortex. Toxicol In Vitro 24:2053–2057
Xia CL, Zhu P, Cai YT, Zhu GB, Mei ZR, Huang H, Luo DX, Yan PK (2012) HIV-infection resistance in PMBC-derived dendritic cells modified with recombinant virus. Arch Virol 157:413–421
Chi X, Amet T, Byrd D, Chang KH, Shah K, Hu N, Grantham A, Hu S, Duan J, Tao F, Nicol G, Yu Q (2011) Direct effects of HIV-1 Tat on excitability and survival of primary dorsal root ganglion neurons: possible contribution to HIV-1-associated pain. PLoS ONE 6:e24412
Alirezaei M, Kiosses WB, Fox HS (2008) Decreased neuronal autophagy in HIV dementia: a mechanism of indirect neurotoxicity. Autophagy 4:963–966
Liu J, Xu C, Chen L, Xu P, Xiong H (2012) Involvement of Kv1.3 and p38 MAPK signaling in HIV-1 glycoprotein 120-induced microglia neurotoxicity. Cell Death Dis 3:e254
Medders KE, Sejbuk NE, Maung R, Desai MK, Kaul M (2010) Activation of p38 MAPK is required in monocytic and neuronal cells for HIV glycoprotein 120-induced neurotoxicity. J Immunol 185:4883–4895
Herbein G, Gras G, Khan KA, Abbas W (2010) Macrophage signaling in HIV-1 infection. Retrovirology 7:34
Desplats P, Dumaop W, Smith D, Adame A, Everall I, Letendre S, Ellis R, Cherner M, Grant I, Masliah E (2013) Molecular and pathologic insights from latent HIV-1 infection in the human brain. Neurology 80:1415–1423
Espey MG, Ellis RJ, Heaton RK, Basile AS (1999) Relevance of glutamate levels in the CSF of patients with HIV-1-associated dementia complex. Neurology 53:1144–1145
Guttman M, Garcia NK, Cupo A, Matsui T, Julien JP, Sanders RW, Wilson IA, Moore JP, Lee KK (2014) CD4-Induced activation in a soluble HIV-1 Env trimer. Structure 22:974–984
Tamamis P, Floudas CA (2014) Molecular recognition of CCR5 by an HIV-1 gp120 V3 loop. PLoS ONE 9:e95767
Silverstein PS, Shah A, Weemhoff J, Kumar S, Singh DP, Kumar A (2012) HIV-1 gp120 and drugs of abuse: interactions in the central nervous system. Curr HIV Res 10:369–383
Shah A, Singh DP, Buch S, Kumar A (2011) HIV-1 envelope protein gp120 up regulates CCL5 production in astrocytes which can be circumvented by inhibitors of NF-κB pathway. Biochem Biophys Res Commun 414:112–117
Bari M, Rapino C, Mozetic P, Maccarrone M (2010) The endocannabinoid system in gp120-mediated insults and HIV-associated dementia. Exp Neurol 224:74–84
Lu TS, Avraham HK, Seng S, Tachado SD, Koziel H, Makriyannis A, Avraham S (2008) Cannabinoids inhibit HIV-1 Gp120-mediated insults in brain microvascular endothelial cells. J Immunol 181:6406–6416
Aridon P, Geraci F, Turturici G, D’Amelio M, Savettieri G, Sconzo G (2011) Protective role of heat shock proteins in Parkinson’s disease. Neurodegener Dis 8:155–168
Tanaka K, Mizushima T (2009) Protective role of HSP70 against various gastrointestinal diseases. Int J Hyperth 25:668–676
Thakur P, Nehru B (2014) Long-term heat shock proteins (HSPs) induction by carbenoxolone improves hallmark features of Parkinson’s disease in a rotenone-based model. Neuropharmacology 79:190–200
Lim MC, Brooke SM, Sapolsky RM (2003) gp120 neurotoxicity fails to induce heat shock defenses, while the over expression of hsp70 protects against gp120. Brain Res Bull 61:183–188
Moloney TC, Hyland R, O’Toole D, Paucard A, Kirik D, O’Doherty A, Gorman AM, Dowd E (2014) Heat shock protein 70 reduces α-synuclein-induced predegenerative neuronal dystrophy in the α-synuclein viral gene transfer rat model of Parkinson’s disease. CNS Neurosci Ther 20:50–58
Sabirzhanov B, Stoica BA, Hanscom M, Piao CS, Faden AI (2012) Over-expression of HSP70 attenuates caspase-dependent and caspase-independent pathways and inhibits neuronal apoptosis. J Neurochem 123:542–554
Aldebasi YH, Aly SM, Rahmani AH (2013) Therapeutic implications of curcumin in the prevention of diabetic retinopathy via modulation of anti-oxidant activity and genetic pathways. Int J Physiol Pathophysiol Pharmacol 5:194–202
Kawanishi N, Kato K, Takahashi M, Mizokami T, Otsuka Y, Imaizumi A, Shiva D, Yano H, Suzuki K (2013) Curcumin attenuates oxidative stress following downhill running-induced muscle damage. Biochem Biophys Res Commun 441:573–578
Derochette S, Franck T, Mouithys-Mickalad A (2013) Curcumin and resveratrol act by different ways on NADPH oxidase activity and reactive oxygen species produced by equine neutrophils. Chem Biol Interact 206:186–193
Noorafshan A, Ashkani-Esfahani S (2013) A review of therapeutic effects of curcumin. Curr Pharm Des 19:2032–2046
Acknowledgments
This study was supported by Grants from the Project of Science and Technology Department of Guangdong Province (Grant No. 2013B021800188) to Xia C and from the Open Fund of Guangdong Provincial Key Laboratory of Reproductive Medicine (Grant No. 2012125) to Xiao G.
Author information
Authors and Affiliations
Corresponding author
Additional information
Chenglai Xia, Yantao Cai and Shuhua Li have contributed equally to this work.
Rights and permissions
About this article

Cite this article
Xia, C., Cai, Y., Li, S. et al. Curcumin Increases HSP70 Expression in Primary Rat Cortical Neuronal Apoptosis Induced by gp120 V3 Loop Peptide. Neurochem Res 40, 1996–2005 (2015). https://doi.org/10.1007/s11064-015-1695-x
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11064-015-1695-x